
The Role of Oxytocin in Domestic Animal’s Maternal Care: Parturition, Bonding, and Lactation

 , ,
, ,  , , ,
, , ,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
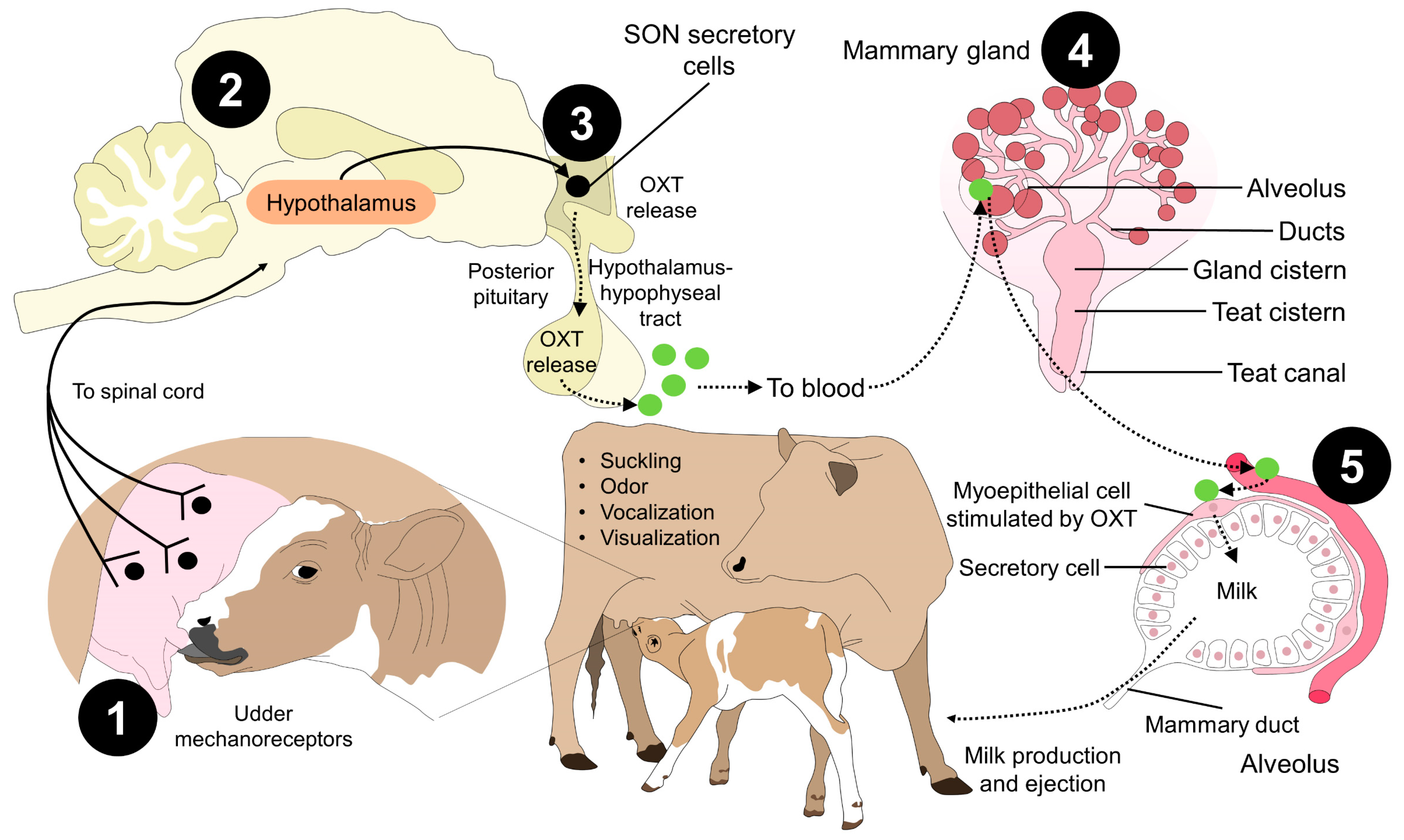{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Neurophysiological Regulation of OXT
3. OXT Participation at Parturition
4. The Role of OXT in the Establishment of Maternal Behavior Regulation
5. Role of OXT in Lactation
6. Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gram, A.; Boos, A.; Kowalewski, M.P. Uterine and placental expression of canine oxytocin receptor during pregnancy and normal and induced parturition. Reprod. Domest. Anim. 2014, 49, 41–49. [Google Scholar] [CrossRef]
- Wang, Q.-H.; Zhang, S.; Qin, L.-M.; Zhang, W.-J.; Liu, F.-H.; Xu, J.-Q.; Ma, Y.-F.; Teng, K.-D. Yimu san improves obstetric ability of pregnant mice by increasing serum oxytocin levels and connexin 43 expression in uterine smooth muscle. J. Zhejiang Univ. B 2017, 18, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Tsingotjidou, A.S. Oxytocin: A multi-functional biomolecule with potential actions in dysfunctional conditions; from animal studies and beyond. Biomolecules 2022, 12, 1603. [Google Scholar] [CrossRef] [PubMed]
- Soloff, M.S.; Chakraborty, J.; Sadhukhan, P.; Senitzer, D.; Wieder, M.; Fernstrom, M.A.; Sweet, P. Purification and characterization of mammary myoepithelial and secretory cells from the lactating rat. Endocrinology 1980, 106, 887–897. [Google Scholar] [CrossRef] [Green Version]
- Faraz, A.; Waheed, A.; Nazir, M.M.; Hameed, A.; Tauqir, N.A.; Mirza, R.H.; Ishaq, H.M.; Bilal, R.M. Impact of oxytocin administration on milk quality, reproductive performance and residual effects in dairy animals—A Review. Punjab Univ. J. Zool. 2020, 35, 61–67. [Google Scholar] [CrossRef]
- Thornton, J.M.; Browne, B.; Ramphul, M. Mechanisms and management of normal labour. Obstet. Gynaecol. Reprod. Med. 2020, 30, 84–90. [Google Scholar] [CrossRef]
- Ogi, A.; Mariti, C.; Pirrone, F.; Baragli, P.; Gazzano, A. The influence of oxytocin on maternal care in lactating dogs. Animals 2021, 11, 1130. [Google Scholar] [CrossRef] [PubMed]
- Lezama-García, K.; Mariti, C.; Mota-Rojas, D.; Martínez-Burnes, J.; Barrios-García, H.; Gazzano, A. Maternal behaviour in domestic dogs. Int. J. Vet. Sci. Med. 2019, 7, 20–30. [Google Scholar] [CrossRef]
- Coria-Avila, G.A.; Pfaus, J.G.; Orihuela, A.; Domínguez-Oliva, A.; José-Pérez, N.; Hernández, L.A.; Mota-Rojas, D. The neurobiology of behavior and its applicability for animal welfare: A review. Animals 2022, 12, 928. [Google Scholar] [CrossRef]
- Jurek, B.; Neumann, I.D. The oxytocin receptor: From intracellular signaling to behavior. Physiol. Rev. 2018, 98, 1805–1908. [Google Scholar] [CrossRef]
- Enriquez, M.F.; Pérez-Torres, L.; Orihuela, A.; Rubio, I.; Corro, M.; Galina, C.S. Relationship between protective maternal behavior and some reproductive variables in Zebu-Type cows (Bos Indicus). Front. Neuroendocrinol. 2021, 9, 2124. [Google Scholar] [CrossRef]
- Ross, H.E.; Young, L.J. Oxytocin and the neural mechanisms regulating social cognition and affiliative behavior. Front. Neuroendocrinol. 2009, 30, 534–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Insel, T.R. The challenge of translation in social neuroscience: A review of oxytocin, vasopressin, and affiliative behavior. Neuron 2010, 65, 768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bales, K.L.; Pfeifer, L.A.; Carter, C.S. Sex differences and developmental effects of manipulations of oxytocin on alloparenting and anxiety in prairie voles. Dev. Psychobiol. 2004, 44, 123–131. [Google Scholar] [CrossRef]
- Madden, J.R.; Clutton-Brock, T.H. Experimental peripheral administration of oxytocin elevates a suite of cooperative behaviours in a wild social mammal. Proc. R. Soc. B Biol. Sci. 2011, 278, 1189–1194. [Google Scholar] [CrossRef] [Green Version]
- Odendaal, J.S.; Meintjes, R. Neurophysiological correlates of affiliative behaviour between humans and dogs. Vet. J. 2003, 165, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Crockford, C.; Wittig, R.M.; Langergraber, K.; Ziegler, T.E.; Zuberbühler, K.; Deschner, T. Urinary oxytocin and social bonding in related and unrelated wild chimpanzees. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122765. [Google Scholar] [CrossRef] [Green Version]
- Wittig, R.M.; Crockford, C.; Deschner, T.; Langergraber, K.E.; Ziegler, T.E.; Zuberbühler, K. Food sharing is linked to urinary oxytocin levels and bonding in related and unrelated wild chimpanzees. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133096. [Google Scholar] [CrossRef] [Green Version]
- Mora-Medina, P.; Orihuela-Trujillo, A.; Arch-Tirado, E.; Roldan-Santiago, P.; Terrazas, A.; Mota-Rojas, D. Sensory factors involved in mother-young bonding in sheep: A review. Vet. Med. 2016, 61, 595–611. [Google Scholar] [CrossRef] [Green Version]
- Bienboire-Frosini, C.; Marcet-Rius, M.; Orihuela, A.; Domínguez-Oliva, A.; Mora-Medina, P.; Olmos-Hernández, A.; Casas-Alvarado, A.; Mota-Rojas, D. Mother–young bonding: Neurobiological aspects and maternal biochemical signaling in altricial domesticated mammals. Animals 2023, 13, 532. [Google Scholar] [CrossRef]
- Kent, J.P. The cow–calf relationship: From maternal responsiveness to the maternal bond and the possibilities for fostering. J. Dairy Res. 2020, 87, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Bragaglio, A.; Braghieri, A.; Napolitano, F.; Domínguez-Oliva, A.; Mora-Medina, P.; Álvarez-Macías, A.; De Rosa, G.; Pacelli, C.; José, N.; et al. Dairy buffalo behavior: Calving, imprinting and allosuckling. Animals 2022, 12, 2899. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, F.; Braghieri, A.; Bragaglio, A.; Rodríguez-González, D.; Mora-Medina, P.; Ghezzi, M.D.; Álvarez-Macías, A.; Lendez, P.A.; Sabia, E.; Domínguez-Oliva, A.; et al. Neurophysiology of milk ejection and prestimulation in dairy buffaloes. Animals 2022, 12, 2649. [Google Scholar] [CrossRef] [PubMed]
- Ybarra-Navarro, N.T. Evaluation of Oxytocin Pharmacokinetic: Pharmacodynamic Profile and Establishment of Its Cardiomyogenic Potential in Swine. Ph.D. Thesis, Faculté de Médecine Vétérinaire, Université de Montréal, Saint-Hyacinthe, QC, Canada, 2010. [Google Scholar]
- Lippert, T.H.; Mueck, A.O.; Seeger, H.; Pfaff, A. Effects of oxytocin outside pregnancy. Horm. Res. Paediatr. 2003, 60, 262–271. [Google Scholar] [CrossRef]
- Andriolo, A.; Schmidek, W. Suckling behaviour in water buffalo (Bubalus bubalis). Rev. Etol. 2001, 3, 129–136. [Google Scholar]
- Mota-Rojas, D.; Velarde, A.; Marcet-Rius, M.; Orihuela, A.; Bragaglio, A.; Hernández-Ávalos, I.; Casas-Alvarado, A.; Domínguez-Oliva, A.; Whittaker, A.L. Analgesia during parturition in domestic animals: Perspectives and controversies on its use. Animals 2022, 12, 2686. [Google Scholar] [CrossRef]
- Brunton, P.J. Endogenous opioid signalling in the brain during pregnancy and lactation. Cell Tissue Res. 2019, 375, 69–83. [Google Scholar] [CrossRef]
- Sheldrick, E.L.; Flint, A.P.F. Post-Translational processing of oxytocin-neurophysin prohormone in the ovine corpus luteum: Activity of peptidyl glycine α-amidating mono-oxygenase and concentrations of its cofactor, ascorbic acid. J. Endocrinol. 1989, 122, 313–322. [Google Scholar] [CrossRef]
- Schams, D.; Kruip, T.A.M.; Koll, R. Oxytocin determination in steroid producing tissues and in vitro production in ovarian follicles. Eur. J. Endocrinol. 1985, 109, 530–536. [Google Scholar] [CrossRef]
- Knickerbocker, J.J.; Sawyer, H.R.; Amann, R.P.; Tekpetey, F.R.; Niswender, G.D. Evidence for the presence of oxytocin in the ovine epididymis. Biol. Reprod. 1988, 39, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Breton, C.; Zingg, H.H. Expression and region-specific regulation of the oxytocin receptor gene in rat brain. Endocrinology 1997, 138, 1857–1862. [Google Scholar] [CrossRef] [PubMed]
- Shughrue, P.J.; Dellovade, T.L.; Merchenthaler, I. Chapter 2 Estrogen modulates oxytocin gene expression in regions of the rat supraoptic and paraventricular nuclei that contain estrogen receptor-β. Prog. Brain Res. 2002, 139, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Patisaul, H.B.; Scordalakes, E.M.; Young, L.J.; Rissman, E.F. Oxytocin, but not oxytocin receptor, is regulated by oestrogen receptor β in the female mouse hypothalamus. J. Neuroendocrinol. 2003, 15, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Murakami, G.; Hunter, R.G.; Fontaine, C.; Ribeiro, A.; Pfaff, D. Relationships among estrogen receptor, oxytocin and vasopressin gene expression and social interaction in male mice. Eur. J. Neurosci. 2011, 34, 469–477. [Google Scholar] [CrossRef]
- Acevedo-Rodriguez, A.; Mani, S.K.; Handa, R.J. Oxytocin and estrogen receptor β in the brain: An overview. Front. Endocrinol. 2015, 6, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowski, M.; Wang, D.; Hajjar, F.; Mukaddam-Daher, S.; McCann, S.M.; Gutkowska, J. Oxytocin and its receptors are synthesized in the rat vasculature. Proc. Natl. Acad. Sci. USA 2000, 97, 6207–6211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankowski, M.; Hajjar, F.; Al Kawas, S.; Mukaddam-Daher, S.; Hoffman, G.; Mccann, S.M.; Gutkowska, J. Rat Heart: A site of oxytocin production and action. Proc. Natl. Acad. Sci. USA 1998, 95, 14558–14563. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.J.; Johnstone, H.; Brunton, P.; Russell, J. Sex-Steroid induction of endogenous opioid inhibition on oxytocin secretory responses to stress. J. Neuroendocrinol. 2001, 12, 343–350. [Google Scholar] [CrossRef]
- Brunton, P.J.; Russell, J.A.; Hirst, J.J. Allopregnanolone in the brain: Protecting pregnancy and birth outcomes. Prog. Neurobiol. 2014, 113, 106–136. [Google Scholar] [CrossRef] [Green Version]
- Ochedalski, T.; Subburaju, S.; Wynn, P.C.; Aguilera, G. Interaction between oestrogen and oxytocin on hypothalamic-pituitary-adrenal axis activity. J. Neuroendocrinol. 2007, 19, 189–197. [Google Scholar] [CrossRef]
- Neumann, I.D.; Krömer, S.A.; Toschi, N.; Ebner, K. Brain oxytocin inhibits the (re)activity of the hypothalamo–pituitary–adrenal axis in male rats: Involvement of hypothalamic and limbic brain regions. Regul. Pept. 2000, 96, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Tančin, V.; Bruckmaier, R.M. Factors affecting milk ejection and removal during milking and suckling of dairy cows. Vet. Med. 2001, 46, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.J.; Dye, S.; Leng, G.; Russell, J.A.; Bicknell, R.J. Endogenous opioid regulation of oxytocin secretion through pregnancy in the rat. J. Neuroendocrinol. 1993, 5, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Stack, E.C.; Balakrishnan, R.; Numan, M.J.; Numan, M. A functional neuroanatomical investigation of the role of the medial preoptic area in neural circuits regulating maternal behavior. Behav. Brain Res. 2002, 131, 17–36. [Google Scholar] [CrossRef] [PubMed]
- Srisawat, R.; Ludwig, M.; Bull, P.M.; Douglas, A.J.; Russell, J.A.; Leng, G. Nitric oxide and the oxytocin system in pregnancy. J. Neurosci. 2000, 20, 6721–6727. [Google Scholar] [CrossRef] [Green Version]
- Purohit, G. Parturition in domestic animals: A review. WebmedCentral 2010, 1, WMC00748. [Google Scholar] [CrossRef]
- Lévy, F. Neuroendocrine control of maternal behavior in non-human and human mammals. Ann. Endocrinol. 2016, 77, 114–125. [Google Scholar] [CrossRef]
- Blanks, A.M.; Thornton, S. The role of oxytocin in parturition. BJOG Int. J. Obstet. Gynaecol. 2003, 110, 46–51. [Google Scholar] [CrossRef]
- Fusi, J.; Veronesi, M. Canine parturition: What is known about the hormonal setting? Domest. Anim. Endocrinol. 2022, 78, 106687. [Google Scholar] [CrossRef]
- Kowalewski, M.P.; Tavares Pereira, M.; Kazemian, A. Canine conceptus-maternal communication during maintenance and termination of pregnancy, including the role of species-specific decidualization. Theriogenology 2020, 150, 329–338. [Google Scholar] [CrossRef]
- Olcese, J.; Beesley, S. Clinical significance of melatonin receptors in the human myometrium. Fertil. Steril. 2014, 102, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Burnes, J.; Muns, R.; Barrios-García, H.; Villanueva-García, D.; Domínguez-Oliva, A.; Mota-Rojas, D. Parturition in mammals: Animal models, pain and distress. Animals 2021, 11, 2960. [Google Scholar] [CrossRef] [PubMed]
- Vallet, J.L.; Miles, J.R.; Brown-Brandl, T.M.; Nienaber, J.A. Proportion of the litter farrowed, litter size, and progesterone and estradiol effects on piglet birth intervals and stillbirths. Anim. Reprod. Sci. 2010, 119, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliviero, C. Sucessful Farrowing in Sows. Ph.D. Thesis, University of Helsinki, Helsinki, Finland, 2010. [Google Scholar]
- Contreras, J.E.; Sáez, J.C.; Bukauskas, F.F.; Bennett, M.V.L. Gating and regulation of connexin 43 (Cx43) hemichannels. Proc. Natl. Acad. Sci. USA 2003, 100, 11388–11393. [Google Scholar] [CrossRef] [Green Version]
- Chan, L.Y.-S.; Fu, L.; Leung, T.N.; Wong, S.F.; Lau, T.K. Obstetric outcomes after cervical ripening by multiple doses of vaginal prostaglandin E2. Acta Obstet. Gynecol. Scand. 2004, 2, 70–74. [Google Scholar]
- Papatsonis, D.; Flenady, V.; Cole, S.; Liley, H. Oxytocin receptor antagonists for inhibiting preterm labour. In Cochrane Database of Systematic Reviews; Papatsonis, D., Ed.; John Wiley & Sons, Ltd.: Chichester, UK, 2005. [Google Scholar]
- Kimura, T.; Tanizawa, O.; Mori, K.; Brownstein, M.J.; Okayama, H. Structure and expression of a human oxytocin receptor. Nature 1992, 356, 526–529. [Google Scholar] [CrossRef]
- Ou, C.-W.; Chen, Z.-Q.; Qi, S.; Lye, S.J. Increased expression of the rat myometrial oxytocin receptor messenger ribonucleic acid during labor requires both mechanical and hormonal signals. Biol. Reprod. 1998, 59, 1055–1061. [Google Scholar] [CrossRef] [Green Version]
- Meier, S.; Lau, T.M.; Jenkin, G.; Fairclough, R.J. Oxytocin-induced prostaglandin F2α release and endometrial oxytocin receptor concentrations throughout pregnancy in ewes. J. Reprod. Fertil. 1995, 103, 233–238. [Google Scholar]
- Fuchs, A.R.; Behrens, O.; Helmer, H.; Liu, C.-H.; Barros, C.M.; Fields, M.J. Oxytocin and vasopressin receptors in bovine endometrium and myometrium during the estrous cycle and early pregnancy. Endocrinology 1990, 127, 629–636. [Google Scholar] [CrossRef]
- Fuchs, A.-R.; Ivell, R.; Ganz, N.; Fields, M.J.; Gimenez, T. Secretion of oxytocin in pregnant and parturient cows: Corpus luteum may contribute to plasma oxytocin at term. Biol. Reprod. 2001, 65, 1135–1141. [Google Scholar] [CrossRef]
- Veiga, G.A.L.; Milazzotto, M.P.; Nichi, M.; Lúcio, C.F.; Silva, L.C.G.; Vannucchi, C.I. Correlation of oxytocin (OTR) and estrogen receptor (ER) MRNA in the canine placenta with the detected circulating levels of oxytocin and estrogen during pregnancy and parturition. Am. J. Anim. Vet. Sci. 2016, 11, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Lundin-Schiller, S.; Kreider, D.L.; Rorie, R.W.; Hardesty, D.; Mitchell, M.D.; Koike, T.I. Characterization of porcine endometrial, myometrial, and mammary oxytocin binding sites during gestation and labor. Biol. Reprod. 1996, 55, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Norwitz, E.R.; Robinson, J.N.; Challis, J.R. The control of labor. N. Engl. J. Med. 1999, 341, 660–666. [Google Scholar] [PubMed]
- Masoudi, R. Effect of estradiol and oxytocin on ovine cervical relaxation. Afr. J. Biotechnol. 2012, 11, 2803–2806. [Google Scholar] [CrossRef]
- Kustritz, M.V.R. Reproductive behavior of small animals. Theriogenology 2005, 64, 734–746. [Google Scholar] [CrossRef]
- Whitman, D.C.; Albers, H.E. Role of oxytocin in the hypothalamic regulation of sexual receptivity in hamsters. Brain Res. 1995, 680, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, C.; Heinonen, M.; Valros, A.; Hälli, O.; Peltoniemi, O.A.T. Effect of the environment on the physiology of the sow during late pregnancy, farrowing and early lactation. Anim. Reprod. Sci. 2008, 105, 365–377. [Google Scholar] [CrossRef]
- Yun, J.; Swan, K.M.; Vienola, K.; Kim, Y.Y.; Oliviero, C.; Peltoniemi, O.A.T.; Valros, A. Farrowing environment has an impact on sow metabolic status and piglet colostrum intake in early lactation. Livest. Sci. 2014, 163, 120–125. [Google Scholar] [CrossRef]
- Alonso-Spilsbury, M.; Mota-Rojas, D.; Martı́nez-Burnes, J.; Arch, E.; López Mayagoitia, A.; Ramı́rez-Necoechea, R.; Olmos, A.; Trujillo, M.E. Use of Oxytocin in penned sows and its effect on fetal intra-partum asphyxia. Anim. Reprod. Sci. 2004, 84, 157–167. [Google Scholar] [CrossRef]
- Oliviero, C.; Heinonen, M.; Valros, A.; Peltoniemi, O. Environmental and sow-related factors affecting the duration of farrowing. Anim. Reprod. Sci. 2010, 119, 85–91. [Google Scholar] [CrossRef]
- van Dijk, A.J.; van Rens, B.T.T.M.; van der Lende, T.; Taverne, M.A.M. Factors affecting duration of the expulsive stage of parturition and piglet birth intervals in sows with uncomplicated, spontaneous farrowings. Theriogenology 2005, 64, 1573–1590. [Google Scholar] [CrossRef] [PubMed]
- Darvelid, A.W.; Linde-Forsberg, C. Dystocia in the bitch: A retrospective study of 182 cases. J. Small Anim. Pract. 1994, 35, 402–407. [Google Scholar] [CrossRef]
- Davidson, A.P. Primary uterine inertia in four labrador bitches. J. Am. Anim. Hosp. Assoc. 2011, 47, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Jungmann, C.; Houghton, C.G.; Nielsen, F.G.; Packeiser, E.-M.; Körber, H.; Reichler, I.M.; Balogh, O.; Goericke-Pesch, S. Involvement of Oxytocin and Progesterone Receptor Expression in the Etiology of Canine Uterine Inertia. Int. J. Mol. Sci. 2022, 23, 13601. [Google Scholar] [CrossRef]
- Nevard, R.P.; Pant, S.D.; Broster, J.C.; Norman, S.T.; Stephen, C.P. Maternal behavior in beef cattle: The physiology, assessment and future directions—A review. Vet. Sci. 2022, 10, 10. [Google Scholar] [CrossRef]
- Coria-Avila, G.A.; Herrera-Covarrubias, D.; García, L.I.; Toledo, R.; Hernández, M.E.; Paredes-Ramos, P.; Corona-Morales, A.A.; Manzo, J. Neurobiology of maternal behavior in nonhuman mammals: Acceptance, recognition, motivation, and rejection. Animals 2022, 12, 3589. [Google Scholar] [CrossRef]
- Geburt, K.; Friedrich, M.; Piechotta, M.; Gauly, M.; König von Borstel, U. Validity of physiological biomarkers for maternal behavior in cows—A comparison of beef and dairy cattle. Physiol. Behav. 2015, 139, 361–368. [Google Scholar] [CrossRef]
- Sanson, A.; Bosch, O.J. Dysfunctions of brain oxytocin signaling: Implications for poor mothering. Neuropharmacology 2022, 211, 109049. [Google Scholar] [CrossRef]
- Orihuela, A.; Mota-Rojas, D.; Strappini, A.; Serrapica, F.; Braghieri, A.; Mora-Medina, P.; Napolitano, F. Neurophysiological mechanisms of cow–calf bonding in buffalo and other farm animals. Animals 2021, 11, 1968. [Google Scholar] [CrossRef]
- Algers, B.; Uvnäs-Moberg, K. Maternal behavior in pigs. Horm. Behav. 2007, 52, 78–85. [Google Scholar] [CrossRef]
- Hall, S.A.; Farish, M.; Coe, J.; Baker, E.; Camerlink, I.; Lawrence, A.B.; Baxter, E.M. Minimally invasive biomarkers to detect maternal physiological status in sow saliva and milk. Animal 2021, 15, 100369. [Google Scholar] [CrossRef] [PubMed]
- Rosvold, E.M.; Newberry, R.C.; Andersen, I.L. Early Mother-young interactions in domestic sows–nest-building material increases maternal investment. Appl. Anim. Behav. Sci. 2019, 219, 104837. [Google Scholar] [CrossRef]
- López-Arjona, M.; Mainau, E.; Navarro, E.; Contreras-Aguilar, M.D.; Escribano, D.; Mateo, S.V.; Manteca, X.; Cerón, J.J.; Martínez-Subiela, S. Oxytocin in bovine saliva: Validation of two assays and changes in parturition and at weaning. BMC Vet. Res. 2021, 17, 140. [Google Scholar] [CrossRef]
- Lévy, F. The onset of maternal behavior in sheep and goats: Endocrine, sensory, neural, and experiential mechanisms. In Patterns of Parental Behavior. Advances in Neurobiology; González-Mariscal, G., Ed.; Springer: Cham, Switzerland, 2022; pp. 79–117. [Google Scholar]
- Lupoli, B.; Johansson, B.; Uvnäs-Moberg, K.; Svennersten-Sjaunja, K. Effect of suckling on the release of oxytocin, prolactin, cortisol, gastrin, cholecystokinin, somatostatin and insulin in dairy cows and their calves. J. Dairy Res. 2001, 68, 175–187. [Google Scholar] [CrossRef]
- Silveira, P.A.; Spoon, R.A.; Ryan, D.P.; Williams, G.L. Evidence for maternal behavior as a requisite link in suckling-mediated anovulation in cows. Biol. Reprod. 1993, 49, 1338–1346. [Google Scholar] [CrossRef]
- Yun, J.; Swan, K.-M.; Farmer, C.; Oliviero, C.; Peltoniemi, O.; Valros, A. Prepartum nest-building has an impact on postpartum nursing performance and maternal behaviour in early lactating sows. Appl. Anim. Behav. Sci. 2014, 160, 31–37. [Google Scholar] [CrossRef]
- Wang, H.; Han, C.; Li, M.; Li, F.; Yang, Y.; Wang, Z.; Lv, S. Effects of parity, litter size and lamb sex on maternal behavior of small Tail Han sheep and their neuroendocrine mechanisms. Small Rumin. Res. 2021, 202, 106451. [Google Scholar] [CrossRef]
- Santos, N.R.; Beck, A.; Fontbonne, A. A review of maternal behaviour in dogs and potential areas for further research. J. Small Anim. Pract. 2020, 61, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.H.; Grieb, Z.A. Species differences in the effect of oxytocin on maternal behavior: A model incorporating the potential for allomaternal contributions. Front. Neuroendocrinol. 2022, 65, 100996. [Google Scholar] [CrossRef]
- Bridges, R.S. Neuroendocrine regulation of maternal behavior. Front. Neuroendocrinol. 2015, 36, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Mota-Rojas, D.; Marcet-Rius, M.; Freitas-de-Melo, A.; Muns, R.; Mora-Medina, P.; Domínguez-Oliva, A.; Orihuela, A. Allonursing in wild and farm animals: Biological and physiological foundations and explanatory hypotheses. Animals 2021, 11, 3092. [Google Scholar] [CrossRef] [PubMed]
- Orihuela, A.; Pérez-Torres, L.I.; Ungerfeld, R. Evidence of cooperative calves’ care and providers’ characteristics in zebu cattle (Bos indicus) raised under extensive conditions. Trop. Anim. Health Prod. 2021, 53, 143. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.L.; Gazal, O.S.; Leshin, L.S.; Stanko, R.L.; Anderson, L.L. Physiological regulation of maternal behavior in heifers: Roles of genital stimulation, intracerebral oxytocin release, and ovarian steroids. Biol. Reprod. 2001, 65, 295–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poindron, P.; Lévy, F.; Keller, M. Maternal responsiveness and maternal selectivity in domestic sheep and goats: The two facets of maternal attachment. Dev. Psychobiol. 2007, 49, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Keverne, B.; Kendrick, K.M. Maternal behaviour in sheep and its neuroendocrine regulation. Acta Paediatr. 1994, 83, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, K.M.; Keverne, E.B.; Hinton, M.R.; Goode, J.A. Cerebrospinal fluid and plasma concentrations of oxytocin and vasopressin during parturition and vaginocervical stimulation in the sheep. Brain Res. Bull. 1991, 26, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Valros, A.; Rundgren, M.; Špinka, M.; Saloniemi, H.; Hultén, F.; Uvnäs-Moberg, K.; Tománek, M.; Krejcί, P.; Algers, B. Oxytocin, prolactin and somatostatin in lactating sows: Associations with mobilisation of body resources and maternal behaviour. Livest. Prod. Sci. 2004, 85, 3–13. [Google Scholar] [CrossRef]
- Wang, C.; Han, Q.; Liu, R.; Ji, W.; Bi, Y.; Wen, P.; Yi, R.; Zhao, P.; Bao, J.; Liu, H. Equipping farrowing pens with straw improves maternal behavior and physiology of min-pig hybrid sows. Animals 2020, 10, 105. [Google Scholar] [CrossRef] [Green Version]
- Kenkel, W.M.; Paredes, J.; Yee, J.R.; Pournajafi-Nazarloo, H.; Bales, K.L.; Carter, C.S. Neuroendocrine and behavioural responses to exposure to an infant in male prairie Voles. J. Neuroendocrinol. 2012, 24, 874–886. [Google Scholar] [CrossRef]
- Olazábal, D.E.; Young, L.J. Oxytocin receptors in the nucleus accumbens facilitate “spontaneous” maternal behavior in adult female prairie voles. Neuroscience 2006, 141, 559–568. [Google Scholar] [CrossRef]
- Robinson, K.J.; Twiss, S.D.; Hazon, N.; Pomeroy, P.P. Maternal oxytocin is linked to close mother-infant proximity in grey seals (Halichoerus grypus). PLoS ONE 2015, 10, e0144577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kockaya, M.; Ercan, N.; Demirbas, Y.S.; Da Graça Pereira, G. Serum oxytocin and lipid levels of dogs with maternal cannibalism. J. Vet. Behav. 2018, 27, 23–26. [Google Scholar] [CrossRef]
- Kendrick, K.M.; Keverne, E.B. Importance of progesterone and estrogen priming for the induction of maternal behavior by vaginocervical stimulation in sheep: Effects of maternal experience. Physiol. Behav. 1991, 49, 745–750. [Google Scholar] [CrossRef]
- McLean, K.; Lawrence, A.; Petherick, J.; Deans, L.; Chirnside, J.; Vaughan, A.; Nielsen, B.; Webb, R. Investigation of the relationship between farrowing environment, sex steroid concentrations and maternal aggression in gilts. Anim. Reprod. Sci. 1998, 50, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Bosch, O.J.; Neumann, I.D. Both oxytocin and vasopressin are mediators of maternal care and aggression in rodents: From central release to sites of action. Horm. Behav. 2012, 61, 293–303. [Google Scholar] [CrossRef]
- Hess, E.H. HESS: The natural history of imprinting. Ann. N. Y. Acad. Sci. 1972, 193, 124–136. [Google Scholar] [CrossRef]
- Ohki-Hamazaki, H. Neurobiology of imprinting. Brain Nerve 2012, 64, 657–664. [Google Scholar] [CrossRef]
- Hess, E.H. Imprinting. Science 1959, 130, 133–141. [Google Scholar] [CrossRef]
- Bridges, R.S. Long-term alterations in neural and endocrine processes induced by motherhood in mammals. Horm. Behav. 2016, 77, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Kim, P. Human maternal brain plasticity: Adaptation to parenting. New Dir. Child Adolesc. Dev. 2016, 2016, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Kim, P.; Strathearn, L.; Swain, J.E. The maternal brain and its plasticity in humans. Horm. Behav. 2016, 77, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Pawluski, J.L.; Lambert, K.G.; Kinsley, C.H. Neuroplasticity in the maternal hippocampus: Relation to cognition and effects of repeated stress. Horm. Behav. 2016, 77, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R. Suckling, milk, and the development of preferences toward maternal cues by neonates: From early learning to filial attachment? In Advances in the Study of Behavior; Elsevier: Amsterdam, The Netherlands, 2006; Volume 36, pp. 1–58. [Google Scholar]
- Mota-Rojas, D.; Bienboire-Frosini, C.; Marcet-Rius, M.; Domínguez-Oliva, A.; Mora-Medina, P.; Lezama-García, K.; Orihuela, A. Mother-young bond in non-human mammals: Neonatal communication pathways and neurobiological basis. Front. Psychol. 2022, 13, 1064444. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Napolitano, F.; Orihuela, A.; Serrapica, F.; Olmos-Hernández, A.; Martínez-Burnes, J.; De Rosa, G. Behavior and welfare of dairy buffaloes: Calving, milking, and weaning. In Biotechnological Applications in Buffalo Research; Chauhan, M., Selokar, N., Eds.; Springer: Singapore, 2022; pp. 97–119. [Google Scholar]
- Mora-Medina, P.; Napolitano, F.; Mota-Rojas, D.; Berdugo-Gutiérrez, J.; Ruiz-Buitrago, J.; Guerrero-Legarreta, I. Imprinting, sucking and allosucking behaviors in buffalo calves. J. Buffalo Sci. 2018, 7, 49–57. [Google Scholar] [CrossRef]
- Orihuela, A.; Mota-Rojas, D.; Urgerfeld, R.; De Rosa, G.; Mora-Medina, P.; Braghieri, A.; Lezama, K.; Domínguez, A.; Napolitano, F. Imprinting in the buffalo and other farm animals: Neurophysiological mechanisms. In El Búfalo de Agua en Latinoamérica. Hallazgos Recientes; Napolitano, F., Mota-Rojas, D., Guerrero-Legarreta, I., Orihuela, A., Eds.; BM Editores: Ciudad de México, México, 2020; pp. 923–958. Available online: https://www.lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 5 June 2020).
- Bolhuis, J.J. Early learning and the development of filial preferences in the chick. Behav. Brain Res. 1999, 98, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Glatzle, M.; Hoops, M.; Kauffold, J.; Seeger, J.; Fietz, S.A. Development of deep and upper neuronal layers in the domestic cat, sheep and pig neocortex. Anat. Histol. Embryol. 2017, 46, 397–404. [Google Scholar] [CrossRef]
- Olazábal, D.E. Role of oxytocin in parental behaviour. J. Neuroendocrinol. 2018, 30, e12594. [Google Scholar] [CrossRef]
- Yoshihara, C.; Numan, M.; Kuroda, K.O. Oxytocin and parental behaviors. In Current Topics in Behavioral Neurosciences; Springer: Berlin/Heidelberg, Germany, 2017; Volume 35, pp. 119–153. [Google Scholar]
- Muir, G.D. Early ontogeny of locomotor behaviour: A comparison between altricial and precocial animals. Brain Res. Bull. 2000, 53, 719–726. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Aoki, N.; Kitajima, T.; Iikubo, E.; Katagiri, S.; Matsushima, T.; Homma, K.J. Thyroid hormone determines the start of the sensitive period of imprinting and primes later learning. Nat. Commun. 2012, 3, 1081. [Google Scholar] [CrossRef] [Green Version]
- Lemche, E. Research evidence from studies on filial imprinting, attachment, and early life stress: A new route for scientific integration. Acta Ethologica 2020, 23, 127–133. [Google Scholar] [CrossRef]
- Dwyer, C.M. Maternal behaviour and lamb survival: From neuroendocrinology to practical application. Animal 2014, 8, 102–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévy, F.; Keller, M. Olfactory mediation of maternal behavior in selected mammalian species. Behav. Brain Res. 2009, 200, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Pissonnier, D.; Thiery, J.C.; Fabre-Nys, C.; Poindron, P.; Keverne, E.B. The Importance of olfactory bulb noradrenalin for maternal recognition in sheep. Physiol. Behav. 1985, 35, 361–363. [Google Scholar] [CrossRef]
- Corona, R.; Meurisse, M.; Cornilleau, F.; Moussu, C.; Keller, M.; Lévy, F. Disruption of adult olfactory neurogenesis induces deficits in maternal behavior in sheep. Behav. Brain Res. 2018, 347, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.; Meurisse, M.; Poindron, P.; Nowak, R.; Ferreira, G.; Shayit, M.; Lévy, F. Maternal experience influences the establishment of visual/auditory, but not olfactory recognition of the newborn lamb by ewes at parturition. Dev. Psychobiol. 2003, 43, 167–176. [Google Scholar] [CrossRef]
- Kojima, S.; Stewart, R.A.; Demas, G.E.; Alberts, J.R. Maternal contact differentially modulates central and peripheral oxytocin in rat pups during a brief regime of mother-pup interaction that induces a filial huddling preference. J. Neuroendocrinol. 2012, 24, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Castro-Sierra, E.; de León, F.C.P.; Domínguez, L.F.G.; Rivera, A.P. Neurotransmisores del sistema límbico. hipocampo, gaba y memoria. primera parte. Salud Ment. 2007, 30, 7–15. [Google Scholar]
- Nowak, R.; Porter, R.H.; Lévy, F.; Orgeur, P.; Schaal, B. Role of mother-young interactions in the survival of offspring in domestic mammals. Rev. Reprod. 2000, 5, 153–163. [Google Scholar] [CrossRef]
- Nowak, R.; Keller, M.; Lévy, F. Mother-Young Relationships in Sheep: A model for a multidisciplinary approach of the study of attachment in mammals. J. Neuroendocrinol. 2011, 23, 1042–1053. [Google Scholar] [CrossRef]
- Tsingotjidou, A.S.; Papadopoulos, G.C. The milk-ejection reflex in the sheep: An anatomical study on the afferent pathway. Anat. Histol. Embryol. 2008, 37, 245–250. [Google Scholar] [CrossRef]
- Dûidić, A. Studies on Milk Ejection and Milk Removal during Machine Milking in Different Species. Ph.D. Thesis, Technische Universität München, München, Germany, 2004; pp. 1–23. [Google Scholar]
- Ralph, C.R.; Tilbrook, A.J. The hypothalamo-pituitary-adrenal (HPA) axis in sheep is attenuated during lactation in response to psychosocial and predator stress. Domest. Anim. Endocrinol. 2016, 55, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Antonijevic, I.A.; Douglas, A.J.; Dye, S.; Bicknell, R.J.; Leng, G.; Russell, J.A. Oxytocin antagonists delay the initiation of parturition and prolong its active phase in rats. J. Endocrinol. 1995, 145, 97–103. [Google Scholar] [CrossRef]
- Nishimori, K.; Young, L.J.; Guo, Q.; Wang, Z.; Insel, T.R.; Matzuk, M.M. Oxytocin is required for nursing but is not essential for parturition or reproductive behavior. Proc. Natl. Acad. Sci. USA 1996, 93, 11699–11704. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, M.; Sabatier, N.; Bull, P.M.; Landgraf, R.; Dayanithi, G.; Leng, G. Intracellular calcium stores regulate activity-dependent neuropeptide release from dendrites. Nature 2002, 418, 85–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.H. Magnocellular neurons and posterior pituitary function. In Comprehensive Physiology; Wiley: Hoboken, NJ, USA, 2016; Volume 6, pp. 1701–1741. [Google Scholar]
- Meddle, S.L.; Bishop, V.R.; Gkoumassi, E.; van Leeuwen, F.W.; Douglas, A.J. Dynamic changes in oxytocin receptor expression and activation at parturition in the rat brain. Endocrinology 2007, 148, 5095–5104. [Google Scholar] [CrossRef] [PubMed]
- Bruckmaier, R.M.; Wellnitz, O. Induction of milk ejection and milk removal in different production systems. J. Anim. Sci. 2008, 86, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Boselli, C.; Campagna, M.C.; Amatiste, S.; Rosati, R.; Borghese, A. Pre-Stimulation effects on teat anatomy and milk flow curves in mediterranean italian buffalo cows. J. Anim. Vet. Adv. 2014, 13, 912–916. [Google Scholar] [CrossRef]
- Slattery, D.A.; Neumann, I.D. No stress please! Mechanisms of stress hyporesponsiveness of the maternal brain. J. Physiol. 2008, 586, 377–385. [Google Scholar] [CrossRef]
- Tilbrook, A.J.; Turner, A.I.; Ibbott, M.D.; Clarke, I.J. Activation of the hypothalamo-pituitary-adrenal axis by isolation and restraint stress during lactation in ewes: Effect of the presence of the lamb and suckling. Endocrinology 2006, 147, 3501–3509. [Google Scholar] [CrossRef] [Green Version]
- Wagner, B.K.; Relling, A.E.; Kieffer, J.D.; Parker, A.J. Brief communication: Plasma cortisol concentration is affected by lactation, but not intra-nasal oxytocin treatment, in beef cows. PLoS ONE 2021, 16, e0249323. [Google Scholar] [CrossRef]
- Cook, C.J. Oxytocin and prolactin suppress cortisol responses to acute stress in both lactating and non-lactating sheep. J. Dairy Res. 1997, 64, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Ellendorff, F.; Schams, D. Characteristics of milk ejection, associated intramammary pressure changes and oxytocin release in the mare. J. Endocrinol. 1988, 119, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Polikarpus, A.; Napolitano, F.; Grasso, F.; Di Palo, R.; Zicarelli, F.; Arney, D.; De Rosa, G. Effect of pre-partum habituation to milking routine on behaviour and lactation performance of buffalo heifers. Appl. Anim. Behav. Sci. 2014, 161, 1–6. [Google Scholar] [CrossRef]
- Polikarpus, A.; Grasso, F.; Pacelli, C.; Napolitano, F.; De Rosa, G. Milking behaviour of buffalo cows: Entrance order and side preference in the milking parlour. J. Dairy Res. 2014, 81, 24–29. [Google Scholar] [CrossRef]
- Espinosa, Y.; Ponce, P.; Capdevila, J. Efecto de la estimulación con bucerro, oxitocina y manual sobre los indicarores de ordeño en búfalas. Rev. Salud Anim. 2011, 33, 90–96. [Google Scholar]
- Boselli, C.; De Marchi, M.; Costa, A.; Borghese, A. Study of milkability and its relation with milk yield and somatic cell in mediterranean italian water buffalo. Front. Vet. Sci. 2020, 7, 432. [Google Scholar] [CrossRef]
- Singh, M.; Aggarwal, A. Effect of oxytocin administration on certain minerals in the milk of buffaloes (Bubalus bubalis). Asian-Australas. J. Anim. Sci. 2001, 14, 1523–1526. [Google Scholar] [CrossRef]
- Keeling, L.J.; De Oliveira, D.; Rustas, B.-O.; Keeling, L.J.; De Oliveira, D.; Rustas, B.-O. Use of mechanical rotating brushes in dairy cows—A potential proxy for performance and welfare. Precis. Dairy Farming 2016, 9, 343–347. [Google Scholar]
- Armstrong, W.E.; Hatton, G.I. The puzzle of pulsatile oxytocin secretion during lactation: Some new pieces. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R26–R28. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, Y.; Onaka, T. Roles of oxytocin in stress responses, allostasis and resilience. Int. J. Mol. Sci. 2021, 23, 150. [Google Scholar] [CrossRef]
- Yukinaga, H.; Hagihara, M.; Tsujimoto, K.; Chiang, H.-L.; Kato, S.; Kobayashi, K.; Miyamichi, K. Recording and manipulation of the maternal oxytocin neural activities in mice. Curr. Biol. 2022, 32, 3821–3829. [Google Scholar] [CrossRef] [PubMed]
- Mačuhová, J.; Tančin, V.; Bruckmaier, R.M. Oxytocin release, milk ejection and milk removal in a multi-box automatic milking system. Livest. Prod. Sci. 2003, 81, 139–147. [Google Scholar] [CrossRef]
- Ciborowska, P.; Michalczuk, M.; Bień, D. The effect of music on livestock: Cattle, poultry and pigs. Animals 2021, 11, 3572. [Google Scholar] [CrossRef]
- Ballou, L.U.; Bleck, J.L.; Bleck, G.T.; Bremel, R.D. The effects of daily oxytocin injections before and after milking on milk production, milk plasmin, and milk composition. J. Dairy Sci. 1993, 76, 1544–1549. [Google Scholar] [CrossRef] [PubMed]
- Hameed, A.; Anjum, F.M.; ur Rehman, Z.; Akhtar, S.; Faraz, A.; Hussain, M.; Ismail, A. Compositional and mineral profile of sahiwal cow milk at various lactation stages as influenced by oxytocin administration. Pak. J. Zool. 2021, 54, 1–7. [Google Scholar] [CrossRef]
- Assad, N.I.; Pandey, A.K.; Sharma, L.M. Oxytocin, Functions, Uses and Abuses: A Brief Review. Theriogenology Insight-Int. J. Reprod. All Anim. 2016, 6, 1–17. [Google Scholar] [CrossRef]
- Borghese, A.; Rasmussen, M.; Thomas, C.S. Milking management of dairy buffalo. Ital. J. Anim. Sci. 2007, 6, 39–50. [Google Scholar] [CrossRef]
- Abecia, J.A.; Palacios, C. Ewes giving birth to female lambs produce more milk than ewes giving birth to male lambs. Ital. J. Anim. Sci. 2018, 17, 736–739. [Google Scholar] [CrossRef]
- Murtaza, S.; Sattar, A.; Ahmad, N.; Ijaz, M.; Omer, T.; Akhtar, M.; Shahzad, M. Long term administration of exogenous oxytocin: Effects on pregnancy rate, and embryonic and fetal losses in Nili-Ravi Buffaloes. J. Anim. Plant Sci. 2020, 30, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Gümen, A.; Keskin, A.; Yilmazbas-Mecitoglu, G.; Karakaya, E.; Cevik, S.; Balci, F. Effects of GnRH, PGF2α and oxytocin treatments on conception rate at the time of artificial insemination in lactating dairy cows. Czech J. Anim. Sci. 2011, 56, 279–283. [Google Scholar] [CrossRef] [Green Version]
- King, M.E.; McKelvey, W.A.C.; Dingwall, W.S.; Matthews, K.P.; Gebbie, F.E.; Mylne, M.J.A.; Stewart, E.; Robinson, J.J. Lambing rates and litter sizes following intrauterine or cervical insemination of frozen/thawed semen with or without oxytocin administration. Theriogenology 2004, 62, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, A. Effect of oxytocin on conception rate in cows. Fırat Univ. J. Health Sci. 2005, 19, 75–78. [Google Scholar]
- Morgan, J.; Fogarty, N.M.; Nicol, H. Oxytocin administration and its effect on ewe milk composition. Asian-Australas. J. Anim. Sci. 2000, 13, 206–208. [Google Scholar]
- Farmer, C.; Lessard, M.; Knight, C.H.; Quesnel, H. Oxytocin injections in the postpartal period affect mammary tight junctions in sows1. J. Anim. Sci. 2017, 95, 3532–3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez, A.; Jiménez, P.; Inoue, K.; Young, L.J.; González-Mariscal, G. Oxytocin antagonist does not disrupt rabbit maternal behavior despite binding to brain oxytocin receptors. J. Neuroendocrinol. 2023, early view, e13236. [Google Scholar] [CrossRef]
- Monks, D.T.; Palanisamy, A. Oxytocin: At birth and beyond. a systematic review of the long-term effects of peripartum oxytocin. Anaesthesia 2021, 76, 1526–1537. [Google Scholar] [CrossRef] [PubMed]
- Osella, M.C.; Cozzi, A.; Spegis, C.; Turille, G.; Barmaz, A.; Lecuelle, C.L.; Teruel, E.; Bienboire-Frosini, C.; Chabaud, C.; Bougrat, L.; et al. The effects of a synthetic analogue of the Bovine Appeasing Pheromone on milk yield and composition in Valdostana dairy cows during the move from winter housing to confined lowland pastures. J. Dairy Res. 2018, 85, 174–177. [Google Scholar] [CrossRef]
- Marnet, P.G.; McKusick, B.C. Regulation of milk ejection and milkability in small ruminants. Livest. Prod. Sci. 2001, 70, 125–133. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mota-Rojas, D.; Marcet-Rius, M.; Domínguez-Oliva, A.; Martínez-Burnes, J.; Lezama-García, K.; Hernández-Ávalos, I.; Rodríguez-González, D.; Bienboire-Frosini, C. The Role of Oxytocin in Domestic Animal’s Maternal Care: Parturition, Bonding, and Lactation. Animals 2023, 13, 1207. https://doi.org/10.3390/ani13071207
Mota-Rojas D, Marcet-Rius M, Domínguez-Oliva A, Martínez-Burnes J, Lezama-García K, Hernández-Ávalos I, Rodríguez-González D, Bienboire-Frosini C. The Role of Oxytocin in Domestic Animal’s Maternal Care: Parturition, Bonding, and Lactation. Animals. 2023; 13(7):1207. https://doi.org/10.3390/ani13071207
Chicago/Turabian StyleMota-Rojas, Daniel, Míriam Marcet-Rius, Adriana Domínguez-Oliva, Julio Martínez-Burnes, Karina Lezama-García, Ismael Hernández-Ávalos, Daniela Rodríguez-González, and Cécile Bienboire-Frosini. 2023. "The Role of Oxytocin in Domestic Animal’s Maternal Care: Parturition, Bonding, and Lactation" Animals 13, no. 7: 1207. https://doi.org/10.3390/ani13071207