
A Monogenean Gill Parasite within the Genus Haliotrema (Ancyrocephalidae) Infecting Argyrops filamentosus Fish: Morphology and Molecular Studies
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Collection
2.2. Parasitological Examination
2.3. Morphological Methods
2.3.1. Light Microscopic (LM) Study
2.3.2. Morphology and Morphometry
2.4. Molecular Analysis
2.4.1. DNA Extraction and PCR Amplification
2.4.2. Sequence Alignment and Molecular Phylogenetic Analysis
3. Results
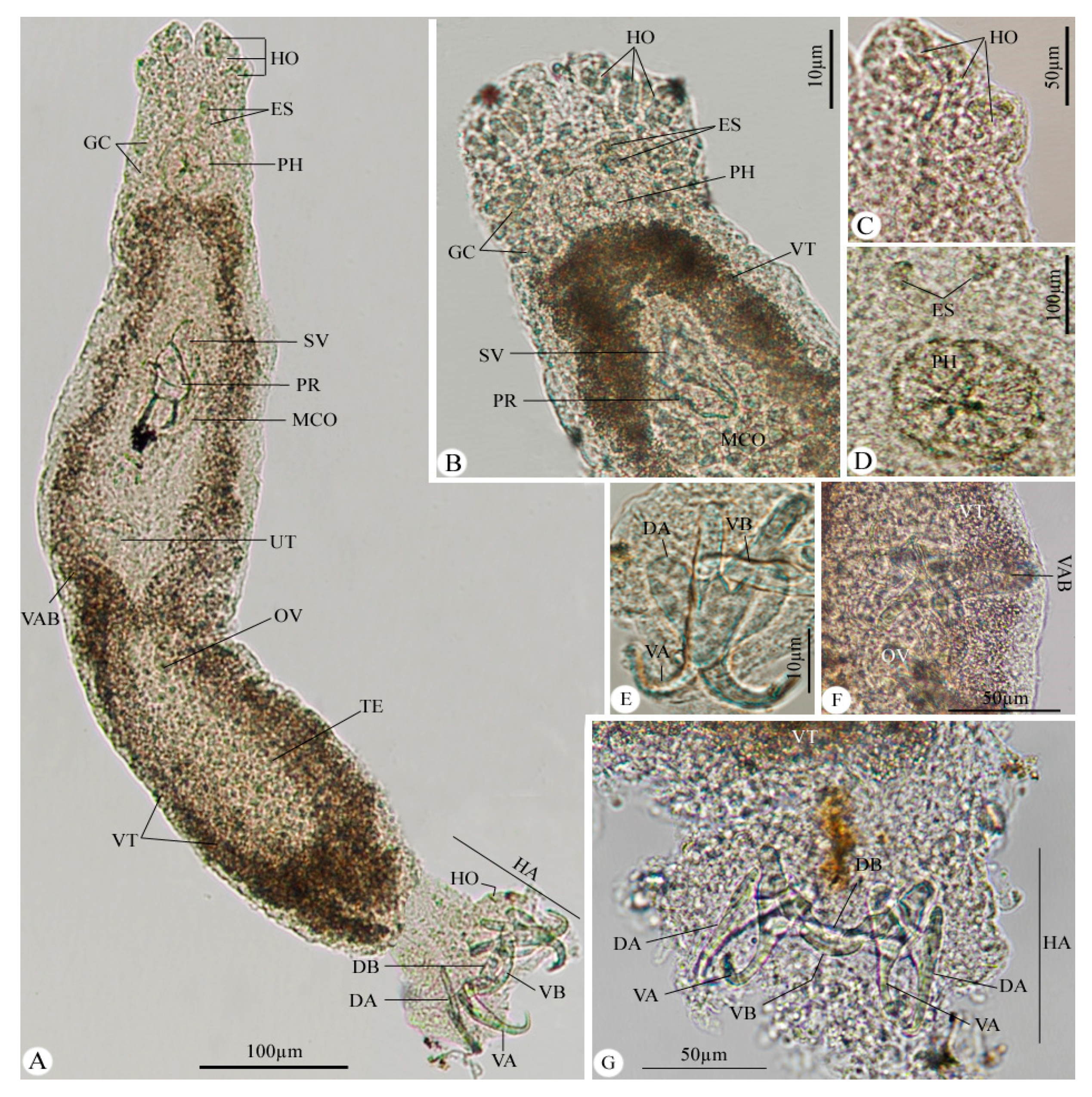
3.1. Microscopic Examinations (Figure 1)

3.2. Remarks
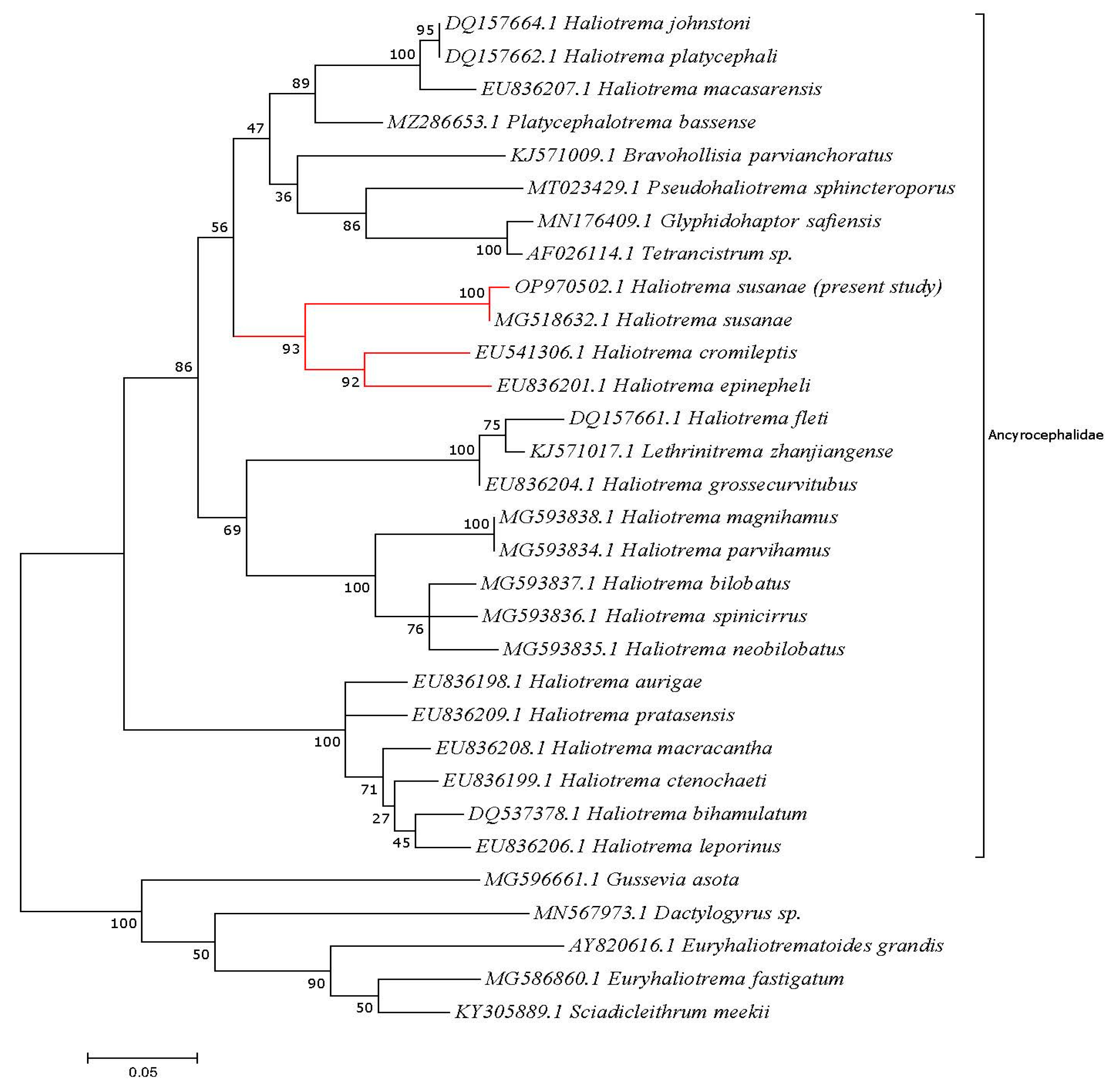
3.3. Molecular Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdel-Gaber, R.; Al Quraishy, S.; Dkhil, M.A.; Elamin, M.; Alghamdi, M. Morphological analysis of Caligus elongatus von Nordmann, 1832 (Copepoda: Caligidae) from the rosy goatfish Parupeneus rubescens (Mullidae). Micro. Res. Tech. 2020, 83, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Machkewskyi, V.K.; Dmitrieva, E.V.; Al-Jufaili, S.; Al-Mazrooei, N.A.M. Microcotyle omanae n. sp. (Monogenea: Microcotylidae), a parasite of Cheimerius nufar (Valenciennes) (Sparidae) from the Arabian Sea. Sys. Parasitol. 2013, 86, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Jayabalan, N.; Al-Marzouqi, A.; Al-Nahdi, A. Reproductive biology of the soldierbream, Argyrops filamentosus (Valenciennes, 1830) from the Arabian Sea coast of Oman. Indian J. Fish. 2011, 58, 9–17. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Catalog of Fishes: Genera, Species, References; California Academy of Sciences: San Francisco, CA, USA, 2018; Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 1 April 2022).
- Ghanem, R.; Jamila, B.S.; Azzurro, E. Documented occurrence of the exotic soldierbream Argyrops filamentosus (Valenciennes, 1830) in the specially protected area of Zembra (Tunisia): A result of a joint monitoring strategy. Cah. De Biol. Mar. 2021, 62, 227–233. [Google Scholar]
- Basurco, B.; Lovatelli, A.; Garcia, B. Current status of Sparidae aquaculture. In Sparidae: Biology and Aquaculture of Gilhead Sea Bream and Other Species; Parlidis, M.A., Mylonas, C.C., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2011; pp. 1–50. [Google Scholar]
- Bakke, T.A.; Cable, J.; Harris, P.D. The biology of gyrodactylid monogeneans: The Russian-doll killers. Adv. Parasitol. 2009, 64, 161–218. [Google Scholar]
- Wong, W.L.; Gorb, S. Attachment ability of a clamp-bearing fish parasite, Diplozoon paradoxum (Monogenea), on gills of the common bream, Abramis brama. J. Exp. Biol. 2013, 216, 3008–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shini, K.K.; Preethakumari, V.M.; Vasandakumar, M.V. Diversity of Monogenean (Platyhelminthes) Parasites in Freshwater Fishes of Kannur District, Kerala, India. Int. J. Rec. Res. Life Sci. 2015, 2, 54–57. [Google Scholar]
- WoRMS. Haliotrema cromileptis Young. 1968. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=518098 (accessed on 27 December 2022).
- Johnston, T.H.; Tiegs, O.W. New gyrodactyloid trematodes from Australian fishes together with a reclassification of the super-family Gyrodactyloidea. Linn. Soc. New South Wales 1922, 47, 83–131. [Google Scholar]
- Young, P.C. Ten new species of Haliotrema (Monogenoidea: Dactylogyridae) from Australian fish and a revision of the genus. J. Zool. 1968, 154, 41–75. [Google Scholar] [CrossRef]
- Soo, O.Y.M. A new species of Haliotrema (Monogenea: Ancyrocephalidae (sensu lato) Bychowsky & Nagibina, 1968) from holocentrids off Langkawi Island, Malaysia with notes on the phylogeny of related Haliotrema species. Parasitol. Int. 2019, 68, 31–39. [Google Scholar] [PubMed]
- LaBlack, L.; Rima, M.; Georgieva, S.; Marzoug, D.; Kostadinova, A. Novel molecular data for monogenean parasites of sparid fishes in the Mediterranean and a molecular phylogeny of the Microcotylidae Taschenberg, 1879. CRPVBD 2022, 2, 100069. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Y.; Zhu, X.Q.; Xie, M.Q.; Li, X. The radiation of Haliotrema (Monogenea: Dactylogyridae: Ancyrocephalinae): Molecular evidence and explanation inferred from LSU rDNA sequences. Parasitology 2006, 132, 659–668. [Google Scholar] [CrossRef]
- Abu Shusha, T.L.; Kalantan, M.Z.; Al-Nazry, H.H.; Al-Ghamdi, Y.A. Fishes from Territorial Saudi Red Sea Waters; Marine Research Centre-Jeddah, Deputy Ministry of Fisheries Affairs, Ministry of Agriculture: Riyadh, Saudi Arabia, 2010.
- Hassan, M.A.; Osman, H.A.M.; Aswathan, M.; Al-Shwared, W.A.; Fita, N.A. Infestation of Cage-Cultured Marine Fish with Benedenia acanthopagri (Monogenea; Capsalidae) in Eastern Province of Saudi Arabia. Glob. Vet. 2015, 14, 219–227. [Google Scholar]
- Amine, F.; Euzet, L.; Kechemir-Issad, N. Description de Lamellodiscus confusus n. sp. (Monogenea: Diplectanidae) parasite de Sarpa salpa (Teleostei: Sparidae). Parasite 2007, 14, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Kritsky, D.C.; Klimpel, S. Cyclocotyloides bergstadi n. sp. (Monogenoidea: Diclidophoridae: Diclidophoropsinae) from the gills of grenadier, Coryphaenoides brevibarbis (Teleostei: Macrouridae), in the Northeast Atlantic Ocean. Comp. Parasitol. 2007, 74, 23–30. [Google Scholar] [CrossRef]
- Al-Zubaidy, A.B. The first record of two polyopisthocotylean monogeneans on gills of the great barracuda, Sphyraena barracuda from the Red Sea, Yemeni coastal waters. J. King Abdulaziz Uni. Mar. Sci. 2013, 24, 3–14. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Littlewood, D.T.J.; Curini-Galletti, M.; Herniou, E.A. The interrelationships of Proseriata (Platyhelminthes: Seriata) tested with molecules and morphology. Mol. Phylogenet. Evol. 2000, 16, 449–466. [Google Scholar] [CrossRef]
- Tkach, V.V.; Littlewood, D.T.J.; Olson, P.D.; Kinsella, J.M.; Swiderski, Z. Molecular phylogenetic analysis of the Microphalloidea Ward, 1901 (Trematoda: Digenea). Sys. Parasitol. 2003, 56, 1–15. [Google Scholar] [CrossRef]
- Zhukov, E.V. Monogenea of the genus Haliotrema Johnston et Tiegs, 1922 from the gills of fishes family Holocentridae and family Acanthuridae of the Gulf of Mexico. Parazitologicheskii sbornik 1980, 29, 41–52. [Google Scholar]
- Klassen, G.J. Phylogeny of Haliotrema species (Monogenea: Ancyrocephalidae) from boxfishes (Tetraodontiformes: Ostraciidae): Are Haliotrema species from boxfishes monophyletic? J. Parasitol. 1994, 80, 596–610. [Google Scholar] [CrossRef] [PubMed]
- Kritsky, D.C.; Stephens, F. Haliotrema abaddon n. sp. (Monogenoidea: Dactylogyridae) from the gills of wild and maricultured West Australian dhufish, Glaucosoma hebraicum (Teleostei: Glaucosomatidae), in Australia. J. Parasitol. 2001, 87, 749–754. [Google Scholar] [CrossRef]
- Kritsky, D.C.; Boeger, W.A. Neotropical Monogenoidea. 41: New and previously described species of Dactylogyridae (Platyhelminthes) from the gills of marine and freshwater perciform fishes (Teleostei) with proposal of a new genus and a hypothesis on phylogeny. Zoosystema 2002, 24, 7–40. [Google Scholar]
- Sun, Y.; Li, M.; Yang, T. Studies on Lethrinitrema Lim & Justine, 2011 (Monogenea: Dactylogyridae), with the description of two new species, a key to the genus and a phylogenetic analysis based on rDNA sequences. Syst. Parasitol. 2014, 88, 119–139. [Google Scholar]
- Al-Nabati, E.; Ali, S.; Al-Quraishy, S.; Alajmi, R.; Al-Shaebi, E.M.; Aljawdah, H.M.; Dkhil, M.A.; Abdel-Gaber, R. Heteromicrocotyla polyorchis Unnithan, 1961 (Monogenea: Heteromicrocotylidae), a gill parasite of the yellow-spotted trevally, Carangoides fulvoguttatus (Carangidae) from Saudi Arabia: Morphology and phylogeny. Microb. Pathog. 2021, 160, 105165. [Google Scholar] [CrossRef] [PubMed]
- Justine, J.L. Species of Pseudorhabdosynochus Yamaguti, 1958 (Monogenea: Diplectanidae) from Epinephelus fasciatus and E. merra (Perciformes: Serranidae) off New Caledonia and other parts of the Indo-Pacific Ocean, with a comparison of measurements of specimens prepared using different methods, and a description of P. caledonicus n. sp. Sys. Parasitol. 2005, 62, 1–37. [Google Scholar]
- Indaryanto, F.R.; Abdullah, M.F.; Wardiatno, Y.; Tiuria, R.; Imai, H. A Description of Lecithocladium angustiovum (Digenea: Hemiuridae) in Short Mackerel, Rastrelliger brachysoma (Scombridae), of Indonesia. Tropical Life Sci. Res. 2015, 26, 31–40. [Google Scholar]
- Li, L.; Zhao, W.T.; Guo, Y.N.; Zhang, L.P. Nematode parasites infection in the starry batfish Halieutaea stellata (vahl) (Lophiiformes: Ogcocephalidae) from the east and south China sea. J. Fish Dis. 2016, 39, 515–529. [Google Scholar] [CrossRef]
- Chaudhary, A.; Mukut, S.; Singh, H.S. Molecular characterization of three species belongs to the Allocreadioidea, Hemiuroidea and Plagiorchioidea (Platyhelminthes: Trematoda) infecting freshwater fishes in India. Helminthologia 2016, 53, 378–384. [Google Scholar] [CrossRef] [Green Version]
- Jovelin, R.; Justine, J.L. Phylogenetic relationships within the polyopisthocotylean monogeneans (Platyhelminthes) inferred from partial 28S rDNA sequences. Int. J. Parasitol. 2001, 32, 393–401. [Google Scholar] [CrossRef]
- Olson, P.D.; Littlewood, D.T.J. Phylogenetics of the Monogenea—Evidence from a medley of molecules. Int. J. Parasitol. 2002, 32, 233–244. [Google Scholar] [CrossRef]
- Šimková, A.; Plaisance, L.; Matějusová, I.; Morand, S.; Verneau, O. Phylogenetic relationships of the Dactylogyridae Bychowsky, 1933 (Monogenea: Dactylogyridea): The need for the systematic revision of the Ancyrocephalinae Bychowsky, 1937. Syst. Parasitol. 2003, 54, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Plaisance, L.; Littlewood, D.T.J.; Olson, P.D.; Morand, S. Molecular phylogeny of gill monogeneans (Platyhelminthes, Monogenea, Dactylogyridae) and colonization of Indo-West Pacific butterflyfish hosts (Perciformes, Chaetodontidae). Zool. Scri. 2005, 34, 425–436. [Google Scholar] [CrossRef]
- Strona, G.; Stefani, F.; Galli, P. Field preservation of monogenean parasites for molecular and morphological analyses. Parasitol. Int. 2009, 58, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Yoon, G.H.; Al-Jufaili, S.; Freeman, M.A.; Bron, J.E.; Paladini, G.; Shinn, A.P. Omanicotyleheterospina n. gen. et n. comb. (Monogenea: Microcotylidae) from the gills of Argyrops spinifer (Forsskål) (Teleostei: Sparidae) from the Sea of Oman. Parasites Vectors 2013, 6, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.W. Phylogenetic study on Microcotyle sp. (Monogenea) from common dentex (Dentex dentex) in the Mediterranean Sea, Greece. Afr. J. Biotechnol. 2015, 14, 2532–2538. [Google Scholar] [CrossRef] [Green Version]
- Tambireddy, N.; Gayatri, T.; Gireesh-Babu, P.; Pavan-Kumar, A. Molecular characterization and phylogeny of some mazocraeidean monogeneans from carangid fish. Acta Parasitol. 2016, 61, 360–368. [Google Scholar] [CrossRef]
- Dang, B.T.; Levsen, A.; Schander, C.; Bristow, G.A. Some Haliotrema (Monogenea: Dactylogyridae) from cultured grouper (Epinephelus spp.) with emphasis on the phylogenetic position of Haliotrema cromileptis. J. Parasitol. 2010, 96, 30–39. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Parasite Species | Family | Source | % Identity | GC Content |
|---|---|---|---|---|
| MG518632.1 Haliotrema susanae | Ancyrocephaliade | GenBank | 99.35 | 49.7 |
| EU541306.1 Haliotrema cromileptis | Ancyrocephaliade | GenBank | 87.28 | 49.4 |
| DQ537378.1 Haliotrema bihamulatum | Ancyrocephaliade | GenBank | 88.20 | 50.4 |
| EU836201.1 Haliotrema epinepheli | Ancyrocephaliade | GenBank | 85.37 | 47.3 |
| EU836207.1 Haliotrema macasarensis | Ancyrocephaliade | GenBank | 85.46 | 46 |
| DQ157664.1 Haliotrema johnstoni | Ancyrocephaliade | GenBank | 85.43 | 45.2 |
| DQ157662.1 Haliotrema platycephali | Ancyrocephaliade | GenBank | 85.43 | 45 |
| MG593838.1 Haliotrema magnihamus | Ancyrocephaliade | GenBank | 85.89 | 45.7 |
| MG593834.1 Haliotrema parvihamus | Ancyrocephaliade | GenBank | 85.89 | 46.7 |
| MG593837.1 Haliotrema bilobatus | Ancyrocephaliade | GenBank | 85.48 | 47.6 |
| MG593836.1 Haliotrema spinicirrus | Ancyrocephaliade | GenBank | 85.24 | 49.2 |
| MG593835.1 Haliotrema neobilobatus | Ancyrocephaliade | GenBank | 84.25 | 48.8 |
| EU836204.1 Haliotrema grossecurvitubus | Ancyrocephaliade | GenBank | 83.01 | 46.1 |
| DQ157661.1 Haliotrema fleti | Ancyrocephaliade | GenBank | 82.71 | 46.8 |
| EU836198.1 Haliotrema aurigae | Ancyrocephaliade | GenBank | 83.54 | 49.3 |
| EU836209.1 Haliotrema pratasensis | Ancyrocephaliade | GenBank | 83.37 | 48.8 |
| EU836208.1 Haliotrema macracantha | Ancyrocephaliade | GenBank | 83.37 | 50.2 |
| EU836206.1 Haliotrema leporinus | Ancyrocephaliade | GenBank | 83.29 | 49.7 |
| EU836199.1 Haliotrema ctenochaeti | Ancyrocephaliade | GenBank | 83.58 | 50.7 |
| MZ286653.1 Platycephalotrema bassense | Ancyrocephaliade | GenBank | 87.67 | 46.5 |
| MT023429.1 Pseudohaliotrema sphincteroporus | Ancyrocephaliade | GenBank | 86.65 | 51.3 |
| KJ571009.1 Bravohollisia parvianchoratus | Ancyrocephaliade | GenBank | 86.08 | 43.4 |
| MN176409.1 Glyphidohaptor safiensis | Ancyrocephaliade | GenBank | 86.26 | 49 |
| AF026114.1 Tetrancistrum sp. | Ancyrocephaliade | GenBank | 86.19 | 48.8 |
| KJ571017.1 Lethrinitrema zhanjiangense | Ancyrocephaliade | GenBank | 82.35 | 46 |
| MG586860.1 Euryhaliotrema fastigatum | Ancyrocephaliade | GenBank | 84.50 | 47 |
| AY820616.1 Euryhaliotrematoides grandis | Ancyrocephaliade | GenBank | 89.58 | 45.4 |
| KY305889.1 Sciadicleithrum meekii | Dactylogyridae | GenBank | 89.58 | 47.4 |
| MN567973.1 Dactylogyrus sp. | Dactylogyridae | GenBank | 88.41 | 51.7 |
| MG596661.1 Gussevia asota | Dactylogyridae | GenBank | 87.82 | 48.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Gaber, R.; Alghamdi, M.; Al Quraishy, S.; Al-Shaebi, E.M.; Elkhadragy, M.F.; El-Ashram, S.; Dkhil, M.A. A Monogenean Gill Parasite within the Genus Haliotrema (Ancyrocephalidae) Infecting Argyrops filamentosus Fish: Morphology and Molecular Studies. Animals 2023, 13, 1010. https://doi.org/10.3390/ani13061010
Abdel-Gaber R, Alghamdi M, Al Quraishy S, Al-Shaebi EM, Elkhadragy MF, El-Ashram S, Dkhil MA. A Monogenean Gill Parasite within the Genus Haliotrema (Ancyrocephalidae) Infecting Argyrops filamentosus Fish: Morphology and Molecular Studies. Animals. 2023; 13(6):1010. https://doi.org/10.3390/ani13061010
Chicago/Turabian StyleAbdel-Gaber, Rewaida, Masheil Alghamdi, Saleh Al Quraishy, Esam M. Al-Shaebi, Manal F. Elkhadragy, Saeed El-Ashram, and Mohamed A. Dkhil. 2023. "A Monogenean Gill Parasite within the Genus Haliotrema (Ancyrocephalidae) Infecting Argyrops filamentosus Fish: Morphology and Molecular Studies" Animals 13, no. 6: 1010. https://doi.org/10.3390/ani13061010