
Antimicrobial Resistance in Physiological and Potentially Pathogenic Bacteria Isolated in Southern Italian Bats
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains Used
2.2. Antimicrobial Susceptibility Testing
2.3. Data Analysis and Visualization
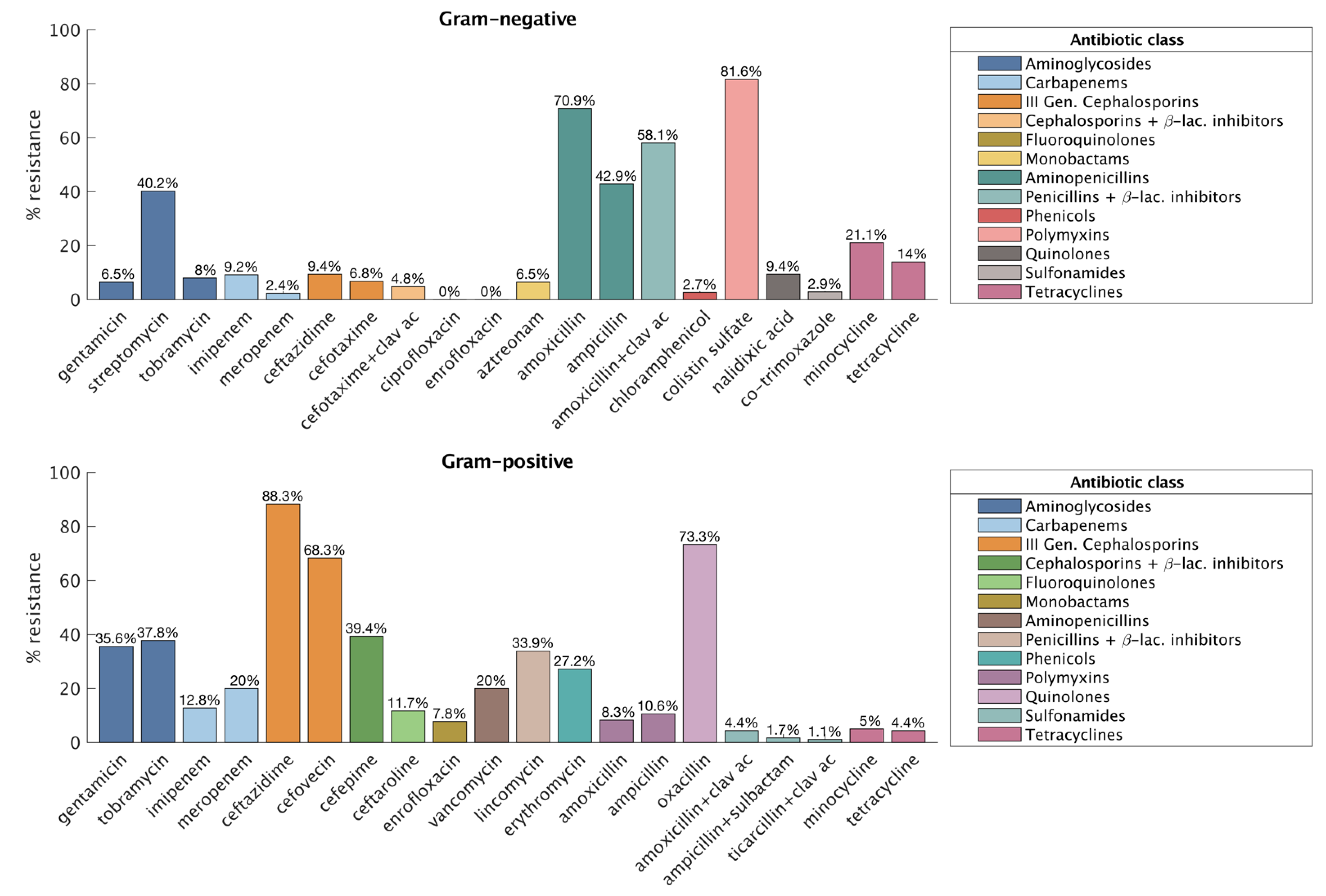
3. Results
3.1. Gram-Negative
3.2. Gram-Positive
3.3. Comparisons between Sites
3.4. Comparisons between Bat Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Banerjee, A.; Kulcsar, K.; Misra, V.; Frieman, M.; Mossman, K. Bats and Coronaviruses. Viruses 2019, 11, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochani, R.K.; Batra, S.; Shaikh, A.; Asad, A. Nipah virus—The rising epidemic: A review. Le Infez. Med. 2019, 27, 117–127. [Google Scholar]
- Rupprecht, C.E.; Turmelle, A.; Kuzmin, I.V. A perspective on lyssavirus emergence and perpetuation. Curr. Opin. Virol. 2011, 1, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühldorfer, K. Bats and Bacterial Pathogens: A Review. Zoonoses Public Health 2013, 60, 93–103. [Google Scholar] [CrossRef]
- Banskar, S.; Bhute, S.S.; Suryavanshi, M.V.; Punekar, S.; Shouche, Y.S. Microbiome analysis reveals the abundance of bacterial pathogens in Rousettus leschenaultii guano. Sci. Rep. 2016, 6, 36948. [Google Scholar] [CrossRef] [Green Version]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Carroll, D.; Wang, J.; Fanning, S.; McMahon, B.J. Antimicrobial resistance in wildlife: Implications for public health. Zoonoses Public Health 2015, 62, 534–542. [Google Scholar] [CrossRef]
- Arnold, K.E.; Williams, N.J.; Bennett, M. ‘Disperse abroad in the land’: The role of wildlife in the dissemination of antimicrobial resistance. Biol. Lett. 2016, 12, 20160137. [Google Scholar] [CrossRef] [Green Version]
- Swift, B.M.C.; Bennett, M.; Waller, K.; Dodd, C.; Murray, A.; Gomes, R.L.; Humphreys, B.; Hobman, J.L.; Jones, M.A.; Whitlock, S.E.; et al. Anthropogenic environmental drivers of antimicrobial resistance in wildlife. Sci. Total Environ. 2019, 649, 12–20. [Google Scholar] [CrossRef]
- Martinez, J.L. Environmental pollution by antibiotics and by antibiotic resistance determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef]
- Wellington, E.M.; Boxall, A.B.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Atterby, C.; Ramey, A.M.; Hall, G.G.; Järhult, J.; Börjesson, S.; Bonnedahl, J. Increased prevalence of antibiotic-resistant E. coli in gulls sampled in Southcentral Alaska is associated with urban environments. Infect. Ecol. Epidemiol. 2016, 6, 32334. [Google Scholar] [CrossRef] [Green Version]
- Devnath, P.; Karah, N.; Graham, J.P.; Rose, E.S.; Asaduzzaman, M. Evidence of Antimicrobial Resistance in Bats and Its Planetary Health Impact for Surveillance of Zoonotic Spillover Events: A Scoping Review. Int. J. Environ. Res. Public Health 2023, 20, 243. [Google Scholar] [CrossRef]
- Mbehang Nguema, P.P.; Onanga, R.; Ndong Atome, G.R.; Obague Mbeang, J.C.; Mabika Mabika, A.; Yaro, M.; Lounnas, M.; Dumont, Y.; Zohra, Z.F.; Godreuil, S.; et al. Characterization of ESBL-Producing Enterobacteria from Fruit Bats in an Unprotected Area of Makokou, Gabon. Microorganisms 2020, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Cláudio, V.C.; Gonzalez, I.; Barbosa, G.; Rocha, V.; Moratelli, R.; Rassy, F. Bacteria richness and antibiotic-resistance in bats from a protected area in the Atlantic Forest of Southeastern Brazil. PLoS ONE 2018, 13, e0203411. [Google Scholar] [CrossRef] [Green Version]
- Sherley, M.; Gordon, D.M.; Collignon, P.J. Variations in antibiotic resistance profile in Enterobacteriaceae isolated from wild Australian mammals. Environ. Microbiol. 2000, 2, 620–631. [Google Scholar] [CrossRef]
- Foti, M.; Spena, M.T.; Fisichella, V.; Mascetti, A.; Colnaghi, M.; Grasso, M.; Piraino, C.; Sciurba, F.; Grasso, R. Physiological and potentially pathogenic microbial flora in troglophile bats. Animals 2022, 12, 2684. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.M.M.; Sherris, J.C.; Turck, M.A. Antibiotic susceptibility testing by a standardised single disc method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Foti, M.; Grasso, R.; Fisichella, V.; Mascetti, A.; Zafarana, M.A.; Colnaghi, M.; Grasso, M.; Spena, M.T. Analysis of Eurasian Stone curlew (Burhinus oedicnemus) microbial flora reveals the presence of multi-drug resistant pathogens in agro-pastoral areas of Sicily (Italy). Heliyon 2020, 6, e05401. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.G. Simultaneous Statistical Inference; Springer: New York, NY, USA, 1966. [Google Scholar]
- Hoffmann, H. Violin.m—Simple Violin Plot Using Matlab Default Kernel Density Estimation; INRES (University of Bonn): Bonn, Germany, 2015. [Google Scholar]
- Thaller, M.C.; Migliore, L.; Marquez, C.; Tapia, W.; Cedeno, V.; Rossolini, G.M.; Gentile, G. Tracking acquired antibiotic resistance in commensal bacteria of Galapagos land iguanas: No man, no resistance. PLoS ONE 2010, 5, e8989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhouani, H.; Silva, N.; Poeta, P.; Torres, C.; Correia, S.; Igrejas, G. Potential impact of antimicrobial resistance in wildlife, environment and human health. Front. Microbiol. 2014, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, V.; Rocha, M.; Valera, A.; Eguiarte, L.E. Genetic structure of natural populations of Escherichia coli in wild hosts on different continents. Appl. Environ. Microbiol. 1999, 65, 3373–3385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassell, J.M.; Ward, M.J.; Muloi, D.; Bettridge, J.M.; Robinson, T.P.; Kariuki, S.; Ogendo, A.; Kiiru, J.; Imboma, T.; Kang’ethe, E.K. Clinically relevant antimicrobial resistance at the wildlife–livestock–human interface in Nairobi: An epidemiological study. Lancet Planet. Health 2019, 3, e259–e269. [Google Scholar] [CrossRef] [Green Version]
- Benavides, J.A.; Shiva, C.; Virhuez, M.; Tello, C.; Appelgren, A.; Vendrell, J.; Solassol, J.; Godreuil, S.; Streicker, D.G. Extended-spectrum beta-lactamase-producing Escherichia coli in common vampire bats Desmodus rotundus and livestock in Peru. Zoonoses Public Health 2018, 65, 454–458. [Google Scholar] [CrossRef] [Green Version]
- Allocati, N.; Petrucci, A.G.; Di Giovanni, P.; Masulli, M.; Di Ilio, C.; De Laurenzi, V. Bat–man disease transmission: Zoonotic pathogens from wildlife reservoirs to human populations. Cell Death Discov. 2016, 2, 16048. [Google Scholar] [CrossRef] [Green Version]
- George, A.M.; Hall, R.M.; Stokes, H.W. Multidrug resistance in Klebsiella pneumoniae: A novel gene, ramA, confers a multidrug resistance phenotype in Escherichia coli. Microbiology 1995, 141, 1909–1920. [Google Scholar] [CrossRef] [Green Version]
- Nowakiewicz, A.; Ziębab, P.; Gnata, S.; Trościańczyka, A.; Osińskaa, M.; Łagowskia, D.; Kosior-Korzeckac, U.; Puziod, I. Bats as a reservoir of resistant Escherichia coli: A methodical view. Can we fully estimate the scale of resistance in the reservoirs of free-living animals? Res. Vet. Sci. 2020, 128, 49–58. [Google Scholar] [CrossRef]
- Concannon, R.; Wynn-Owen, K.; Simpson, V.R.; Birtles, R.J. Molecular characterization of haemoparasites infecting bats (Microchiroptera) in Cornwall, UK. Parasitology 2005, 131, 489–496. [Google Scholar] [CrossRef]
- Evans, N.J.; Bown, K.; Timofte, D.; Simpson, V.R.; Birtles, R.J. Fatal borreliosis in bat caused by relapsing fever spirochete, United Kingdom. Emerg. Infect. Dis. 2009, 15, 1331–1333. [Google Scholar] [CrossRef]
- Veikkolainen, V.; Vesterinen, E.J.; Lilley, T.M.; Pulliainen, A.T. Bats as reservoir hosts of human bacterial pathogen, Bartonella mayotimonensis. Emerg. Infect. Dis. 2014, 20, 960–967. [Google Scholar] [CrossRef]
- Millán, J.; López-Roig, M.; Delicado, V.; Serra-Cobo, J.; Esperón, F. Widespread infection with hemotropic mycoplasmas in bats in Spain, including a hemoplasma closely related to “Candidatus Mycoplasma hemohominis”. Comp. Immunol. Microbiol. Infect. Dis. 2015, 39, 9–12. [Google Scholar] [CrossRef]
- Bai, Y.; Urushadze, L.; Osikowicz, L.; McKee, C.; Kuzmin, I.; Kandaurov, A.; Babuadze, G.; Natradze, I.; Imnadze, P.; Kosoy, M. Molecular Survey of Bacterial Zoonotic Agents in Bats from the Country of Georgia, Caucasus. PLoS ONE 2017, 12, e0171175. [Google Scholar] [CrossRef]
- Stuckey, M.J.; Boulouis, H.J.; Cliquet, F.; Picard-Meyer, E.; Servat, A.; Aréchiga-Ceballos, N.; Echevarría, J.E.; Chomel, B.B. Potentially Zoonotic Bartonella in Bats from France and Spain. Emerg. Infect. Dis. 2017, 23, 539–541. [Google Scholar] [CrossRef]
- Corduneanu, A.; Sándor, A.D.; Ionică, A.M.; Hornok, S.; Leitner, N.; Bagó, Z.; Stefke, K.; Fuehrer, H.P.; Mihalca, A.D. Bartonella DNA in heart tissues of bats in central and eastern Europe and a review of phylogenetic relations of bat-associated bartonellae. Parasites Vectors 2018, 11, 489. [Google Scholar] [CrossRef]
- Hornok, S.; Szőke, K.; Estók, P.; Krawczyk, A.; Haarsma, A.J.; Kováts, D.; Boldogh, S.A.; Morandini, P.; Szekeres, S.; Takács, N.; et al. Assessing bat droppings and predatory bird pellets for vector-borne bacteria: Molecular evidence of bat-associated Neorickettsia sp. in Europe. Antonie Van Leeuwenhoek 2018, 111, 1707–1717. [Google Scholar] [CrossRef]
- Vittecoq, M.; Godreuil, S.; Prugnolle, F.; Durand, P.; Brazier, L.; Renaud, N.; Arnal, A.; Aberkane, S.; Jean-Pierre, H.; Gauthie-Clerc, M.; et al. Antimicrobial resistance in wildlife. J. Appl. Ecol. 2016, 53, 519–529. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Superfamily | Family | Subfamily | Species | n. Individuals |
|---|---|---|---|---|
| Vespertilionoidea | Vespertilionoidae | Myotinae | Myotis myotis | 47 |
| Myotis capaccinii | 8 | |||
| Miniopteridae | Miniopterus schreibersii | 91 | ||
| Rhinolophoidea | Rhinolophidae | Rhinolophus hipposideros | 43 | |
| Total | 189 |
| Site | Number of Sampled Individuals | ||||
|---|---|---|---|---|---|
| Myotis myotis | Miniopterus schreibersii | Rhinolophus hipposideros | Myotis capaccinii | Total | |
| Grotta dei Pipistrelli Pantalica (SR) | 13 | 12 | 17 | 8 | 50 |
| Grotta Palombara (SR) | 5 | 9 | 14 | ||
| Grotta Chiusazza (SR) | 12 | 13 | 7 | 32 | |
| Grotta dei Pipistrelli Cassano (CS) | 16 | 15 | 31 | ||
| Grotta del Burrò (CT) | 1 | 15 | 17 | 33 | |
| Grave Grubbo (KR) | 27 | 2 | 29 | ||
| Total | 47 | 91 | 43 | 8 | 189 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foti, M.; Grasso, R.; Fisichella, V.; Mascetti, A.; Colnaghi, M.; Grasso, M.; Spena, M.T. Antimicrobial Resistance in Physiological and Potentially Pathogenic Bacteria Isolated in Southern Italian Bats. Animals 2023, 13, 966. https://doi.org/10.3390/ani13060966
Foti M, Grasso R, Fisichella V, Mascetti A, Colnaghi M, Grasso M, Spena MT. Antimicrobial Resistance in Physiological and Potentially Pathogenic Bacteria Isolated in Southern Italian Bats. Animals. 2023; 13(6):966. https://doi.org/10.3390/ani13060966
Chicago/Turabian StyleFoti, Maria, Rosario Grasso, Vittorio Fisichella, Antonietta Mascetti, Marco Colnaghi, Maria Grasso, and Maria Teresa Spena. 2023. "Antimicrobial Resistance in Physiological and Potentially Pathogenic Bacteria Isolated in Southern Italian Bats" Animals 13, no. 6: 966. https://doi.org/10.3390/ani13060966