
Morphological and Molecular Investigations of Aquaporin-7 (AQP-7) in Male Camelus dromedarius Reproductive Organs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pre-Experimental Ethics and Experimental Sampling
2.2. Immunohistochemistry Paraffin Protocol (IHC-P) for AQP-7 Detection
2.3. RNA Isolation and Semi-Quantitative Real Time RT-PCR Analysis
2.4. Statistical Analysis
3. Results
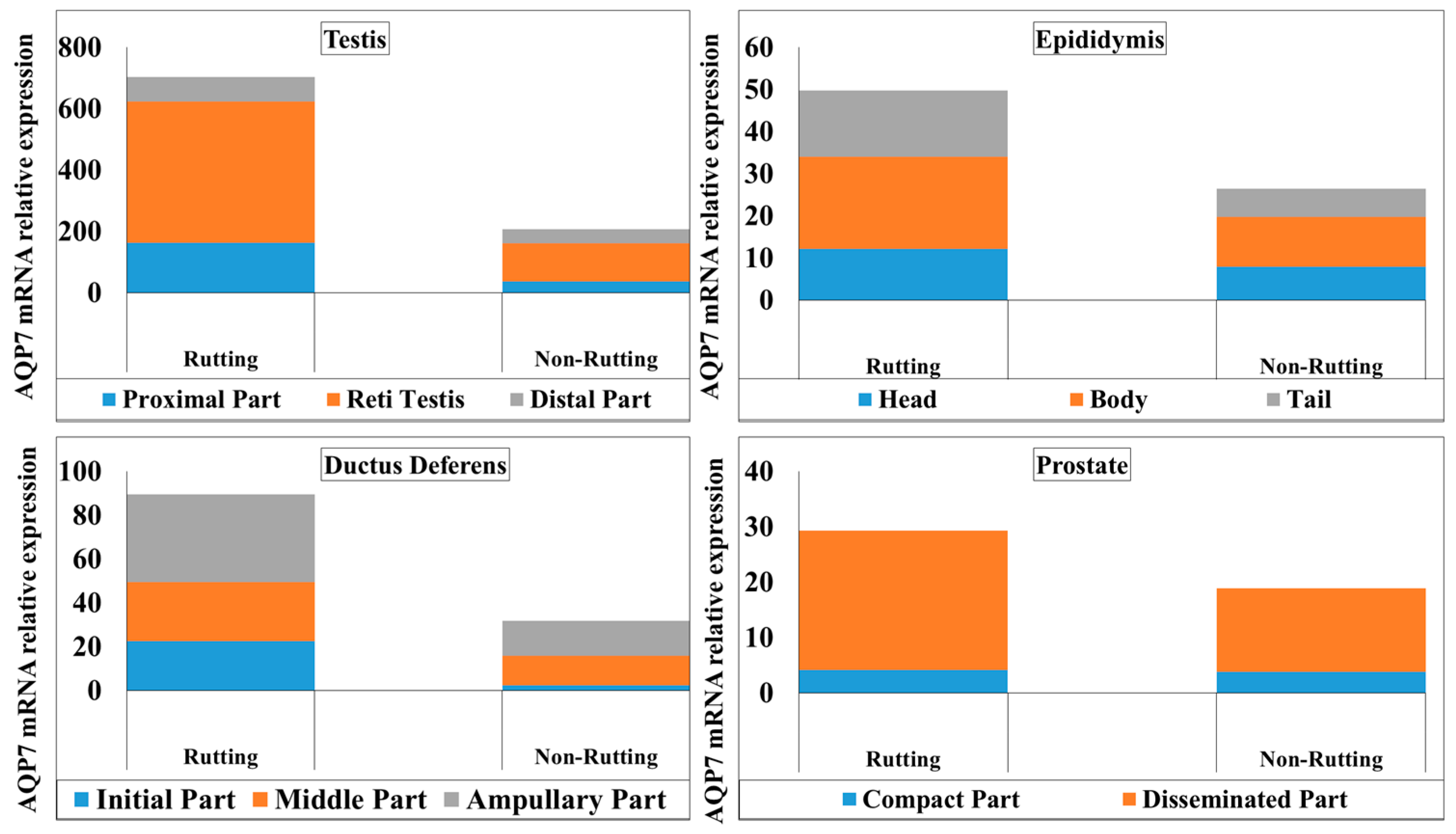
3.1. AQP-7 Gene Expression
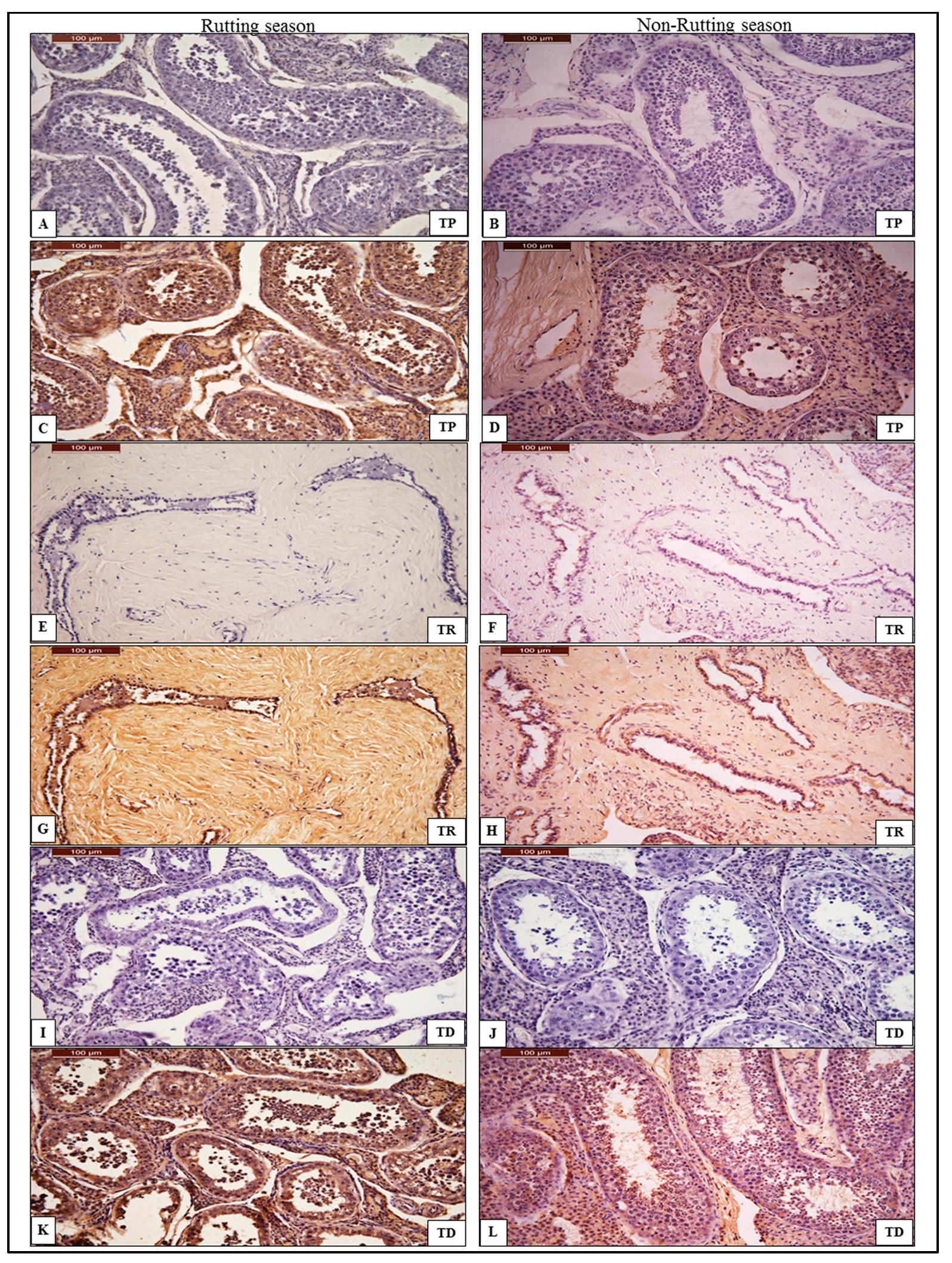
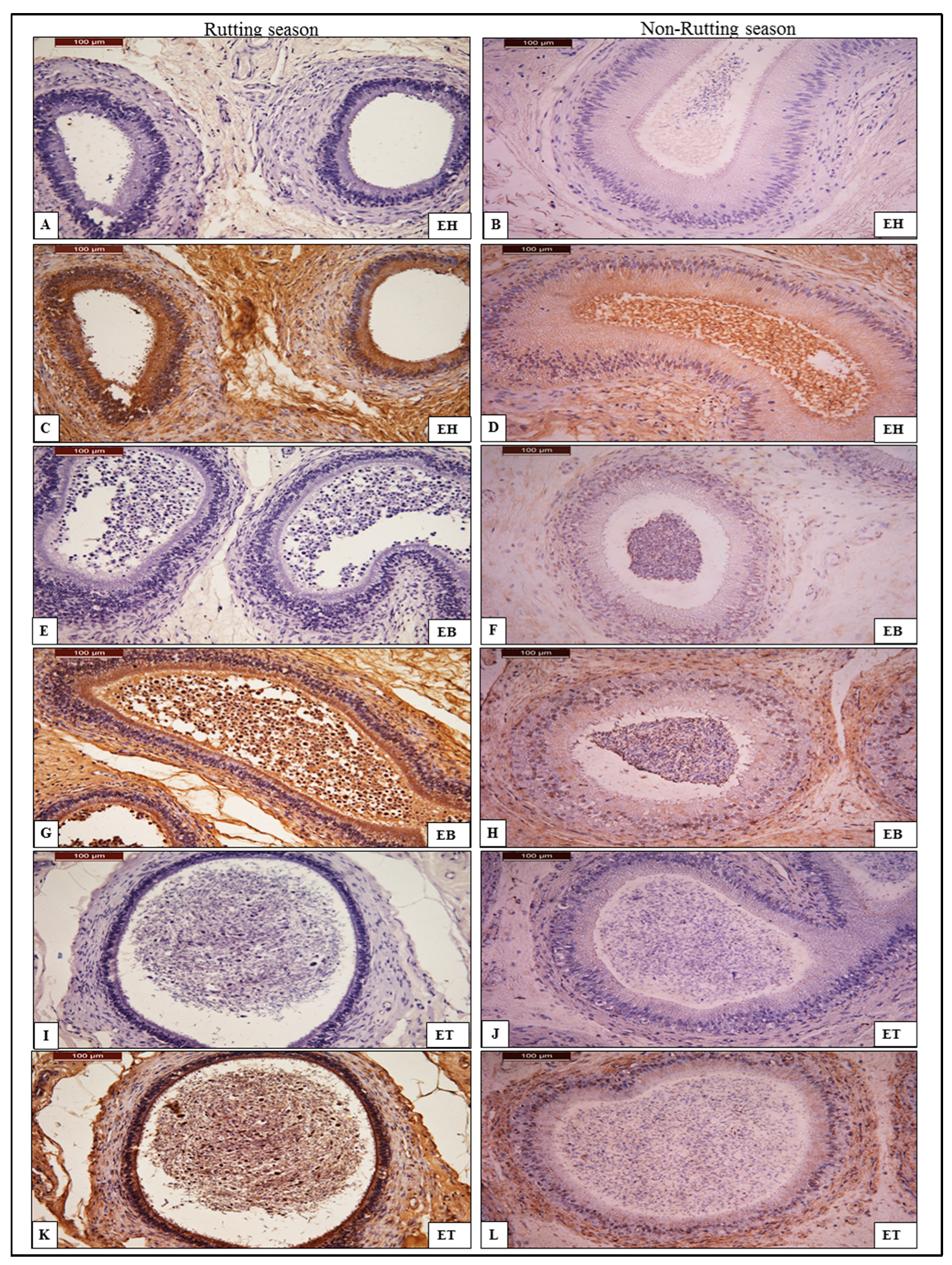
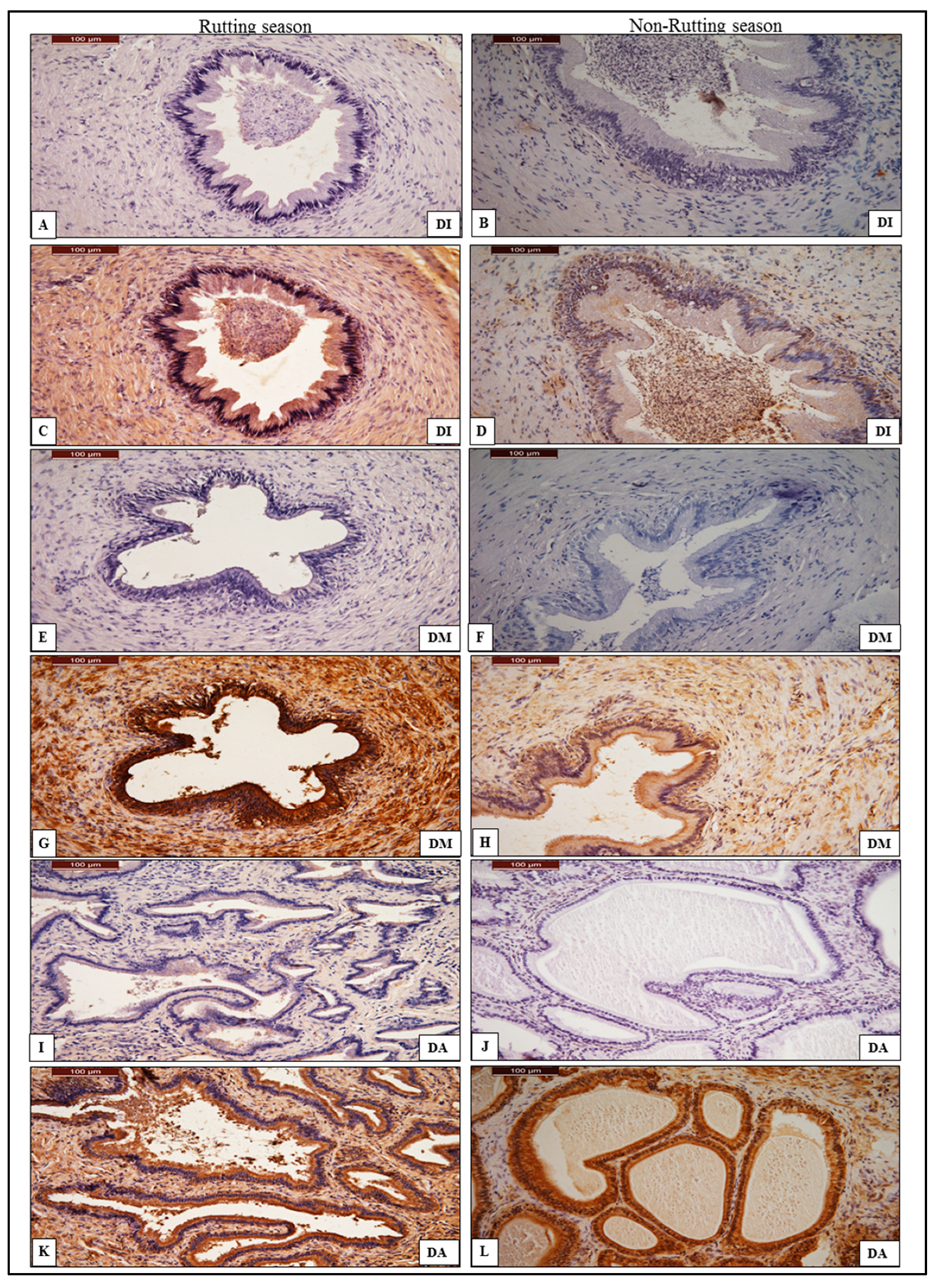
3.2. Immunohistochemistry of AQP-7
3.2.1. Testes
3.2.2. Epididymis
3.2.3. Ductus Deferens
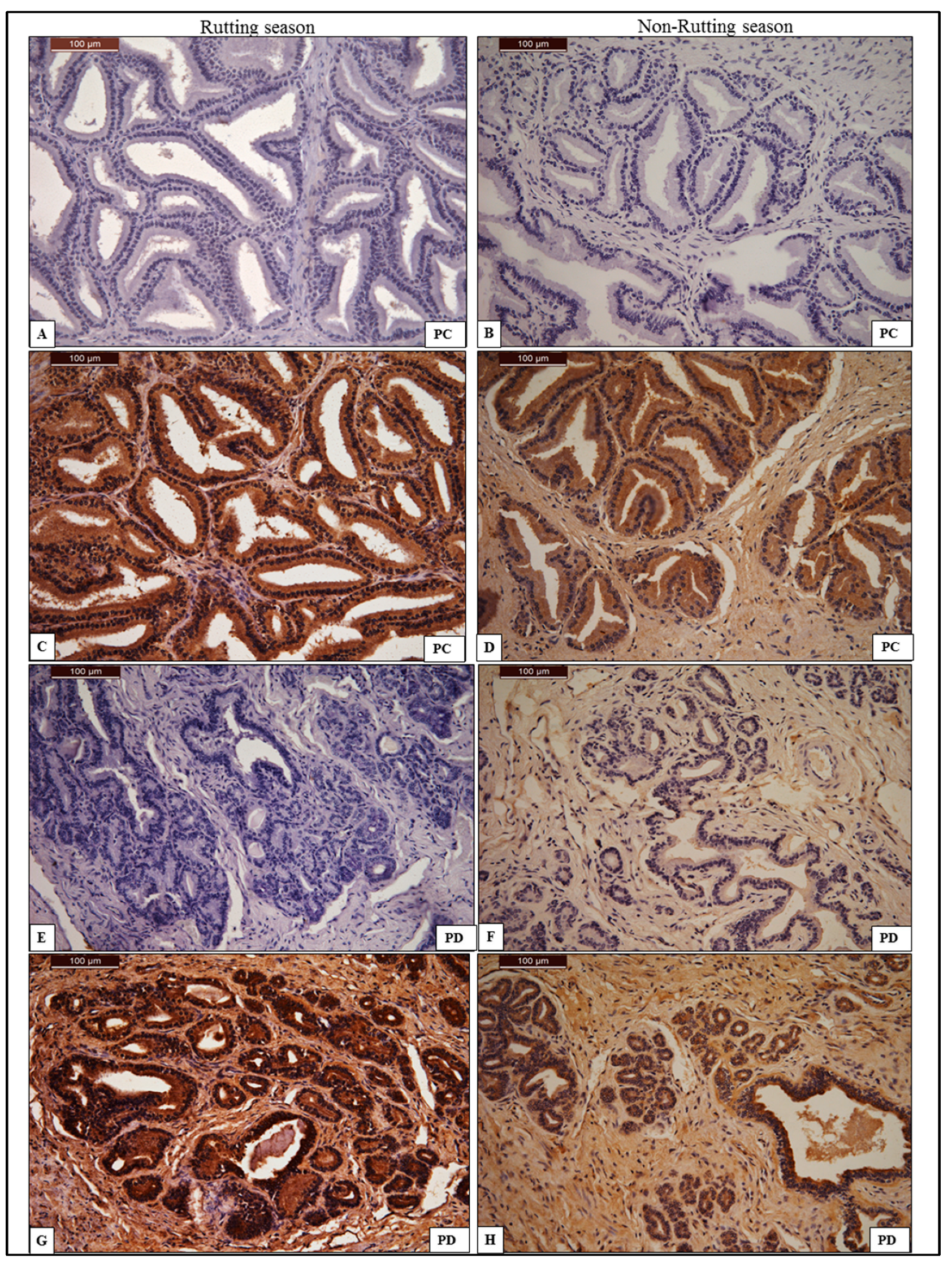
3.2.4. Prostate Gland
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ishibashi, K.; Hara, S.; Kondo, S. Aquaporin water channels in mammals. Clin. Exp. Nephrol. 2009, 13, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Kreida, S.; Toernroth-Horsefield, S. Structural insights into aquaporin selectivity and regulation. Urr. Opin. Struct. Biol. 2015, 33, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Gomes, D.; Agasse, A.; Thiebaud, P.; Delrot, S.; Geros, H.; Chaumont, F. Aquaporins are multifunctional water and solute transporters highly divergent in living organisms. Biochim. Biophys. Acta 2009, 1788, 1213–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domeniconi, R.F.; Orsi, A.M.; Justulin, L.A.; Beu, C.C., Jr.; Felisbino, S.L. Aquaporin 9 (AQP9) localization in the adult dog testis excurrent ducts by immunohistochemistry. Anat. Rec. 2007, 290, 1519–1525. [Google Scholar] [CrossRef] [Green Version]
- Arrighi, S.; Aralla, M. Immunolocalization of aquaporin water channels in the domestic cat male genital tract. Reprod. Domest. Anim. 2014, 49, 17–26. [Google Scholar] [CrossRef]
- Ishibashi, K.; Tanaka, Y.; Morishita, Y. The role of mammalian superaquaporins inside the cell. Biochim. Biophys. Acta 2014, 1840, 1507–1512. [Google Scholar] [CrossRef]
- Mosca, A.F.; Almeida, A.; de Wragg, D.; Martins, A.P.; Sabir, F.; Leoni, S.; Moura, T.F.; Prista, C.; Casini, A.; Soveral, G. Molecular basis of aquaporin-7 permeability regulation by pH. Cells. 2018, 7, 207. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Kageyama, Y.; Okada, Y.; Kawakami, S.; Kihara, K.; Ishibashi, K.; Sasaki, S. Localization of aquaporin-7 in human testis and ejaculated sperm: Possible involvement in maintenance of sperm quality. J. Urol. 2004, 172, 2073–2076. [Google Scholar] [CrossRef]
- Arrighi, S.; Romanello, M.G.; Domeneghini, C. Ultrastructure of epididymal epithelium in Equus caballus. Ann. Anat. Anatomischer Anzeiger 1993, 175, 1–9. [Google Scholar] [CrossRef]
- Klein, C.; Troedsson, M.H.; Rutllant, J. Region-specific expression of aquaporin subtypes in equine testis, epididymis, and ductus deferens. Anat. Rec. 2013, 296, 1115–1126. [Google Scholar] [CrossRef] [Green Version]
- Calamita, G.; Mazzone, A.; Bizzoca, A.; Svelto, M. Possible involvement of aquaporin 7 and 8 in rat testis development and spermatogenesis. Biochem. Biophys. Res. Commun. 2001, 288, 619–625. [Google Scholar] [CrossRef] [PubMed]
- Squillacioti, C.; Mirabella, N.; Liguori, G.; Germano, G.; Pelagalli, A. Aquaporins are differentially regulated in canine cryptorchid efferent ductules and epididymis. Animals 2021, 11, 1539. [Google Scholar] [CrossRef] [PubMed]
- Marai, I.F.M.; Zeidan, A.E.B.; Abdel-Samee, A.M.; Abizaid, A.; Fadiel, A. Camels’ reproductive and physiological performance traits as affected by environmental conditions. Trop. Subtrop. Agroecosystems 2009, 10, 129–149. [Google Scholar]
- Merkt, H.; Rath, O.; Musa, B.; El-Naggar, M.A. Reproduction in Camels; FAO Animal Production and Health: Rome, Italy, 1990; Volume 82. [Google Scholar]
- Musa, B.; Merkt, H.; Sieme, H.; Hago, B.; Hoppen, H. The female camel (Camelus dromedarius) and the artificial insemination. In Proceedings of the UCDEC Workshop, Paris, France, 8–13 July 1990; pp. 257–261. [Google Scholar]
- Arrighi, S.; Bosi, G.; Accogli, G.; Desantis, S. Seasonal and ageing-depending changes of aquaporins 1 and 9 expression in the genital tract of buffalo bulls (Bubalus bubalis). Reprod. Domest. Anim. 2016, 51, 515–523. [Google Scholar] [CrossRef]
- Al-Eknah, M.M.; Homeida, A.M.; Al-Bisher, B.E. Physicochemical properties of the cervical mucus of the pregnant camel (Camelus dromedarius). Pak. Vet. 1997, 17, 91–93. [Google Scholar]
- Al Eknah, M.M. Reproduction in old world camels. Anim. Reprod. Sci. 2000, 60–61, 583–592. [Google Scholar] [CrossRef]
- El-Wishy, A.B. Reproduction in the male dromedary (Camelus dromedarius): A Review. Anim. Reprod. Sci. 1988, 17, 217–241. [Google Scholar] [CrossRef]
- Verkman, A.S. Water permeability measurement in living cells and complex tissues. J. Membr. Biol. 2000, 173, 73–87. [Google Scholar] [CrossRef]
- Madeira, A.; Moura, T.F.; Soveral, G. Detecting aquaporin function and regulation. Front. Chem. 2016, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Pastor-Soler, N.; Bagnis, C.; Sabolic, I.; Tyszkowski, R.; McKee, M.; Van Hoek, A.; Breton, S.; Brown, D. Aquaporin 9 expression along the male reproductive tract. Biol. Reprod. 2001, 65, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Levine, N.; Marsh, D.J. Micropuncture studies of the electrochemical aspects of fluid and electrolyte transport in individual seminiferous tubules, the epididymis and the vas deferens in rats. J. Physiol. 1971, 213, 557–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R. Plasma membrane structure and remodelling during sperm maturation in the epididymis. J. Reprod. Fertil. Suppl. 1998, 53, 73–84. [Google Scholar] [PubMed]
- Da Silva, N.; Piétrement, C.; Brown, D.; Breton, S. Segmental and cellular expression of aquaporins in the male excurrent duct. Biochim. Biophys. Acta. 2006, 1758, 1025–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domeniconi, R.F.; Orsi, A.M.; Justulin, L.A.; Leme Beu, C.C., Jr.; Felisbino, S.L. Immunolocalization of aquaporins 1, 2 and 7 in rete testis, efferent ducts, epididymis and vas deferens of adult dog. Cell Tissue Res. 2008, 332, 329–335. [Google Scholar] [CrossRef]
- An, F.; Wang, J. Immunohistochemical study of aquaporin 1 and 2 in the reproductive tract of the bactrian camel (Camelus bactrianus). Int. J. Morphol. 2016, 34, 1218–1222. [Google Scholar] [CrossRef] [Green Version]
- Badran, H.H.; Hermo, L.S. Expression and regulation of aquaporins 1, 8, and 9 in the testis, efferent ducts, and epididymis of adult rats and during postnatal development. J. Androl. 2002, 23, 358–373. [Google Scholar]
- Hara-Chikuma, M.; Verkman, A.S. Physiological roles of glycerol-transporting aquaporins: The aquaglyceroporins. Cell Mol. Life Sci. 2006, 63, 1386–1392. [Google Scholar] [CrossRef]
- Rodriguez, A.; Catalan, V.; Gomez-Ambrosi, J.; Fruhbeck, G. Aquaglyceroporins serve as metabolic gateways in adiposity and insulin resistance control. Cell Cycle 2011, 10, 1548–1556. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Location of Tissue Sample | ||
|---|---|---|---|
| Testis | Proximal part (TP) | Distal part (TD) | Rete testis (TR) |
| Epididymis | Head (EH) | Body (EB) | Tail (ET) |
| Ductus deferens | Initial part (DI) | Middle part (DM) | Ampullary part (DA) |
| Prostate | Compact part (PC) | Disseminated part (PD) | |
| Primers | Sequence | GenBank Reference |
|---|---|---|
| AQP-7 | F-5-CAGAGAGGAAGAGGCGGTCT-3 R-5-CGTGAACTCGGCTAGGAACT-3 | XM_010975828.2 |
| GAPDH | F-5-TCGATCCCCCAACACACTTG-3 | XM_010990867.2 |
| R-5-TGATGGTGCATGACAAGGCA-3 |
| Immunoreactivity of Rabbit Polyclonal Anti-AQP-7 Antibody | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Tissue Sample | Testis | Epididymis | Ductus Deferens | Prostate Gland | |||||||
| TP | TR | TD | EH | EB | ET | DI | DM | DA | PC | PD | |
| Rutting | + + + + | + + + + | + + + + | + + + + | + + + + | + + + + | + + + + | + + + + | + + + + | + + + + | + + + + |
| Non-Rutting | + | + + + + | + + + + | + | + | + + | + + | + + | + + + | + + + + | + + + + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Thnaian, T.A. Morphological and Molecular Investigations of Aquaporin-7 (AQP-7) in Male Camelus dromedarius Reproductive Organs. Animals 2023, 13, 1158. https://doi.org/10.3390/ani13071158
Al-Thnaian TA. Morphological and Molecular Investigations of Aquaporin-7 (AQP-7) in Male Camelus dromedarius Reproductive Organs. Animals. 2023; 13(7):1158. https://doi.org/10.3390/ani13071158
Chicago/Turabian StyleAl-Thnaian, Thnaian A. 2023. "Morphological and Molecular Investigations of Aquaporin-7 (AQP-7) in Male Camelus dromedarius Reproductive Organs" Animals 13, no. 7: 1158. https://doi.org/10.3390/ani13071158