
Genetic Parameters for a Weighted Analysis of Survivability in Dairy Cattle
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
Statistical Model
3. Results
3.1. The Least-Squares Method Fixed Effects
3.2. Components of Variance and Genetic Parameters
3.3. Variability of Effects in the Solution
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Punsmann, T.; Distl, O. Length of productive life and longevity in dairy cows: Heritability of traits for longevity and their genetic correlations with milk performance and functional traits. Züchtungskunde 2017, 89, 125–139. [Google Scholar]
- Alvåsen, K.; Dohoo, I.; Roth, A.; Emanuelson, U. Farm characteristics and management routines related to cow longevity: A survey among Swedish dairy farmers. Acta Vet. Scand. 2018, 60, 38. [Google Scholar] [CrossRef] [PubMed]
- Mirhabibi, S.; Kashan, N.; Gharahveysi, S. Genetic evaluation of survival traits in the Holstein dairy cows of Iran. Egypt. J. Vet. Sci. 2018, 49, 71–74. [Google Scholar] [CrossRef]
- Krupová, Z.; Wolfová, M.; Krupa, E.; Přibyl, J.; Zavadilová, L. Claw health and feed efficiency as new selection criteria in the Czech Holstein cattle. Czech J. Anim. Sci. 2018, 63, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Mu, T.; Ma, Y.; Wang, X.; Ma, Y. Analysis of Longevity Traits in Holstein Cattle: A Review. Front. Genet. 2021, 12, 695543. [Google Scholar] [CrossRef]
- Bach, A. Associations between several aspects of heifer development and dairy cow survivability to second lactation. J. Dairy Sci. 2011, 94, 1052–1057. [Google Scholar] [CrossRef] [Green Version]
- Schuster, J.C.; Barkema, H.W.; De Vries, A.; Kelton, D.F.; Orsel, K. Invited review: Academic and applied approach to evaluating longevity in dairy cows. J. Dairy Sci. 2021, 103, 11008–11024. [Google Scholar] [CrossRef]
- Khansefid, M.; Haile-Mariam, M.; Pryce, J.E. Improving the accuracy of predictions for cow survival by multivariate evaluation model. Anim. Prod. Sci. 2021, 61, 1828–1836. [Google Scholar] [CrossRef]
- Beaudeau, F.; Ducrocq, V.; Fourichon, C.; Seegers, H. Effect of disease on length of productive life of French Holstein dairy cows assessed by survival analysis. J. Dairy Sci. 1995, 78, 103–117. [Google Scholar] [CrossRef]
- Berry, D.P.; Harris, B.L.; Winkelman, A.M.; Montgomerie, W. Phenotypic associations between traits other than production and longevity in New Zealand dairy cattle. J. Dairy Sci. 2005, 88, 2962–2974. [Google Scholar] [CrossRef] [Green Version]
- De Vries, A.; Marcondes, M.I. Overview of factors affecting productive lifespan of dairy cows. Animal 2020, 14, 155–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Heide, E.M.; Kamphuis, C.; Veerkamp, R.F.; Athanasiadis, I.N.; Azzopardi, G.; van Pelt, M.L.; Ducro, B.J. Improving predictive performance on survival in dairy cattle using an ensemble learning approach. Comput. Electron. Agric. 2020, 177, 105675. [Google Scholar] [CrossRef]
- Van Pelt, M.L.; Meuwissen, T.H.E.; De Jong, G.; Veerkamp, R.F. Genetic analysis of longevity in Dutch dairy cattle using random regression. J. Dairy Sci. 2015, 98, 4117–4130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steri, R.; Moioli, B.; Catillo, G.; Galli, A.; Buttazzoni, L. Genome-wide association study for longevity in the Holstein cattle population. Animal 2019, 13, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Siatka, K.; Sawa, A.; Krężel-Czopek, S.; Bogucki, M. Longevity of Holstein-Friesian cows and some factors affecting their productive life-a review. Anim. Sci. Pap. Rep. 2020, 38, 107–116. [Google Scholar]
- Adamczyk, K.; Jagusiak, W.; Węglarz, A. Associations between the breeding values of Holstein Friesian bulls and longevity and culling reasons of their daughters. Animal 2021, 15, 5. [Google Scholar] [CrossRef]
- Van Eetvelde, M.; Verdru, K.; de Jong, G.; van Pelt, M.L.; Meesters, M.; Opsomer, G. Researching 100 t cows: An innovative approach to identify intrinsic cows factors associated with a high lifetime milk production. Prev. Vet. Med. 2021, 193, 105392. [Google Scholar] [CrossRef]
- Dallago, G.M.; Wade, K.M.; Cue, R.I.; McClure, J.T.; Lacroix, R.; Pellerin, D.; Vasseur, E. Keeping dairy cows for longer: A critical literature review on dairy cow longevity in high milk-producing countries. Animals 2021, 11, 808. [Google Scholar] [CrossRef]
- Afiani, F.A.; Joezy-Shekalgorabi, S.; Amin-Afshar, M.; Sadeghi, A.A.; Jensen, J. Additive genetic and permanent environmental correlation between different parts of lactation in moderate and cold regions. Czech J. Anim. Sci. 2021, 66, 112–121. [Google Scholar] [CrossRef]
- Krejčová, H.; Přibyl, J.; Přibylová, J.; Štípková, M.; Mielenz, N. Genetic evaluation of daily gains of dual-purpose bulls using a random regression model. Czech J. Anim. Sci. 2008, 53, 227–237. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, T.; Coffey, M.; Mrode, R.; Wall, E. Understanding the genetics of survival in dairy cows. J. Dairy Sci. 2013, 96, 3296–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samoré, A.B.; Rizzi, R.; Rossoni, A.; Bagnato, A. Genetic parameters for functional longevity, type traits, somatic cell scores, milk flow and production in the Italian Brown Swiss. Ital. J. Anim. Sci. 2010, 9, e28. [Google Scholar] [CrossRef]
- Páchová, E.; Zavadilová, L.; Sölkner, J. Genetic evaluation of the length of productive life in Holstein cattle in the Czech Republic. Czech J. Anim. Sci. 2005, 50, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Zavadilová, L.; Štípková, M. Genetic correlations between longevity and conformation traits in the Czech Holstein population. Czech J. Anim. Sci. 2012, 57, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Clasen, J.B.; Norberg, E.; Madsen, P.; Pedersen, J.; Kargo, M. Estimation of genetic parameters and heterosis for longevity in crossbred Danish dairy cattle. J. Dairy Sci. 2017, 100, 6337–6342. [Google Scholar] [CrossRef] [PubMed]
- Ducrocq, V.; Solkner, J. The survival kit (v. 3.0): A Fortran package for the analysis of survival data. In Proceedings of the 6th World Congress on Genetics Applied to Livestock Production, Armidale, Australia, 11–16 January 1998; University of New England: Armidale, Australia, 1998. [Google Scholar]
- Holtsmark, M.; Heringstad, B.; Ødegård, J. Predictive abilities of different statistical models for analysis of survival data in dairy cattle. J. Dairy Sci. 2009, 92, 5730–5738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamrozik, J.; Fatehi, J.; Schaeffer, L.R. Comparison of models for genetic evaluation of survival traits in dairy cattle: A simulation study. J. Anim. Breed. Genet. 2008, 125, 75–83. [Google Scholar] [CrossRef]
- Kern, E.L.; Cobuci, J.A.; Costa, C.N.; McManus, C.M.; Campos, G.S.; Almeida, T.P.; Campos, R.V. Genetic association between herd survival and linear type traits in Holstein cows under tropical conditions. Ital. J. Anim. Sci. 2014, 13, 665–672. [Google Scholar] [CrossRef]
- Heise, J.; Liu, Z.; Stock, K.F.; Rensing, S.; Reinhardt, F.; Simianer, H. The genetic structure of longevity in dairy cows. J. Dairy Sci. 2016, 99, 1253–1265. [Google Scholar] [CrossRef] [Green Version]
- Zavadilová, L.; Němcová, E.; Štípková, M. Effect of type traits on functional longevity of Czech Holstein cows estimated from a Cox proportional hazards model. J. Dairy Sci. 2011, 94, 4090–4099. [Google Scholar] [CrossRef]
- Kyselová, J.; Tichý, L.; Jochová, K. The role of molecular genetics in animal breeding: A minireview. Czech J. Anim. Sci. 2021, 66, 107–111. [Google Scholar] [CrossRef]
- Török, E.; Komlósi, I.; Béri, B.; Füller, I.; Vágó, B.; Posta, J. Principal component analysis of conformation traits in Hungarian Simmental cows. Czech J. Anim. Sci. 2021, 66, 39–45. [Google Scholar] [CrossRef]
- Plemdat. Odhady Plemenné Hodnoty Pro Dlouhověkost. (in Czech). 2020. Available online: https://www.cmsch.cz/plemdat/dokumentace/ (accessed on 29 September 2022).
- Misztal, I.; Tsuruta, S.; Lourenco, D.; Masuda, Y.; Aguilar, I.; Legarra, A.; Vitezica, Z. Manual for BLUPF90 Family Programs. University of Georgia. 2018. Available online: http://nce.ads.uga.edu/wiki/doku.php?id=documentation (accessed on 8 December 2022).
- Sewalem, A.; Kistemaker, G.J.; Ducrocq, V.; Van Doormaal, B.J. Genetic analysis of herd life in Canadian dairy cattle on a lactation basis using a Weibull proportional hazards model. J. Dairy Sci. 2005, 88, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, O.; Aihara, M.; Nishiura, A.; Takeda, H.; Satoh, M. Genetic Analysis of the Cumulative Pseudo-Survival Rate During Lactation of Holstein Cattle in Japan by Using Random Regression Models. J. Dairy Sci. 2015, 98, 5781–5795. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Method of Weighting * | Without Weight | w1 | w2 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Individual | Failure to Survive a Stage | BV | r2 | Weight | BV | r2 | Weight | BV | r2 |
| A | 1 | −0.73 | 0.02 | 9 | −6.96 | 0.11 | 16 | −10.09 | 0.15 |
| B | 2 | −0.60 | 0.04 | 8 | −5.42 | 0.12 | 13 | −7.63 | 0.15 |
| C | 3 | −0.47 | 0.05 | 7 | −3.89 | 0.12 | 10 | −4.95 | 0.15 |
| D | 4 | −0.34 | 0.07 | 6 | −2.37 | 0.13 | 8 | −2.76 | 0.15 |
| E | 5 | −0.21 | 0.09 | 5 | −0.85 | 0.14 | 6 | −0.42 | 0.15 |
| F | 6 | −0.09 | 0.10 | 4 | 0.69 | 0.14 | 5 | 1.07 | 0.15 |
| G | 7 | 0.03 | 0.12 | 3 | 2.25 | 0.14 | 3 | 3.75 | 0.15 |
| H | 8 | 0.15 | 0.13 | 2 | 3.84 | 0.15 | 2 | 5.34 | 0.15 |
| I | 9 | 0.26 | 0.15 | 1 | 5.49 | 0.15 | 1 | 6.99 | 0.15 |
| J | Survival 9 | 1.99 | 0.15 | 1 | 7.22 | 0.15 | 1 | 8.71 | 0.15 |
| Statistical Model | ||||
|---|---|---|---|---|
| without Weight | with Weight (w2) | |||
| Cow with records | 298,290 | |||
| Individuals in the pedigree | 660,476 | |||
| HYS, records | Number 15,919, Mean size 113.93, Range 15 to 650 | |||
| Milk deviation in the herd (SD) | Average 0.99 (0.21), Range 0.06 to 4.60 | |||
| Correlation survival rate x milk | 0.13 | 0.22 | ||
| Period | Frequency % | Discarding % (SD) | Weighted Frequency % | Weighted Discarding % (SD) |
| Average | 12.29 (32.8) | 49.01 (50.0) | ||
| 1 | 16.45 | 3.64 (18.7) | 14.78 | 37.68 (48.5) |
| 2 | 15.85 | 8.69 (28.2) | 18.83 | 55.31 (49.7) |
| 3 | 14.47 | 18.67 (39.0) | 22.55 | 69.66 (46.0) |
| 4 | 11.77 | 3.43 (18.2) | 8.48 | 22.11 (41.5) |
| 5 | 11.36 | 12.74 (33.3) | 10.82 | 46.71 (49.9) |
| 6 | 9.92 | 25.61 (43.6) | 11.67 | 63.26 (48.2) |
| 7 | 7.38 | 5.35 (22.5) | 4.75 | 14.49 (35.2) |
| 8 | 6.98 | 16.56 (37.2) | 4.73 | 28.41 (45.1) |
| 9 | 5.83 | 28.60 (45.2) | 3.39 | 28.60 (45.2) |
| Number of survival records | 1,813,636 | |||
| Average weight (w2) | 1.72 | |||
| Sum of weights (w2) | 3,119,557 | |||
| Effects Considered | Without Weight | With Weight (w2) | ||
|---|---|---|---|---|
| SD | R2 | SD | R2 | |
| Simple records | 32.81 | 49.99 | ||
| Effects of the herd–year–season least-squares method | 32.14 | 0.05 | 48.13 | 0.08 |
| Herd–year–season + period | 31.26 | 0.10 | 45.26 | 0.18 |
| Herd–year–season + period + milk | 30.99 | 0.12 | 43.88 | 0.23 |
| Herd–year–season + period + milk + sire | 30.94 | 0.12 | 43.72 | 0.24 |
| Quantity | Statistical Model | |||
|---|---|---|---|---|
| SURV | SURVm | SURVSw2 | SURVmw2 | |
| s2r | 965.97 (1.02) | 947.02 (1.00) | 1545.40 (1.35) | 1529.40 (1.34) |
| s2G | 12.46 (0.02) | 14.60 (0.03) | 181.26 (7.56) | 166.81 (6.94) |
| sPE | 0.00 | 0.00 | 1397.70 (6.76) | 1269.60 (6.22) |
| s2P | 978.43 (1.02) | 961.62 (1.00) | 3124.36 (5.19) | 2965.81 (4.78) |
| h2 | 0.013 (0.0000) | 0.015 (0.0000) | 0.058 (0.0024) | 0.056 (0.0023) |
| r | 0.013 (0.0000) | 0.015 (0.0000) | 0.505 (0.0009) | 0.484 (0.0009) |
| k = s2r/s2G | 77.52 (0.1558) | 64.54 (0.1302) | 8.536 (0.3598) | 9.169 (0.3856) |
| Effects | Statistical Model | ||||
|---|---|---|---|---|---|
| SURV | SURVm | SURVw2 | SURVmw2 | ||
| Herd–year–season | SD | 10.73 | 10.59 | 18.37 | 17.94 |
| Min | −88.08 | −88.92 | −105.62 | −100.20 | |
| Max | 28.02 | 27.07 | 57.40 | 51.34 | |
| Periods | SD | 8.81 | 8.71 | 19.98 | 19.65 |
| Min | −13.25 | −13.17 | −30.48 | −30.10 | |
| Max | 9.84 | 9.68 | 34.69 | 33.95 | |
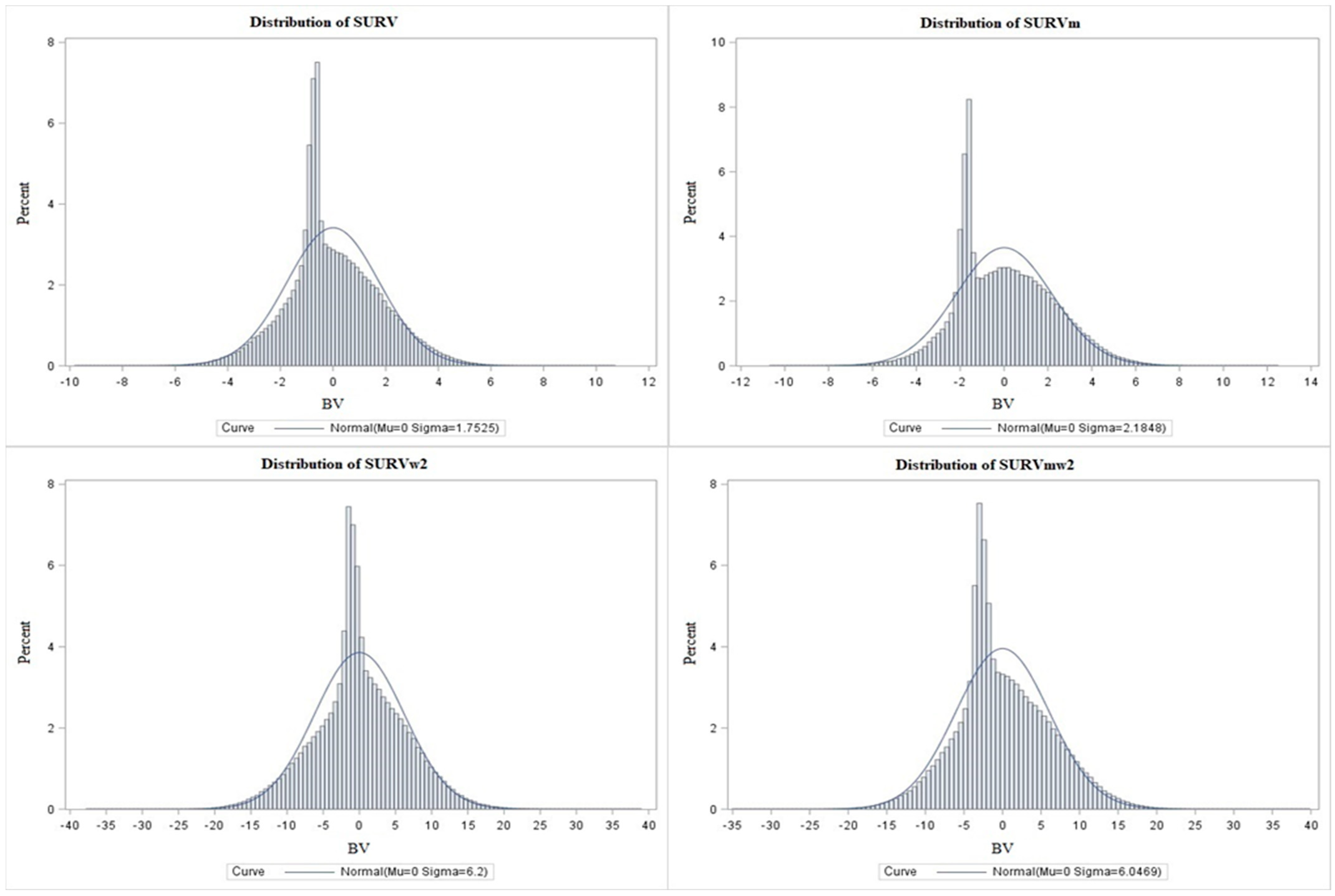
| Breeding values | SD | 1.75 | 2.18 | 6.20 | 6.04 |
| Min | −10.64 | −12.43 | −38.79 | −39.64 | |
| Max | 9.76 | 10.63 | 37.46 | 34.95 | |
| Permanent environment | SD | - | - | 33.93 | 32.19 |
| Min | - | - | −107.96 | −137.79 | |
| Max | - | - | 85.18 | 86.68 | |
| Milk regression | - | 20.13 | - | 31.22 | |
| Period | Statistical Model | |||
|---|---|---|---|---|
| SURV | SURVm | SURVw2 | SURVmw2 | |
| 1 | 9.43 | 9.52 | 34.69 | 33.95 |
| 2 | 4.48 | 4.61 | 19.28 | 18.86 |
| 3 | −5.15 | −4.92 | −6.29 | −6.73 |
| 4 | 9.84 | 9.68 | 14.80 | 14.85 |
| 5 | 0.68 | 0.63 | −7.71 | −7.49 |
| 6 | −11.69 | −11.45 | −30.48 | −30.10 |
| 7 | 8.31 | 7.97 | 2.63 | 2.89 |
| 8 | −2.64 | −2.88 | −12.33 | −12.16 |
| 9 | −13.25 | −13.17 | −14.58 | −14.06 |
| Herd–Year–Season | Breeding Values | Permanent Environment | |||||
|---|---|---|---|---|---|---|---|
| SURVm | SURVw2 | SURVmw2 | SURVm | SURVw2 | SURVmw2 | SURVmw2 | |
| SURV | 100 | 88 | 89 | 91 | 92 | 94 | |
| SURVm | 88 | 89 | 75 | 89 | |||
| SURVw2 | 99 | 96 | 99 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Černá, M.; Zavadilová, L.; Vostrý, L.; Bauer, J.; Šplíchal, J.; Vařeka, J.; Fulínová, D.; Brzáková, M. Genetic Parameters for a Weighted Analysis of Survivability in Dairy Cattle. Animals 2023, 13, 1188. https://doi.org/10.3390/ani13071188
Černá M, Zavadilová L, Vostrý L, Bauer J, Šplíchal J, Vařeka J, Fulínová D, Brzáková M. Genetic Parameters for a Weighted Analysis of Survivability in Dairy Cattle. Animals. 2023; 13(7):1188. https://doi.org/10.3390/ani13071188
Chicago/Turabian StyleČerná, Michaela, Ludmila Zavadilová, Luboš Vostrý, Jiří Bauer, Jiří Šplíchal, Jan Vařeka, Daniela Fulínová, and Michaela Brzáková. 2023. "Genetic Parameters for a Weighted Analysis of Survivability in Dairy Cattle" Animals 13, no. 7: 1188. https://doi.org/10.3390/ani13071188