
Dietary Frankincense (Boswellia serrata) Oil Modulates the Growth, Intestinal Morphology, the Fatty Acid Composition of Breast Muscle, Immune Status, and Immunoexpression of CD3 and CD20 in Broiler Chickens
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
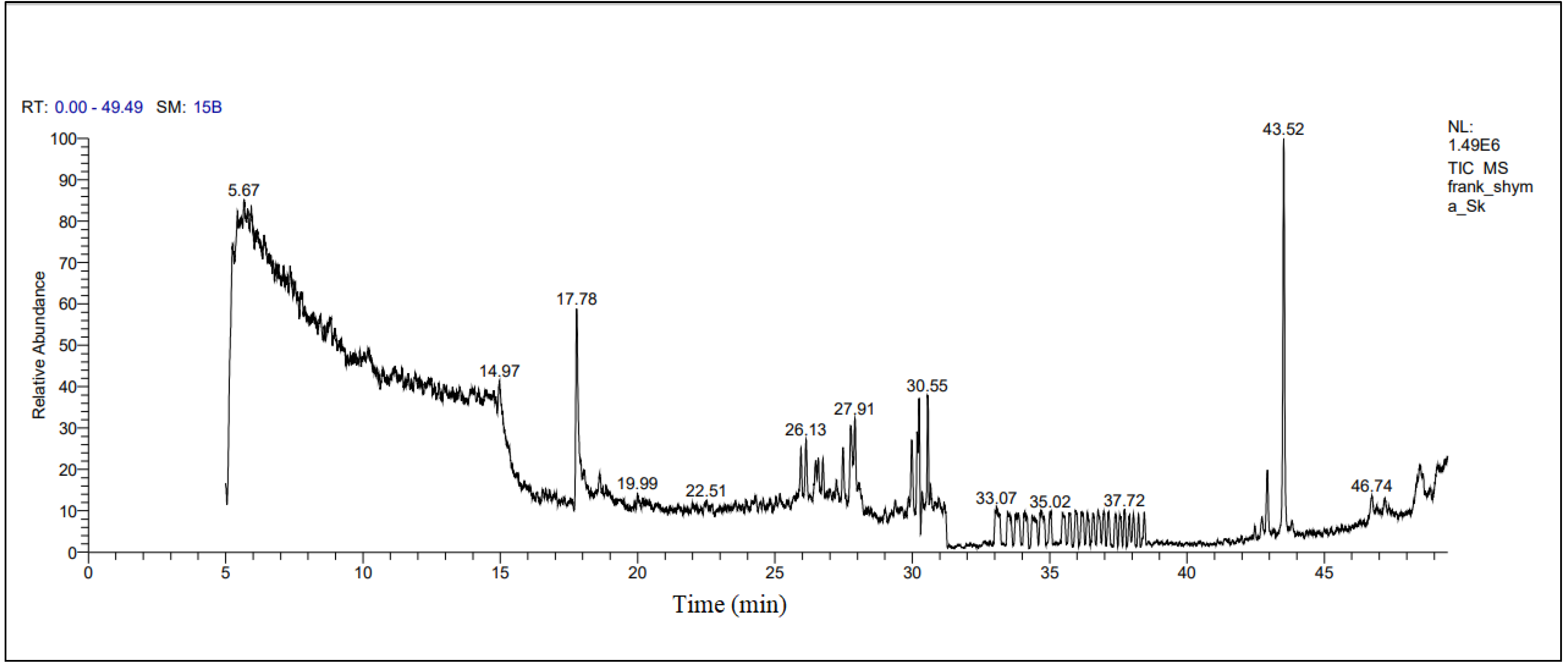
2.1. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis of FO
2.2. Birds
2.3. Experimental Design and Diets
2.4. Growth Performance
2.5. Percentage Calculations of the Dressing, Internal Organs, and Immune Organs
2.6. Sampling
2.7. Fatty Acid Analysis of the Breast Muscle
2.8. Intestinal Histology and Morphometric Measures
2.9. Blood Biochemical Parameters
2.10. Immunological Parameters
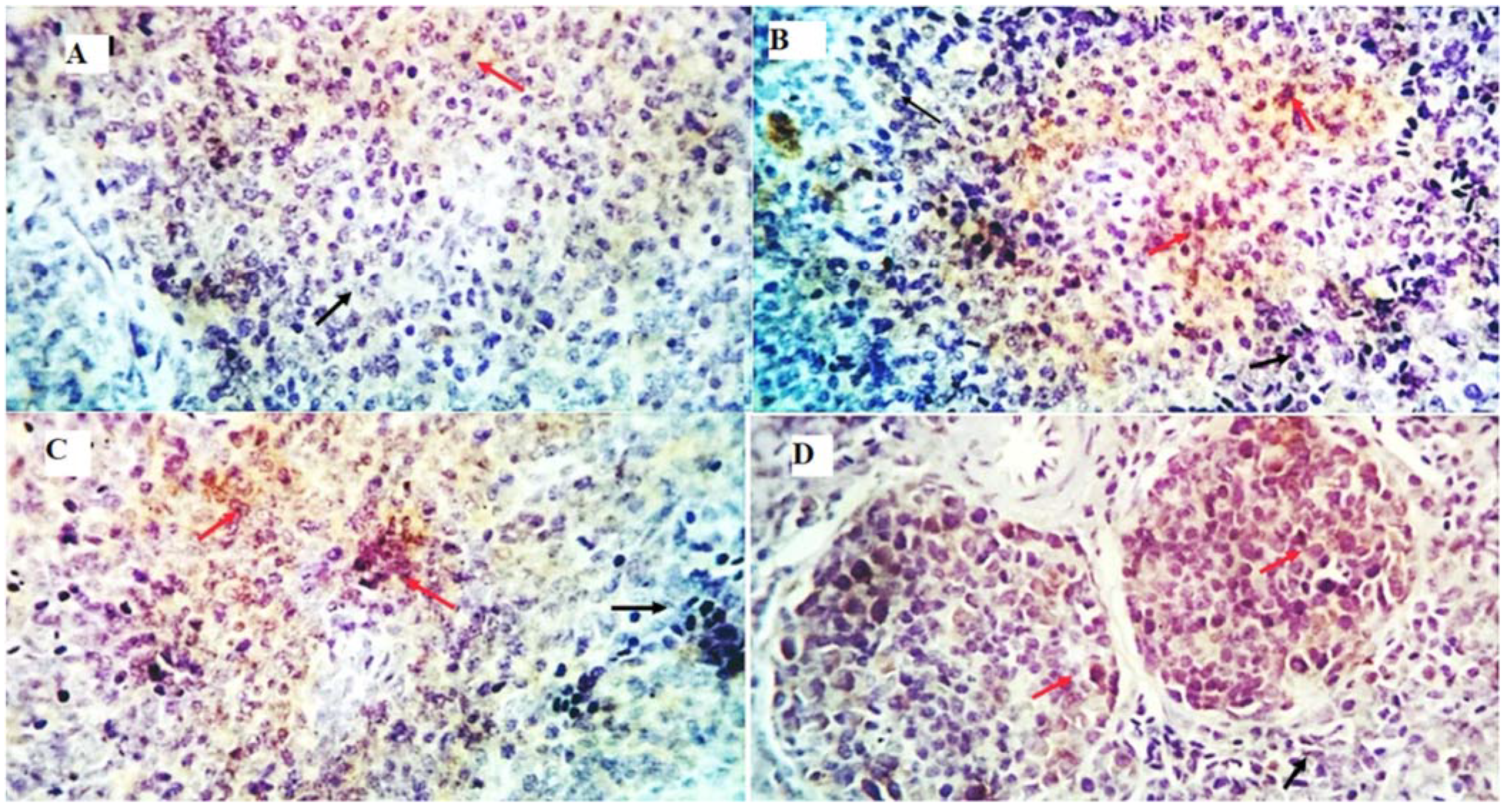
2.11. Immunohistochemical Examination
2.12. Statistical Analysis
3. Results
3.1. Determination of Bioactive Compounds in FO
3.2. Growth Performance
3.3. Percentages of the Dressing, Internal Organs, and Immune Organs
3.4. Fatty Acid Composition of Breast Muscle
3.5. Histological Examination
3.6. Serum Biochemical Parameters
3.7. Immunological Parameters
3.8. Immunohistochemical Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abd El-Hack, M.E.; Alagawany, M. Performance, egg quality, blood profile, immune function, and antioxidant enzyme activities in laying hens fed diets with thyme powder. J. Anim. Feed Sci. 2015, 24, 127–133. [Google Scholar] [CrossRef]
- Belitz, H.-D.; Grosch, W.; Schieberle, P. Meat. In Food Chemistry; Springer: Berlin/Heidelberg, Germany, 2009; pp. 563–616. [Google Scholar]
- European Commission. European Union Register of Feed Additives Pursuant to Regulation (EC) No 1831/2003; European Commission: Brussels, Belgium, 2014. [Google Scholar]
- Afsharypuor, S.; Rahmany, M. Essential oil constituents of two African Olibanums available in Isfahan Commercial Market. Iran. J. Pharm. Sci. 2005, 1, 167–170. [Google Scholar]
- Al-Yasiry, A.R.M.; Kiczorowska, B. Frankincense-therapeutic properties. Adv. Hyg. Exp. Med. Postep. Hig. I Med. Dosw. 2016, 70, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Yates, R.L.; Wenninger, J.A. Constituents of olibanum oil: Sesquiterpene hydrocarbons. J. Assoc. Off. Anal. Chem. 1970, 53, 941–948. [Google Scholar] [CrossRef]
- Al-Yasiry, A.; Kiczorowska, B.; Samolińska, W. Effect of Boswellia serrata resin supplementation on basic chemical and mineral element composition in the muscles and liver of broiler chickens. Biol. Trace Elem. Res. 2017, 179, 294–303. [Google Scholar] [CrossRef]
- Yasrib, Q.; Abid, H.; Shashank, K.S.; Ajit, K.S. Potential role of natural molecules in health and disease: Importance of boswellic acid. J. Med. Plants Res. 2010, 4, 2778–2786. [Google Scholar]
- Hamidpour, S.; Hamidpour, M.; Shahlari, M.; Hamidpour, R. Chemistry, pharmacology and medicinal property of frankincense (Boswellia species): From the selection of traditional applications to the novel phytotherapy for the prevention and treatment of serious diseases. Glob. J. Med. Res. 2015, 15, 1–9. [Google Scholar]
- Olukemi, M.A.; Kandakai-Olukemi, Y.T.; Mawak, J.D. Antibacterial activity of the stem bark of Boswellia dalzielii. J. Pharm. Bioresour. 2005, 2, 32076. [Google Scholar] [CrossRef]
- Kiczorowska, B.; Al-Yasiry, A.; Samolińska, W.; Marek, A.; Pyzik, E. The effect of dietary supplementation of the broiler chicken diet with Boswellia serrata resin on growth performance, digestibility, and gastrointestinal characteristics, morphology, and microbiota. Livest. Sci. 2016, 191, 117–124. [Google Scholar] [CrossRef]
- Mikhaeil, B.R.; Maatooq, G.T.; Badria, F.A.; Amer, M.M. Chemistry and immunomodulatory activity of frankincense oil. Z. Nat. C 2003, 58, 230–238. [Google Scholar] [CrossRef]
- Ayub, M.A.; Hanif, M.A.; Sarfraz, R.A.; Shahid, M. Biological activity of Boswellia serrata Roxb. oleo gum resin essential oil: Effects of extraction by supercritical carbon dioxide and traditional methods. Int. J. Food Prop. 2018, 21, 808–820. [Google Scholar] [CrossRef]
- Sadhasivam, S.; Palanivel, S.; Ghosh, S. Synergistic antimicrobial activity of Boswellia serrata Roxb. ex Colebr. (Burseraceae) essential oil with various azoles against pathogens associated with skin, scalp and nail infections. Lett. Appl. Microbiol. 2016, 63, 495–501. [Google Scholar] [CrossRef]
- Montaser, M.M.; El-Sharnouby, M.E.; El-Noubi, G.; El-Shaer, H.M.; Khalil, A.A.; Hassanin, M.; Amer, S.A.; El-Araby, D.A. Boswellia serrata resin extract in diets of Nile tilapia, Oreochromis niloticus: Effects on the growth, health, immune response, and disease resistance to Staphylococcus aureus. Animals 2021, 11, 446. [Google Scholar] [CrossRef]
- Camarda, L.; Dayton, T.; Di Stefano, V.; Pitonzo, R.; Schillaci, D. Chemical composition and antimicrobial activity of some oleogum resin essential oils from Boswellia spp. (Burseraceae). Ann. Di Chim. J. Anal. Environ. Cult. Herit. Chem. 2007, 97, 837–844. [Google Scholar]
- Gangwal, M.; Vardhan, D. Antifungal studies of volatile constituents of Boswellia serrata. Asian J. Chem. 1995, 7, 675. [Google Scholar]
- Etuk, E.; Agaie, B.; Onyeyili, P.; Ottah, C. Toxicological studies of aqueous stem bark extract of Boswellia dalzielii in albino rats. Indian J. Pharmacol. 2006, 38, 359. [Google Scholar] [CrossRef]
- Ameen, A.M.; Elkazaz, A.Y.; Mohammad, H.M.; Barakat, B.M. Anti-inflammatory and neuroprotective activity of boswellic acids in rotenone parkinsonian rats. Can. J. Physiol. Pharmacol. 2017, 95, 819–829. [Google Scholar] [CrossRef]
- Rajabian, A.; Sadeghnia, H.; Fanoudi, S.; Hosseini, A. Genus Boswellia as a new candidate for neurodegenerative disorders. Iran. J. Basic Med. Sci. 2020, 23, 277. [Google Scholar]
- Lawless, J. The Illustrated Encyclopedia of Essential Oils Element Books; Element Books Ltd.: Rockport, MA, USA, 1995. [Google Scholar]
- Amer, S.A.; Abdel-Wareth, A.A.; Gouda, A.; Saleh, G.K.; Nassar, A.H.; Sherief, W.R.; Albogami, S.; Shalaby, S.I.; Abdelazim, A.M.; Abomughaid, M.M. Impact of Dietary Lavender Essential Oil on the Growth and Fatty Acid Profile of Breast Muscles, Antioxidant Activity, and Inflammatory Responses in Broiler Chickens. Antioxidants 2022, 11, 1798. [Google Scholar] [CrossRef]
- Ross, A. 708 Broiler Nutrition Specification 2014. Available online: http://en.aviagen.com/ross-708/ (accessed on 6 October 2014).
- Association, A.V.M. AVMA Guidelines for the Euthanasia of Animals: 2013 Edition; American Veterinary Medical Association: Schaumburg, IL, USA, 2013. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International; AOAC: Rockville, MD, USA, 2000. [Google Scholar]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Amer, S.A.; Al-Khalaifah, H.S.; Gouda, A.; Osman, A.; Goda, N.I.; Mohammed, H.A.; Darwish, M.I.; Hassan, A.M.; Mohamed, S.K.A. Potential effects of anthocyanin-rich roselle (Hibiscus sabdariffa L.) extract on the growth, intestinal histomorphology, blood biochemical parameters, and the immune status of broiler chickens. Antioxidants 2022, 11, 544. [Google Scholar] [CrossRef]
- Grant, G.H. Amino acids and proteins. In Fundamentals of Clinical Chemistry; Academic Press: Cambridge, MA, USA, 1987. [Google Scholar]
- Doumas, B.; Baysa, D.; Carter, R.; Peters, T.; Schaffer, R. Determination of serum total protein. Clin. Chem. 1981, 27, 1642. [Google Scholar] [CrossRef] [PubMed]
- Doumas, B.T.; BIGGS, H.G.; ARENDS, R.L.; PINTO, P.V. Determination of serum albumin. In Standard Methods of Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 1972; Volume 7, pp. 175–188. [Google Scholar]
- Trinder, P. Determination of blood glucose using an oxidase-peroxidase system with a non-carcinogenic chromogen. J. Clin. Pathol. 1969, 22, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Lie, Ø.; Syed, M.; Solbu, H. Improved agar plate assays of bovine lysozyme and haemolytic complement activity. Acta Vet. Scand. 1986, 27, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Saber, S.; Khalil, R.M.; Abdo, W.S.; Nassif, D.; El-Ahwany, E. Olmesartan ameliorates chemically-induced ulcerative colitis in rats via modulating NFκB and Nrf-2/HO-1 signaling crosstalk. Toxicol. Appl. Pharmacol. 2019, 364, 120–132. [Google Scholar] [CrossRef]
- Hashish, H.; Kamal, R. Effect of curcumin on the expression of Caspase-3 and Bcl-2 in the spleen of diabetic rats. J. Exp. Clin. Anat. 2015, 14, 18–23. [Google Scholar] [CrossRef]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Tabatabaei, S.N. Effect of olibanum (Boswellia thurifera) as a feed additive on performance, some blood biochemical and intestinal morphology in broiler chicks. Res. Opin. Anim. Vet. Sci. 2016, 6, 130–134. [Google Scholar]
- Gupta, I.; Parihar, A.; Malhotra, P.; Gupta, S.; Lüdtke, R.; Safayhi, H.; Ammon, H.P. Effects of gum resin of Boswellia serrata in patients with chronic colitis. Planta Med. 2001, 67, 391–395. [Google Scholar] [CrossRef]
- Krieglstein, C.F.; Anthoni, C.; Rijcken, E.J.; Laukötter, M.; Spiegel, H.-U.; Boden, S.E.; Schweizer, S.; Safayhi, H.; Senninger, N.; Schürmann, G. Acetyl-11-keto-β-boswellic acid, a constituent of a herbal medicine from Boswellia serrata resin, attenuates experimental ileitis. Int. J. Color. Dis. 2001, 16, 88–95. [Google Scholar] [CrossRef]
- Al-Yasiry, A.; Kiczorowska, B.; Samolińska, W.; Kowalczuk-Vasilev, E.; Kowalczyk-Pecka, D. The effect of Boswellia serrata resin diet supplementation on production, hematological, biochemical and immunological parameters in broiler chickens. Animal 2017, 11, 1890–1898. [Google Scholar] [CrossRef]
- Mohamed, S.H.; Attia, A.I.; Reda, F.M.; Abd El-Hack, M.E.; Ismail, I.E. Impacts of dietary supplementation of Boswellia serrata on growth, nutrients digestibility, immunity, antioxidant status, carcase traits and caecum microbiota of broilers. Ital. J. Anim. Sci. 2021, 20, 205–214. [Google Scholar] [CrossRef]
- Ismail, I.E.; Abdelnour, S.A.; Shehata, S.A.; Abd El-Hack, M.E.; El-Edel, M.A.; Taha, A.E.; Schiavitto, M.; Tufarelli, V. Effect of dietary Boswellia serrata resin on growth performance, blood biochemistry, and cecal microbiota of growing rabbits. Front. Vet. Sci. 2019, 6, 471. [Google Scholar] [CrossRef]
- Al-Yasiry, A.R.M.; Kiczorowska, B.; Samolińska, W.; Kowalczuk-Vasilev, E. Growth performance, digestibility, haematology, biochemistry, and some humoral immunity blood parameters of broiler chickens fed different levels of Boswellia serrata resin. Anim. Prod. Sci. 2017, 58, 1885–1891. [Google Scholar] [CrossRef]
- Tabatabaei, S.N.; Modaresi, M.; Moatar, F.; Pirestani, A.; Tavalaeean, E. Effect of different levels of Boswellia serrata Triana & Planch on immune responses of broilers chicks. J. Herb. Drugs (Int. J. Med. Herbs) 2012, 3, 49–54. [Google Scholar]
- Smet, K.; Raes, K.; Huyghebaert, G.; Haak, L.; Arnouts, S.; De Smet, S. Lipid and protein oxidation of broiler meat as influenced by dietary natural antioxidant supplementation. Poult. Sci. 2008, 87, 1682–1688. [Google Scholar] [CrossRef]
- Amer, S.A.; Mohamed, W.A.; Gharib, H.S.; Al-Gabri, N.A.; Gouda, A.; Elabbasy, M.T.; El-Rahman, A.; Ghada, I.; Omar, A.E. Changes in the growth, ileal digestibility, intestinal histology, behavior, fatty acid composition of the breast muscles, and blood biochemical parameters of broiler chickens by dietary inclusion of safflower oil and vitamin C. BMC Vet. Res. 2021, 17, 68. [Google Scholar] [CrossRef]
- Kishawy, A.T.; Amer, S.A.; Abd El-Hack, M.E.; Saadeldin, I.M.; Swelum, A.A. The impact of dietary linseed oil and pomegranate peel extract on broiler growth, carcass traits, serum lipid profile, and meat fatty acid, phenol, and flavonoid contents. Asian-Australas. J. Anim. Sci. 2019, 32, 1161. [Google Scholar] [CrossRef]
- Dhama, K.; Latheef, S.K.; Mani, S.; Samad, H.A.; Karthik, K.; Tiwari, R.; Khan, R.U.; Alagawany, M.; Farag, M.R.; Alam, G.M. Multiple beneficial applications and modes of action of herbs in poultry health and production-a review. Int. J. Pharmacol. 2015, 11, 152–176. [Google Scholar] [CrossRef]
- Kiczorowska, B.; Samolińska, W.; Al-Yasiry, A.; Zając, M. Immunomodulant feed supplement Boswellia serrata to support broiler chickens’ health and dietary and technological meat quality. Poult. Sci. 2020, 99, 1052–1061. [Google Scholar] [CrossRef]
- Gou, Z.; Cui, X.; Li, L.; Fan, Q.; Lin, X.; Wang, Y.; Jiang, Z.; Jiang, S. Effects of dietary incorporation of linseed oil with soybean isoflavone on fatty acid profiles and lipid metabolism-related gene expression in breast muscle of chickens. Animal 2020, 14, 2414–2422. [Google Scholar] [CrossRef]
- Zanetti, E.; De Marchi, M.; Dalvit, C.; Molette, C.; Rémignon, H.; Cassandro, M. Carcase characteristics and qualitative meat traits of three Italian local chicken breeds. Br. Poult. Sci. 2010, 51, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Nkukwana, T.T.; Muchenje, V.; Masika, P.; Hoffman, L.C.; Dzama, K.; Descalzo, A.M. Fatty acid composition and oxidative stability of breast meat from broiler chickens supplemented with Moringa oleifera leaf meal over a period of refrigeration. Food Chem. 2014, 142, 255–261. [Google Scholar] [CrossRef]
- Mpofu, D.A.; Marume, U.; Mlambo, V.; Hugo, A. The effects of Lippia javanica dietary inclusion on growth performance, carcass characteristics and fatty acid profiles of broiler chickens. Anim. Nutr. 2016, 2, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Hou, D.-X. The biofunctions of phytochemicals and their applications in farm animals: The Nrf2/Keap1 system as a target. Engineering 2017, 3, 738–752. [Google Scholar] [CrossRef]
- Kiczorowska, B.; Samolińska, W.; Al-Yasiry, A.R.M.; Kowalczyk-Pecka, D. Effect of dietary supplementation on growth performance, gastrointestinal microflora, and morphology of broilers. Ann. Anim. Sci. 2016, 16, 835–849. [Google Scholar] [CrossRef]
- Pluske, J.R.; Thompson, M.J.; Atwood, C.S.; Bird, P.H.; Williams, I.H.; Hartmann, P.E. Maintenance of villus height and crypt depth, and enhancement of disaccharide digestion and monosaccharide absorption, in piglets fed on cows’ whole milk after weaning. Br. J. Nutr. 1996, 76, 409–422. [Google Scholar] [CrossRef]
- Samanya, M.; Yamauchi, K.-e. Histological alterations of intestinal villi in chickens fed dried Bacillus subtilis var. natto. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2002, 133, 95–104. [Google Scholar] [CrossRef]
- Oster, M.; Gerlinger, C.; Heide, K.; Just, F.; Borgelt, L.; Wolf, P.; Polley, C.; Vollmar, B.; Muráni, E.; Ponsuksili, S. Lower dietary phosphorus supply in pigs match both animal welfare aspects and resource efficiency. Ambio 2018, 47, 20–29. [Google Scholar] [CrossRef]
- Nari, N.; Ghasemi, H.A. Growth performance, nutrient digestibility, bone mineralization, and hormone profile in broilers fed with phosphorus-deficient diets supplemented with butyric acid and Saccharomyces boulardii. Poult. Sci. 2020, 99, 926–935. [Google Scholar] [CrossRef]
- Waters, M.J.; Brooks, A.J. Growth hormone and cell growth. Dev. Biol. GH Secret. Growth Treat. 2012, 23, 86–95. [Google Scholar]
- Liu, Z.; Cordoba-Chacon, J.; Kineman, R.D.; Cronstein, B.N.; Muzumdar, R.; Gong, Z.; Werner, H.; Yakar, S. Growth hormone control of hepatic lipid metabolism. Diabetes 2016, 65, 3598–3609. [Google Scholar] [CrossRef]
- Blas, J.; Scanes, C. Sturkie’s Avian Physiology; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Frankincense, D.L. Effect of supplementation of different levels frankincense to drinking water on certain hematological traits of broiler. Environ. Sci 2013, 8, 589–601. [Google Scholar]
- Guo, Y.; Chen, S.; Xia, Z.; Yuan, J. Effects of different types of polyunsaturated fatty acids on immune function and PGE2 synthesis by peripheral blood leukocytes of laying hens. Anim. Feed. Sci. Technol. 2004, 116, 249–258. [Google Scholar] [CrossRef]
- Ryan, G. CD3 conformation is crucial for signalling. Nat. Rev. Immunol. 2010, 10, 7. [Google Scholar] [CrossRef]
- Tedder, T.F.; Engel, P. CD20: A regulator of cell-cycle progression of B lymphocytes. Immunol. Today 1994, 15, 450–454. [Google Scholar] [CrossRef]
- Takagi, Y.; Choi, I.-S.; Yamashita, T.; Nakamura, T.; Suzuki, I.; Hasegawa, T.; Oshima, M.; Gu, Y.-H. Immune activation and radioprotection by propolis. Am. J. Chin. Med. 2005, 33, 231–240. [Google Scholar] [CrossRef]
- Paguigan, N.D.; Rivera-Chávez, J.; Stempin, J.J.; Augustinović, M.; Noras, A.I.; Raja, H.A.; Todd, D.A.; Triplett, K.D.; Day, C.; Figueroa, M. Prenylated diresorcinols inhibit bacterial quorum sensing. J. Nat. Prod. 2019, 82, 550–558. [Google Scholar] [CrossRef]
- Wang, X.; He, H.; Liu, J.; Xie, S.; Han, J. Inhibiting roles of farnesol and HOG in morphological switching of Candida albicans. Am. J. Transl. Res. 2020, 12, 6988. [Google Scholar]
- Sachivkina, N.; Vasilieva, E.; Lenchenko, E.; Kuznetsova, O.; Karamyan, A.; Ibragimova, A.; Zhabo, N.; Molchanova, M. Reduction in pathogenicity in yeast-like fungi by farnesol in quail model. Animals 2022, 12, 489. [Google Scholar] [CrossRef]
- Hallen-Adams, H.E.; Suhr, M.J. Fungi in the healthy human gastrointestinal tract. Virulence 2017, 8, 352–358. [Google Scholar] [CrossRef]
- Rudenko, P.; Sachivkina, N.; Vatnikov, Y.; Shabunin, S.; Engashev, S.; Kontsevaya, S.; Karamyan, A.; Bokov, D.; Kuznetsova, O.; Vasilieva, E. Role of microorganisms isolated from cows with mastitis in Moscow region in biofilm formation. Vet. World 2021, 14, 40. [Google Scholar] [CrossRef] [PubMed]
- Singkum, P.; Muangkaew, W.; Suwanmanee, S.; Pumeesat, P.; Wongsuk, T.; Luplertlop, N. Suppression of the pathogenicity of Candida albicans by the quorum-sensing molecules farnesol and tryptophol. J. Gen. Appl. Microbiol. 2019, 65, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Zawrotniak, M.; Wojtalik, K.; Rapala-Kozik, M. Farnesol, a quorum-sensing molecule of Candida albicans triggers the release of neutrophil extracellular traps. Cells 2019, 8, 1611. [Google Scholar] [CrossRef] [PubMed]
- Dixon, E.F.; Hall, R.A. Noisy neighbourhoods: Quorum sensing in fungal–polymicrobial infections. Cell. Microbiol. 2015, 17, 1431–1441. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant and anti-inflammatory activities of essential oils: A short review. Molecules 2010, 15, 9252–9287. [Google Scholar] [CrossRef]
- Schrier, D.; Baragi, V.; Connor, D.; Dyer, R.; Jordan, J.; Imre, K.; Lesch, M.; Mullican, M.; Okonkwo, G.; Conroy, M. The pharmacologic effects of 5-[3, 5-bis (1, 1-dimethylethyl)-4-hydroxyphenyl]-1, 3, 4-thiadiazole-2 (3H)-thione, choline salt (CI-986), a novel inhibitor of arachidonic acid metabolism in models of inflammation, analgesia and gastric irritation. Prostaglandins 1994, 47, 17–30. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R. Quorum sensing and phytochemicals. Int. J. Mol. Sci. 2013, 14, 12607–12619. [Google Scholar] [CrossRef]
- Ren, J.; Wang, J.; Karthikeyan, S.; Liu, H.; Cai, J. Natural anti-phytopathogenic fungi compound phenol, 2, 4-bis (1, 1-dimethylethyl) from Pseudomonas fluorescens TL-1. Indian J. Biochem. Biophys. 2019, 56, 162–168. [Google Scholar]
- Yoon, M.-A.; Jeong, T.-S.; Park, D.-S.; Xu, M.-Z.; Oh, H.-W.; Song, K.-B.; Lee, W.S.; Park, H.-Y. Antioxidant effects of quinoline alkaloids and 2, 4-di-tert-butylphenol isolated from Scolopendra subspinipes. Biol. Pharm. Bull. 2006, 29, 735–739. [Google Scholar] [CrossRef]
- Rangel-Sánchez, G.; Castro-Mercado, E.; García-Pineda, E. Avocado roots treated with salicylic acid produce phenol-2, 4-bis (1, 1-dimethylethyl), a compound with antifungal activity. J. Plant Physiol. 2014, 171, 189–198. [Google Scholar] [CrossRef]
- Abdullah, A.-S.H.; Mirghani, M.E.S.; Jamal, P. Antibacterial activity of Malaysian mango kernel. Afr. J. Biotechnol. 2011, 10, 18739–18748. [Google Scholar]
- Kumar, P.S.; Duraipandiyan, V.; Ignacimuthu, S. Isolation, screening and partial purification of antimicrobial antibiotics from soil Streptomyces sp. SCA 7. Kaohsiung J. Med. Sci. 2014, 30, 435–446. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Starter Period (4–10 d) | Grower Period (11–23 d) | Finisher Period (24–35 d) |
|---|---|---|---|
| Yellow corn | 55.9 | 59.3 | 62 |
| Corn gluten, 60% | 3.935 | 5.275 | 6.07 |
| Soybean meal, 48% | 33.42 | 28.1 | 23.825 |
| Soybean oil | 2.2 | 3 | 4 |
| Calcium dibasic phosphate | 1.5 | 1.4 | 1.3 |
| Calcium carbonate | 1.2 | 1.2 | 1.1 |
| Common salt | 0.15 | 0.15 | 0.15 |
| DL-methionine, 98% | 0.4 | 0.3 | 0.33 |
| Premix * | 0.3 | 0.3 | 0.3 |
| Lysine HCl, 78% | 0.47 | 0.45 | 0.40 |
| Phytase | 0.005 | 0.005 | 0.005 |
| Threonine | 0.1 | 0.1 | 0.1 |
| Choline | 0.07 | 0.07 | 0.07 |
| Antimycotoxin | 0.1 | 0.1 | 0.1 |
| Na2CO3 | 0.25 | 0.25 | 0.25 |
| Chemical composition | |||
| Crude protein% | 23.05 | 21.52 | 20.15 |
| ME (Kcal/kg) | 3004 | 3101 | 3201 |
| Calcium % | 0.941 | 0.904 | 0.832 |
| Available phosphorus % | 0.481 | 0.448 | 0.417 |
| Lysine % | 1.46 | 1.31 | 1.16 |
| Methionine % | 0.721 | 0.610 | 0.626 |
| Threonine % | 0.824 | 0.765 | 0.713 |
| Bioactive Compounds | Retention Time | Peak Area % |
|---|---|---|
| Farnesol | 43.53 | 12.42 |
| ç-Elemene | 43.53 | 12.42 |
| à-Farnesene | 43.53 | 12.42 |
| Phenol, bis(1,1-dimethylethyl) | 17.78 | 7.15 |
| Phenol, 2,4-bis(1,1-dimethylethyl) 2,4-di-tert-butylphenol | 17.78 | 7.15 |
| Phenol, 3,5-bis(1,1-dimethylethyl) | 17.78 | 7.15 |
| 3-Thujanol | 30.25 | 3.62 |
| Bicyclo [3.1.0] hexan-3-ol, 4-methyl-1-(1-methylethyl) | 30.25 | 3.62 |
| Methyl 11,12-tetradecadienoate | 30.25 | 3.62 |
| 10-Undecyn-1-ol | 30.56 | 3.50 |
| (3-Cyclopropylbicyclo [4.1.0] hept-7-Yl)methanol | 30.56 | 3.50 |
| E,E-6,11-Tridecadien-1-ol acetate | 30.56 | 3.50 |
| Bicyclo [4.1.0]heptane,-3-cyclopropyl,-7-hydroxymethyl, trans | 30.56 | 3.50 |
| Dichloroacetic acid, dodec-9-ynyl | 30.56 | 3.50 |
| Trait Measured | FO0 | FO200 | FO400 | FO600 | Regression | |
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| Initial BW (g) | 99.44 ± 0.01 | 99.38 ± 0.63 | 98.75 ± 0.01 | 99.17 ± 0.72 | 0.271 | 0.404 |
| Starter period (4–10 days) | ||||||
| BW (g) | 303.25 ± 24.11 b | 356.53 ± 14.02 a | 329.83 ± 15.13 ab | 335.28 ± 10.01 ab | 0.145 | 0.04 |
| BWG (g) | 203.81 ± 24.11 b | 257.15 ± 14.29 a | 231.08 ± 15.13 ab | 236.11 ± 10.66 ab | 0.141 | 0.04 |
| FI (g) | 271.85 ± 5.59 | 273.12 ± 7.60 | 258.96 ± 11.39 | 267.08 ± 4.16 | 0.189 | 0.462 |
| FCR | 1.34 ± 0.13 a | 1.06 ± 0.03 b | 1.13 ± 0.11 b | 1.13 ± 0.05 b | 0.039 | 0.024 |
| Grower period (11–23 days) | ||||||
| BW (g) | 1073.33 ± 50.07 b | 1272.00 ± 12.53 a | 1228.61 ± 55.09 a | 1289.06 ± 37.52 a | 0.001 | 0.022 |
| BWG (g) | 770.08 ± 29.98 b | 915.47 ± 23.17 a | 898.78 ± 63.99 a | 953.78 ± 27.99 a | 0.001 | 0.08 |
| FI (g) | 1205.19 ± 29.66 | 1216.04 ± 100.87 | 1125.21 ± 38.56 | 1175.83 ± 29.13 | 0.265 | 0.568 |
| FCR | 1.57 ± 0.03 a | 1.33 ± 0.15 b | 1.26 ± 0.12 b | 1.23 ± 0.01 b | 0.002 | 0.09 |
| Finisher period (24–35 days) | ||||||
| BW (g) | 1762.22 ± 56.13 b | 2209.72 ± 8.67 a | 2186.11 ± 59.12 a | 2161.67 ± 88.93 a | <0.01 | <0.01 |
| BWG (g) | 688.89 ± 13.84 c | 937.72 ± 21.09 ab | 957.50 ± 41.89 a | 872.61 ± 55.19 b | <0.01 | <0.01 |
| FI (g) | 1487.78 ± 125.59 | 1607.08 ± 134.10 | 1452.92 ± 53.16 | 1455.00 ± 66.14 | 0.362 | 0.345 |
| FCR | 2.16 ± 0.16 a | 1.71 ± 0.11 b | 1.52 ± 0.04 b | 1.67 ± 0.16 b | <0.01 | <0.01 |
| Overall performance | ||||||
| Final BW, g | 1762.22 ± 56.13 b | 2209.72 ± 8.67 a | 2186.11 ± 59.12 a | 2161.67 ± 88.93 a | <0.01 | <0.01 |
| Total BWG, g | 1662.78 ± 56.13 b | 2110.35 ± 8.23 a | 2087.36 ± 59.12 a | 2062.50 ± 89.64 a | <0.01 | <0.01 |
| Total FI, g | 2964.81 ± 159.15 | 3096.25 ± 238.20 | 2837.08 ± 78.07 | 2897.92 ± 58.05 | 0.273 | 0.697 |
| FCR | 1.78 ± 0.04 a | 1.47 ± 0.11 b | 1.36 ± 0.03 b | 1.41 ± 0.07 b | <0.01 | <0.01 |
| FO0 | FO200 | FO400 | FO600 | Regression | ||
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| Dressing % | 58.577 ± 0.53 | 57.573 ± 0.90 | 57.70 ± 0.35 | 58.13 ± 0.94 | 0.367 | 0.220 |
| Liver % | 2.66 ± 0.40 | 2.47 ± 0.34 | 2.38 ± 0.48 | 2.44 ± 0.44 | 0.512 | 0.620 |
| Spleen % | 0.13 ± 0.05 | 0.10 ± 0.02 | 0.12 ± 0.02 | 0.15 ± 0.08 | 0.635 | 0.302 |
| Intestine % | 5.43 ± 0.35 | 6.06 ± 0.43 | 5.91 ± 0.58 | 5.21 ± 0.42 | 0.518 | 0.05 |
| Gizzard % | 1.78 ± 0.31 | 2.24 ± 0.28 | 1.92 ± 0.08 | 1.73 ± 0.20 | 0.483 | 0.05 |
| Bursa % | 0.13 ± 0.02 | 0.15 ± 0.04 | 0.16 ± 0.03 | 0.11 ± 0.01 | 0.611 | 0.084 |
| FO0 | FO200 | FO400 | FO600 | Regression | ||
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| ALA % | 0.033 ±0.01 b | 0.053 ±0.01 a | 0.056 ±0.01 a | 0.066 ±0.01 a | 0.001 | 0.290 |
| EPA % | 0.026 ±0.01 c | 0.040 ±0.00 b | 0.046 ±0.01 b | 0.050 ±0.01 a | <0.01 | 0.067 |
| DPA % | 0.023 ±0.01 b | 0.040 ±0.01 a | 0.040 ±0.01 a | 0.046 ±0.01 a | <0.01 | 0.067 |
| DHA % | 0.016 ±0.01 b | 0.030 ±0.01 a | 0.036 ±0.01 a | 0.036 ±0.01 a | 0.001 | 0.050 |
| LA % | 0.826 ±0.06 | 0.890 ±0.10 | 0.863 ±0.01 | 0.860 ±0.01 | 0.656 | 0.374 |
| AA % | 1.14 ±0.04 b | 1.20 ±0.08 b | 1.28 ±0.01 a | 1.29 ±0.02 a | 0.002 | 0.393 |
| n-3 (%). | 0.096 ±0.02 b | 0.163 ±0.01a | 0.176 ±0.02a | 0.193 ±0.02 a | <0.01 | 0.03 |
| n-6 (%). | 1.97 ±0.05 c | 2.09 ±0.03 b | 2.15 ±0.01 a | 2.15 ±0.02 a | <0.01 | 0.01 |
| n-3: n-6 ratio | 0.049 ±0.01 b | 0.078 ±0.002 a | 0.082 ±0.01 a | 0.089 ±0.01 a | <0.01 | 0.04 |
| FO0 | FO200 | FO400 | FO600 | Regression | ||
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| Duodenum | ||||||
| VH µm | 690.68 ± 13.0 c | 814.78 ± 3.94 a | 765.07 ± 22.55 b | 663.11 ± 36.42 c | 0.05 | <0.01 |
| VW µm | 181.57 ± 3.73 a | 184.47 ± 4.00 a | 121.82 ± 3.05 c | 134.42 ± 3.08 b | <0.01 | 0.043 |
| CD µm | 140.56 ± 3.24 c | 183.42 ± 6.60 a | 150.04 ± 6.39 bc | 159.67 ± 3.45 b | 0.110 | <0.01 |
| MCT µm | 140.75 ± 2.10 c | 254.81 ± 10.90 a | 179.56 ± 5.404 b | 125.03 ± 2.88 d | <0.01 | <0.01 |
| Jejunum | ||||||
| VH µm | 811.11 ± 23.5 c | 862.11 ± 16.3 bc | 897.38 ± 45.85 b | 995.43 ± 12.77 a | <0.01 | 0.180 |
| VW µm | 115.36 ± 8.48 d | 266.06 ± 10.62 a | 158.32 ± 7.06 c | 247.52 ± 5.66 b | <0.01 | <0.01 |
| CD µm | 112.76 ± 3.06 c | 190.68 ± 14.98 a | 154.90 ± 7.56 b | 160.45 ± 4.78 b | 0.002 | <0.01 |
| MCT µm | 115.15 ± 3.42 b | 166.06 ± 6.19 a | 116.28 ± 2.50 a | 118.98 ± 2.81 a | 0.006 | <0.01 |
| Ileum | ||||||
| VH µm | 676.29 ± 30.4 b | 522.81 ± 8.32 c | 765.23 ± 12.76 a | 474.49 ± 14.72 d | <0.01 | <0.01 |
| VW µm | 111.73 ± 3.72 c | 126.89 ± 10.06 b | 210.60 ± 7.63 a | 205.00 ± 6.98 a | <0.01 | 0.04 |
| CD µm | 106.89 ± 2.92 c | 165.94 ± 7.50 a | 134.92 ± 5.83 b | 137.26 ± 7.80 b | 0.006 | <0.01 |
| MCT µm | 147.36 ± 2.63 b | 301.06 ± 29.31 a | 112.81 ± 2.63 c | 139.43 ± 4.54 bc | 0.001 | <0.01 |
| FO0 | FO200 | FO400 | FO600 | Regression | ||
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| Total proteins (g/dL) | 3.393 ± 0.27 d | 3.983 ± 0.06 c | 4.453 ± 0.36 b | 5.240 ± 0.13 a | <0.01 | 0.497 |
| Albumin (g/dL) | 1.250 ± 0.03 b | 1.347 ± 0.01 b | 1.730 ± 0.39 ab | 1.943 ± 0.44 a | 0.012 | 0.739 |
| Total globulins (g/dL) | 2.143 ± 0.026 c | 2.637 ± 0.06 bc | 2.723 ± 0.06 b | 3.297 ± 0.51 a | 0.001 | 0.816 |
| GH (ng/mL) | 2.73 ± 0.35 c | 4.03 ± 0.47 b | 5.00 ± 0.36 a | 5.23 ± 0.40 a | <0.01 | 0.050 |
| T3 (ng/mL) | 3.39 ± 0.17 c | 4.26 ± 0.06 b | 4.69 ± 0.36 ab | 5.10 ± 0.35 a | <0.01 | 0.176 |
| T4 (ng/mL) | 19.41 ± 0.21 d | 21.65 ± 0.91 c | 23.31 ± 0.89 b | 25.35 ± 0.79 a | <0.01 | 0.822 |
| Glucose (mg/dL) | 336.67 ± 2.08 | 340.33 ± 5.13 | 340.00 ± 3.61 | 341.33 ± 3.06 | 0.184 | 0.594 |
| Leptin (ng/mL) | 2.10 ± 0.06 | 1.92 ± 0.24 | 1.94 ± 0.19 | 2.20 ± 0.04 | 0.440 | 0.050 |
| FO0 | FO200 | FO400 | FO600 | Regression | ||
|---|---|---|---|---|---|---|
| Linear | Quadratic | |||||
| Lysozyme enzyme (µg/mL) | 133.00 ±2.65 c | 172.67 ±14.43 b | 184.00 ±8.00 ab | 192.00 ±1.00 a | <0.01 | 0.011 |
| Complement 3 (g/L) | 1.063 ±0.02 d | 1.190 ±0.02 c | 1.257 ±0.02 b | 1.293 ±0.02 a | <0.01 | 0.003 |
| IL10 (ug/mL) | 1.733 ±0.51 c | 3.500 ±0.35 b | 3.967 ±0.21 ab | 4.400 ±0.26 a | <0.01 | 0.011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amer, S.A.; Gouda, A.; Saleh, G.K.; Nassar, A.H.; Abdel-Warith, A.-W.A.; Younis, E.M.; Altohamy, D.E.; Kilany, M.S.; Davies, S.J.; Omar, A.E. Dietary Frankincense (Boswellia serrata) Oil Modulates the Growth, Intestinal Morphology, the Fatty Acid Composition of Breast Muscle, Immune Status, and Immunoexpression of CD3 and CD20 in Broiler Chickens. Animals 2023, 13, 971. https://doi.org/10.3390/ani13060971
Amer SA, Gouda A, Saleh GK, Nassar AH, Abdel-Warith A-WA, Younis EM, Altohamy DE, Kilany MS, Davies SJ, Omar AE. Dietary Frankincense (Boswellia serrata) Oil Modulates the Growth, Intestinal Morphology, the Fatty Acid Composition of Breast Muscle, Immune Status, and Immunoexpression of CD3 and CD20 in Broiler Chickens. Animals. 2023; 13(6):971. https://doi.org/10.3390/ani13060971
Chicago/Turabian StyleAmer, Shimaa A., Ahmed Gouda, Gehan K. Saleh, Arwa H. Nassar, Abdel-Wahab A. Abdel-Warith, Elsayed M. Younis, Dalia E. Altohamy, Maha S. Kilany, Simon J. Davies, and Anaam E. Omar. 2023. "Dietary Frankincense (Boswellia serrata) Oil Modulates the Growth, Intestinal Morphology, the Fatty Acid Composition of Breast Muscle, Immune Status, and Immunoexpression of CD3 and CD20 in Broiler Chickens" Animals 13, no. 6: 971. https://doi.org/10.3390/ani13060971