
Feeding Habits of Dentex maroccanus and the Effect of Body Size
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
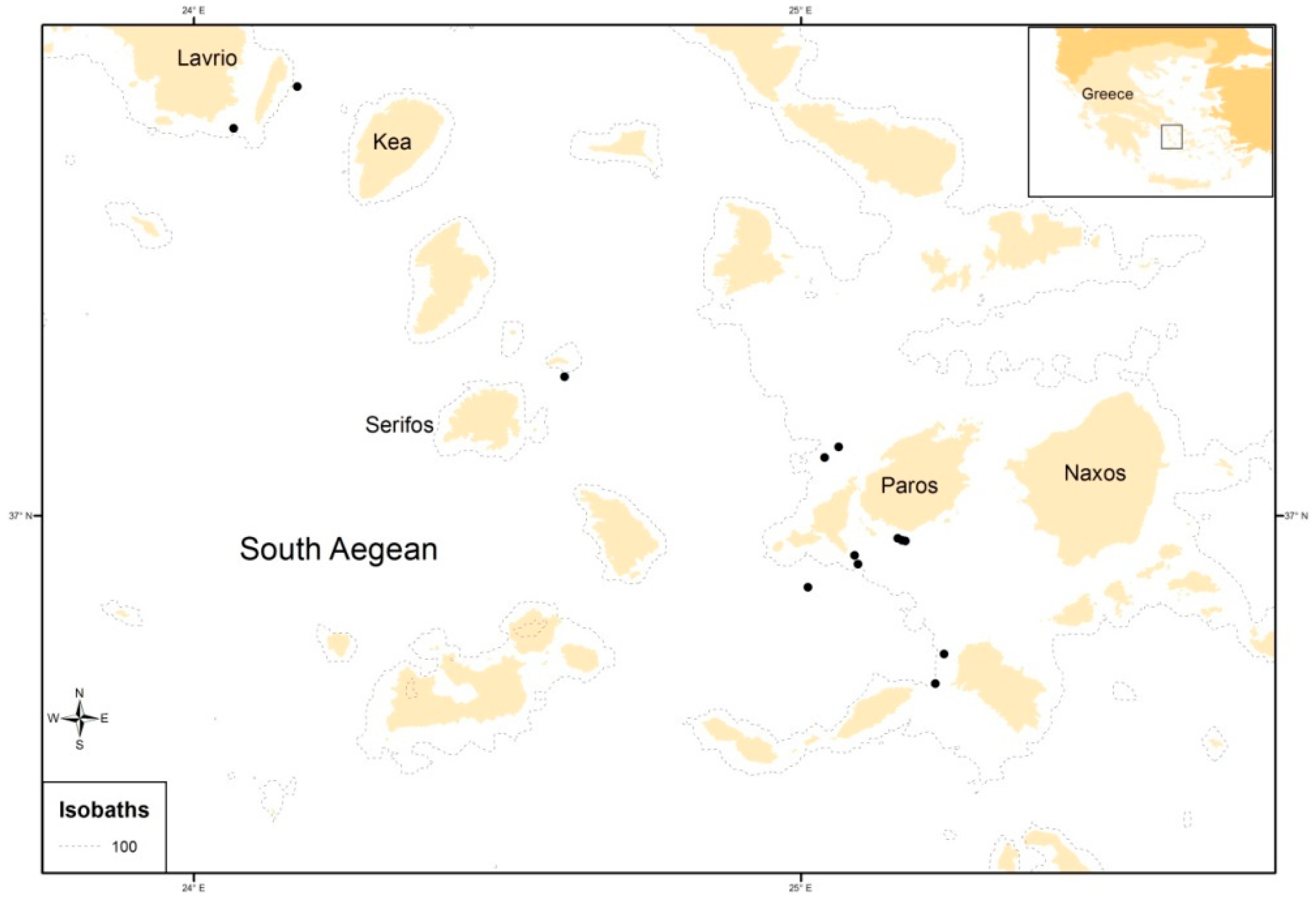
2. Materials and Methods
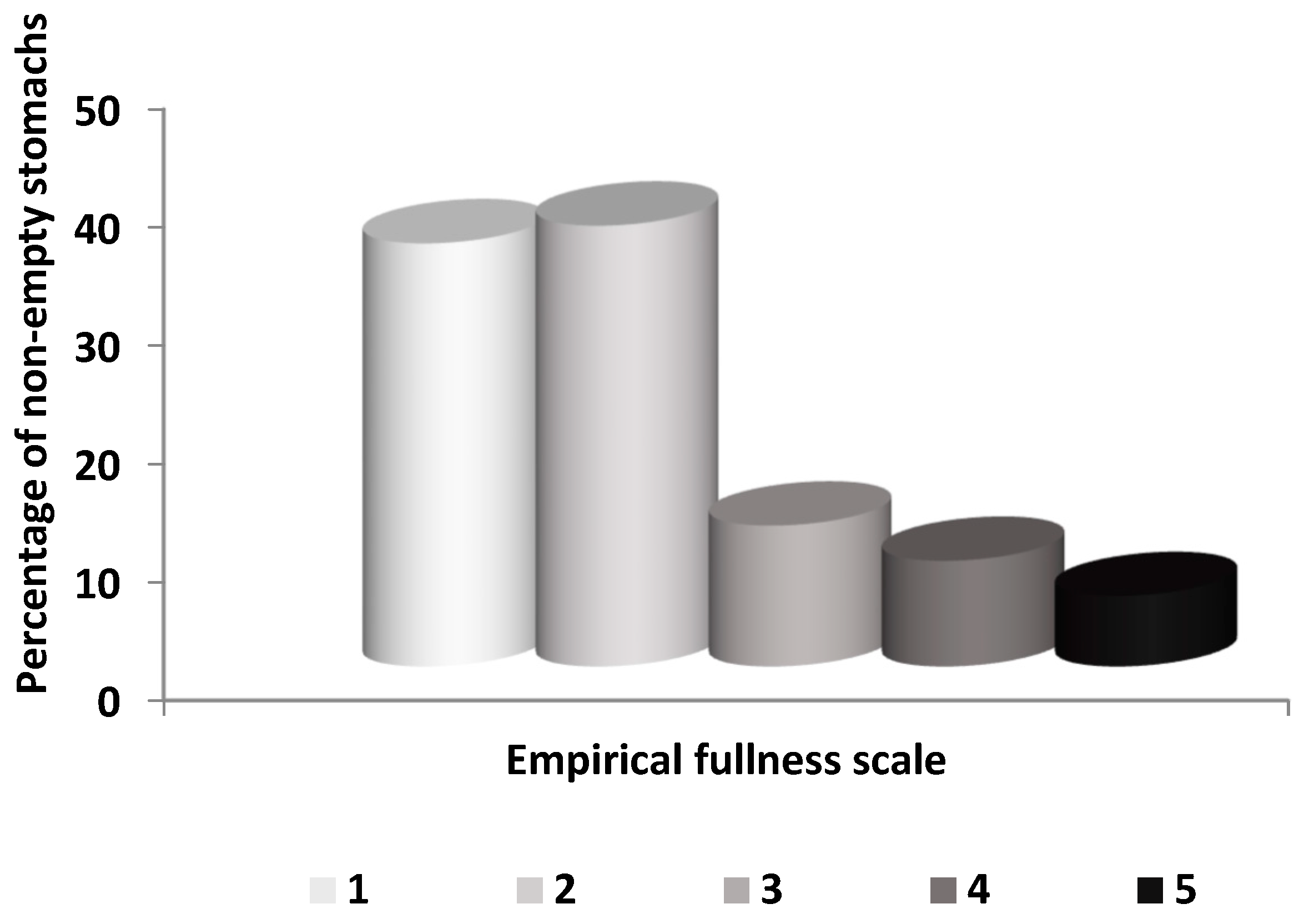
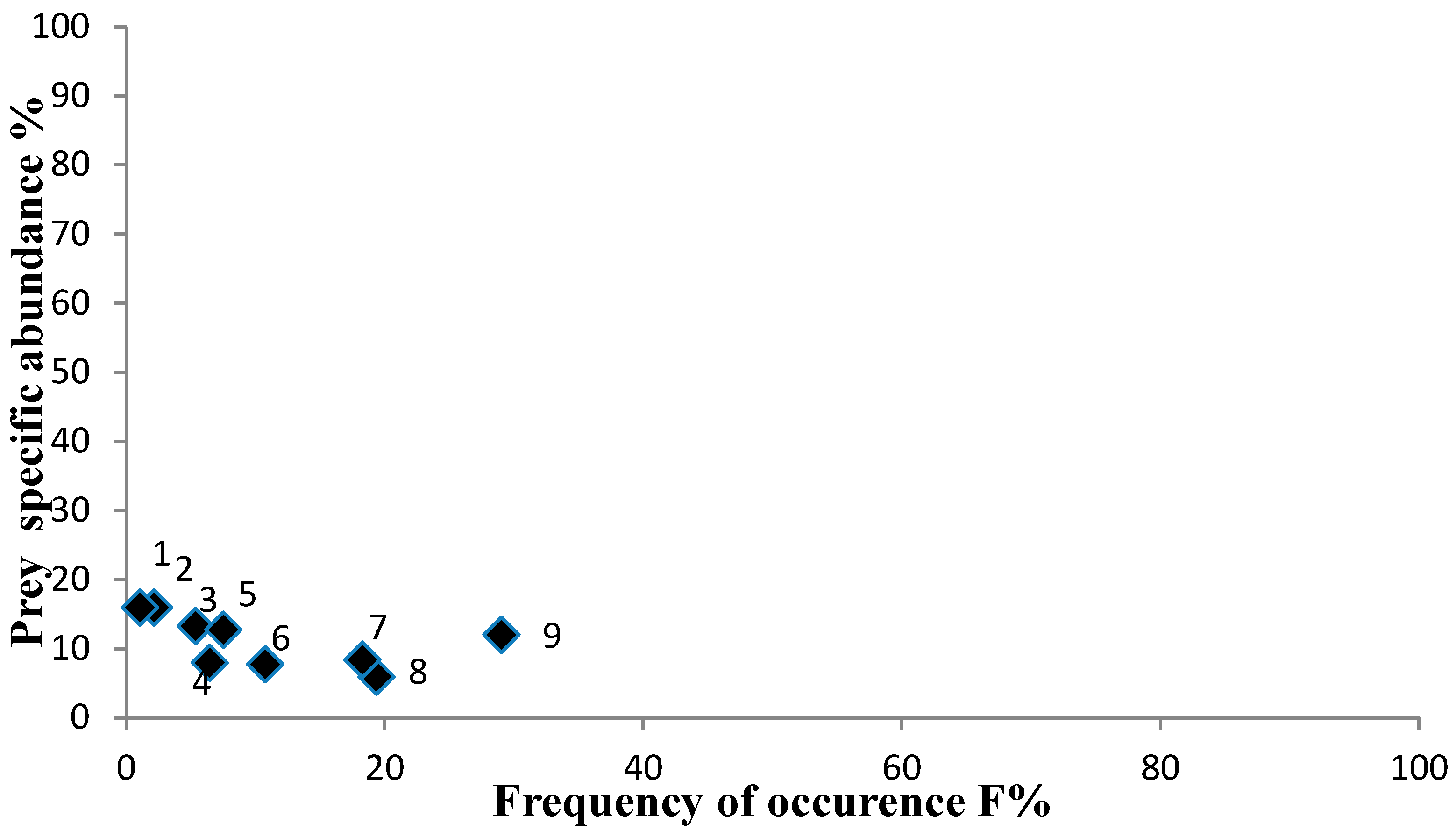
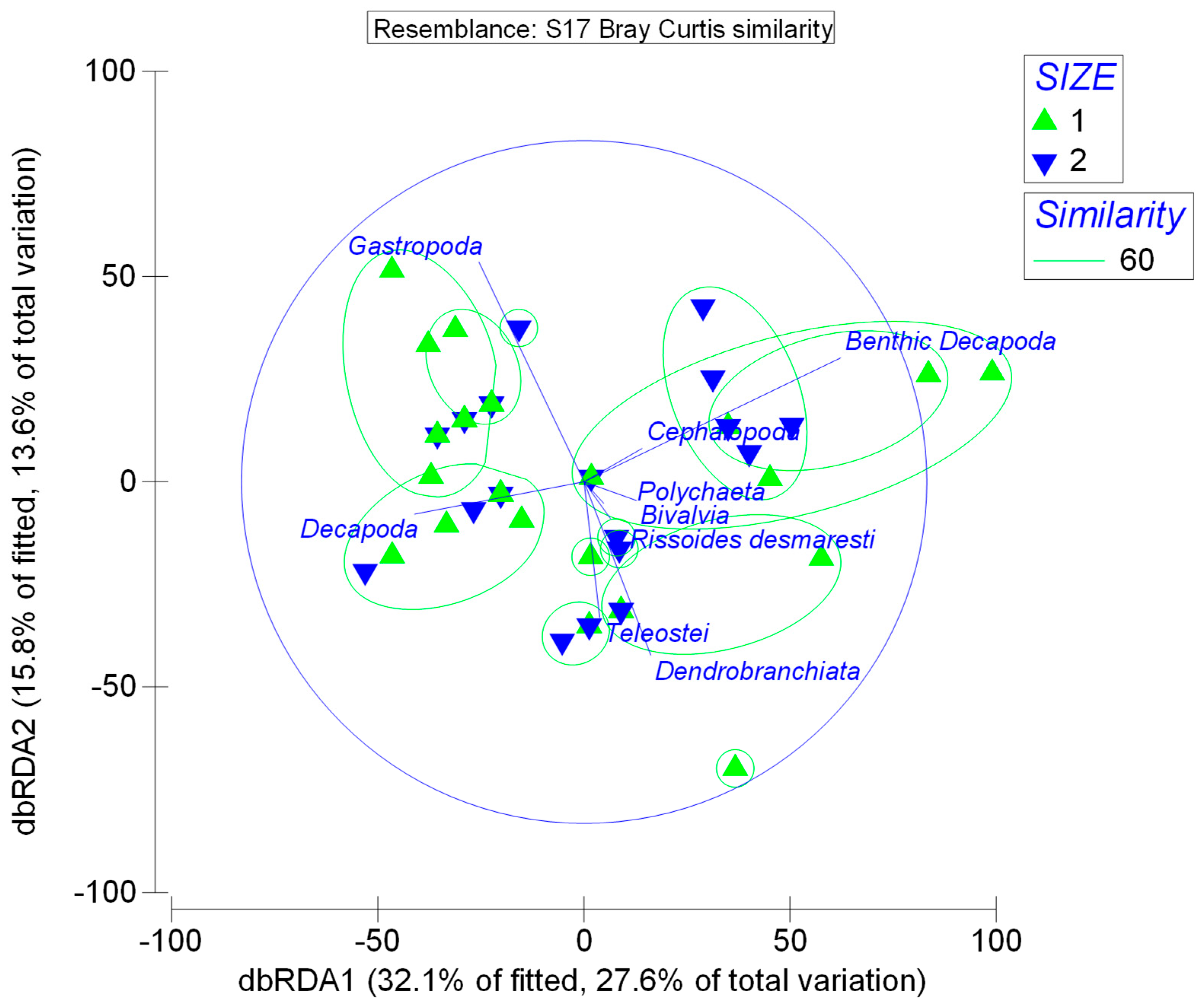
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baker, R.; Buckland, A.; Sheaves, M. Fish gut content analysis: Robust measures of diet composition. Fish Fish. 2014, 15, 170–177. [Google Scholar] [CrossRef]
- Sivadas, M.; Bhaskaran, M. Stomach content analysis of the Indian mackerel Rastrelliger kanagurta (Cuvier) from Calicut, Kerala. Indian J. Fish. 2009, 56, 143–146. [Google Scholar]
- Heuvel, C.E.; Haffner, G.D.; Zhao, Y.; Colborne, S.F.; Despenic, A.; Fisk, A.T. The influence of body size and season on the feeding ecology of three freshwater fishes with different diets in Lake Erie. J. Great Lakes Res. 2019, 45, 795–804. [Google Scholar] [CrossRef]
- Sánchez-Hernández, J.; Nunn, A.D.; Adams, C.E.; Amundsen, P.A. Causes and consequences of ontogenetic dietary shifts: A global synthesis using fish models. Biol. Rev. 2019, 94, 539–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauchot, M.L.; Hureau, J.C. Sparidae. In Fishes of the North-Eastern Atlantic and the Mediterranean (FNAM); Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E., Eds.; UNESCO: Paris, France, 1986; Volume 2, pp. 889–890. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. World Wide Web Electronic Publication. Available online: www.fishbase.org (accessed on 1 January 2023).
- Fischer, W.; Schneider, M.; Bauchot, M.L. Mediterranee et mer noire. In Fiches FAO d’ Identification des Espèces pour les Besoins de la Pêche; Zone De Pêche 37, Révision 1; Organisation des Nations Unies pour l’Alimentation et l’Agriculture: Rome, Italy, 1987; Volume 2, p. 1358. [Google Scholar]
- Maravelias, C.; Tsitsika, E.; Papaconstantinou, C. Evidence of Morocco dentex (Dentex maroccanus) distribution in the NE Mediterranean and relationships with environmental factors determined by Generalized Additive Modelling. Fish. Oceanogr. 2007, 16, 294–302. [Google Scholar] [CrossRef]
- Asimakopoulos, C.; Mytilineou, C. Distribution pattern of Dentex maroccanus in relation to environmental factors in the Aegean Sea. In Proceedings of the 4th International Congress on Applied Ichthyology, Oceanography & Aquatic Environment (HydroMediT 2021), Virtual, 4–6 November 2021; pp. 534–535. [Google Scholar]
- Hemida, F.; Ghazli, R. Contribution À L’écologie De Dentex maroccanus (Valenciennes, 1830) Des Côtes Algériennes: Répartition Géographique, Bathymétrique Et En Fonction Du Substrat; Rapport de la Commission Internationale de l’Exploration de la mer Méditerranée (CIESM): Marseille, France, 1998; Volume 35, pp. 448–449. [Google Scholar]
- Papaconstantinou, C.; Petrakis, G.; Caragitsou, E.; Vasilopoulou, V.; Mytilineou, C. Investigation of the Abundance and Distribution of the Demersal Stocks of Primary Importance to the Greek Fishery in the Southern Aegean Sea. In National Centre for Marine Research Technical Report; NCMR Publications: Athens, Greece, 1998; p. 312. [Google Scholar]
- Chemmam-Abdelkader, B.; Kraiem, M.M.; El Abed, A. Etude de l’age et de la croissance de deux especes de dentes (Dentex dentex et Dentex maroccanus) des cotes Tunisiennes. Bull. L’institut Natl. Des. Sci. Technol. Mer Salammbô 2004, 31, 43–51. [Google Scholar]
- Karakulak, F.S.; Erk, H.; Bilgin, B. Length-weight relationships for 47 coastal fish species from the northern Aegean Sea, Turkey. J. Appl. Ichthyol. 2006, 22, 274–278. [Google Scholar] [CrossRef]
- Ismen, A.; Özen, O.; Altinağaç, U.; Özekinci, U.; Ayaz, A. Weight-length relationships of 63 fish species in Saros Bay, Turkey. J. Appl. Ichthyol. 2007, 23, 707–708. [Google Scholar] [CrossRef]
- Ceyhan, T.; Akyol, O.; Erdem, M. Length-weight relationships of fishes from Gökova Bay, Turkey (Aegean Sea). Turk. J. Zool. 2009, 33, 69–72. [Google Scholar] [CrossRef]
- Mohdeb, R.; Kara, M.H. Age, growth and reproduction of the Morocco dentex Dentex maroccanus of the eastern coast of Algeria. J. Mar. Biol. Assoc. U. K. 2014, 95, 1261–1270. [Google Scholar] [CrossRef]
- Gul, G.; Ismen, A.; Arslan, M. Age, growth, and reproduction of Dentex maroccanus (Actinopterygii: Perciformes: Sparidae) in the Saros Bay (north Aegean Sea). Acta Ichthyol. Piscat. 2014, 44, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Bayhan, B.; Heral, O.; Taskavak, E.; Topkara, E.T.; Kara, A. Morphometric characteristics of the Morocco dentex Dentex dentex, Valenciennes, 1830, in the Izmir Bay. Rapp. Comm. Int. Mer Méditerranée 2016, 41, 335. [Google Scholar]
- Evangelopoulos, A.; Batjakas, I.; Koutsoubas, D. Length–weight relationships of 9 commercial fish species from the North Aegean Sea. Acta Adriat. 2017, 58, 187–192. [Google Scholar] [CrossRef]
- Taylan, B.; Bayhan, B.; Heral, O. Fecundity of Morocco Dentex Dentex maroccanus Valenciennes, 1830 distributed in Izmir Bay (Central Aegean Sea of Turkey). Turk. J. Agric. Food Sci. Technol. 2018, 6, 624. [Google Scholar] [CrossRef] [Green Version]
- Heral, O.; Bayhan, B. Age and growth of Morocco Dentex Dentex maroccanus Valenciennes, 1830 (Actinopterygii: Sparidae) in Izmir Bay, Central Aegean Sea, Turkey. Acta Zool. Bulg. 2020, 72, 149–154. [Google Scholar]
- Bilecenoğlu, M.; Yokeş, M.B. New additions to the fish fauna of Sea of Marmara: Dentex maroccanus Valenciennes, 1830 (Sparidae) and Gobius couchi Miller & El-Tawil, 1974 (Gobiidae). FishTaxa 2022, 24, 42–48. [Google Scholar]
- Lamrini, A.; Bouymajjane, A. Biologie de Dentex maroccanus (Valenciennes, 1830) dans la région de Safi. Actes Inst. Agron. Vét. 2002, 22, 11–18. [Google Scholar]
- Bayhan, B.; Sever, T.; Heral, O. Diet composition of the Morocco dentex: Dentex maroccanus Valenciennes, 1830 (Teleostei: Sparidae) in the central Turkish Aegean Sea. Oceanol. Hydrobiol. Stud. 2017, 46, 133–139. [Google Scholar] [CrossRef]
- Mohdeb, R.; Derbal, F.; Kara, M.H. Diet composition and variations of Morocco dentex Dentex maroccanus (Sparidae) from Eastern Algeria. Cybium Int. J. Ichthyol. 2017, 41, 317–324. [Google Scholar] [CrossRef]
- Hyslop, E.J. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef] [Green Version]
- Hureau, J.C. Biologie comparée de quelques poisons antarctiques (Nototheniidae). Bull. L’inst. Océanograph. Monaco 1970, 68, 139–164. [Google Scholar]
- Pinkas, L.; Oliphant, M.S.; Iverson, I.L.K. Food habits of albacore, blue fin tuna and bonito in California waters. Fish. Bull. 1971, 152, 1–105. [Google Scholar]
- Hacunda, J.S. Trophic relationships among demersal fishes in a coastal area of the Gulf of Maine. Fish. Bull. 1981, 79, 775–788. [Google Scholar]
- Amundsen, P.A.; Gabler, H.M.; Staldvik, F.J. A new approach to graphical analysis of feeding strategy from stomach contents data—Modification of the Costello (1990) method. J. Fish Biol. 1996, 48, 607–614. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E: Plymouth, UK, 2014. [Google Scholar]
- Hulbert, S.H. The measurement of niche overlap and some relatives. Ecology 1978, 59, 67–77. [Google Scholar] [CrossRef]
- Novakowski, G.C.; Hahn, N.S.; Fugi, R. Diet seasonality and food overlap of the fish assemblage in a pantanal pond. Neotrop. Ichthyol. 2008, 6, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Pauly, D.; Christensen, V. Primary production required to sustain global fisheries. Nature 1995, 374, 255–257. [Google Scholar] [CrossRef]
- Pauly, D.; Trites, A.W.; Capuli, E.; Christensen, V. Diet composition and trophic levels of marine mammals. ICES J. Mar. Sci. 1998, 55, 467–481. [Google Scholar] [CrossRef]
- Pauly, D.; Sa-a, P. Estimating trophic levels from individual food items. In FishBase 2000: Concepts, Design and Data Sources; Froese, R., Pauly, D., Eds.; ICLARM: Los Baños, Philippines, 2000; p. 185. [Google Scholar]
- Dias, T.S.; Stein, R.J.; Fialho, C.B. Ontogenetic variations and feeding habits of a Neotropical annual fish from southern Brazil. Iheringia-Sér. Zool. 2017, 107, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Nikolsky, G.V. The Ecology of Fishes; Academic Press: London, UK, 1976; p. 352. [Google Scholar]
- Stergiou, K.I.; Karpouzi, V.S. Feeding habits and trophic levels of Mediterranean fish. Rev. Fish Biol. Fish. 2002, 11, 217–254. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
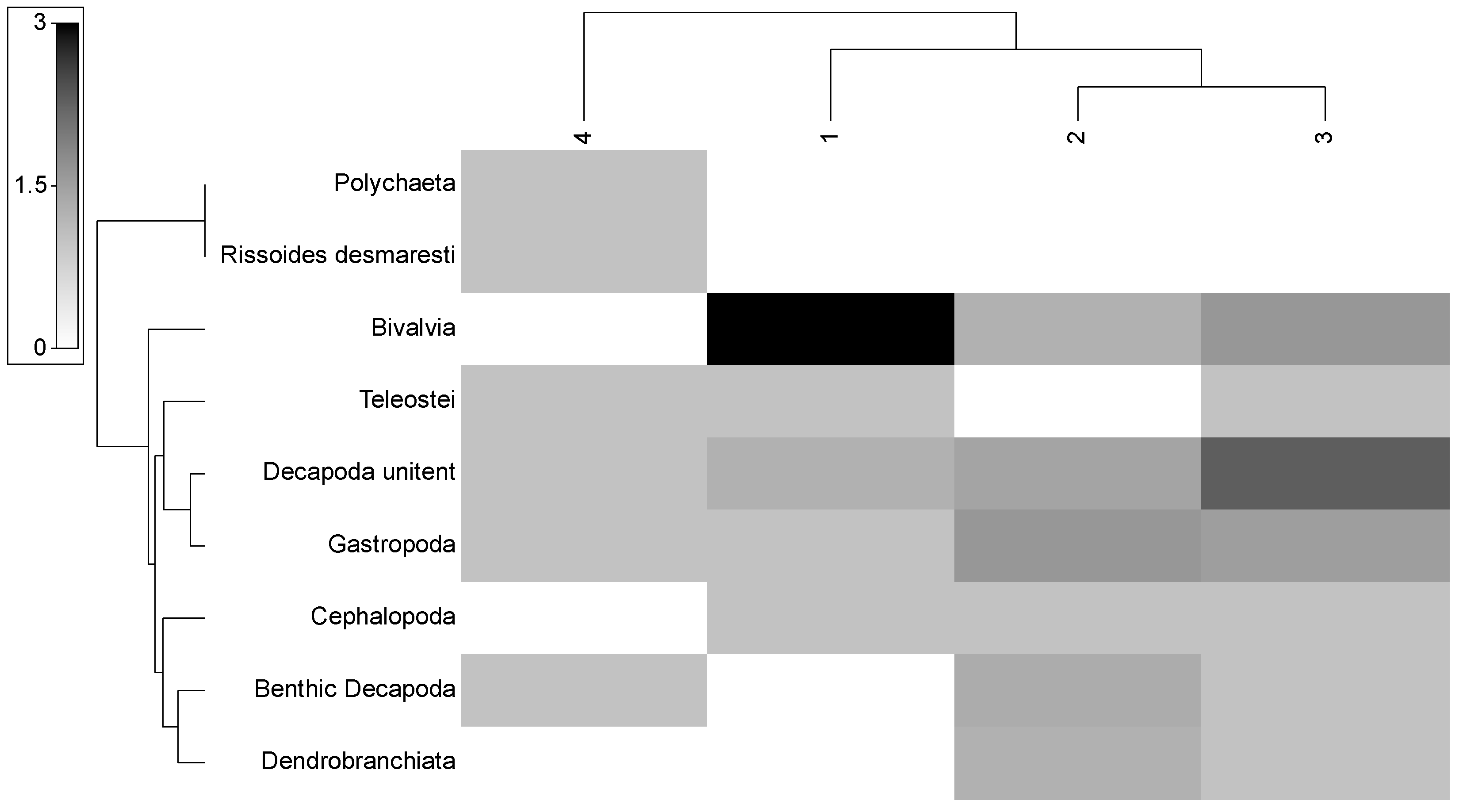
| Prey Taxa | %N | %F | %W | Q | %IRI |
|---|---|---|---|---|---|
| ANNELIDA | |||||
| Polychaeta | 0.8 | 1.06 | 0.30 | 0.24 | 0.04 |
| MOLLUSCA | |||||
| Gastropoda | 20.00 | 19.15 | 5.34 | 106.80 | 8.18 |
| Gastropoda unident. | 12.80 | 12.77 | 4.27 | 54.64 | 7.69 |
| Chilodontaidae | 0.80 | 1.06 | 0.21 | 0.17 | 0.04 |
| Fasciolariidae | 2.40 | 1.06 | 0.12 | 0.29 | 0.09 |
| Naticidae | 1.60 | 2.13 | 0.35 | 0.56 | 0.15 |
| Pyramidellidae | 2.40 | 2.13 | 0.39 | 0.94 | 0.21 |
| Bivalvia | 12.80 | 11.70 | 2.82 | 36.10 | 2.66 |
| Bivalvia unident. | 4.80 | 5.32 | 1.35 | 6.50 | 1.15 |
| Nuculanidae | 0.80 | 1.06 | 0.91 | 0.73 | 0.06 |
| Pectinidae | 7.20 | 5.32 | 0.55 | 3.95 | 1.45 |
| Cephalopoda | 4.80 | 6.38 | 18.89 | 90.67 | 5.34 |
| ARTHROPODA-Crustacea | |||||
| Decapoda unident. | 34.40 | 28.72 | 34.98 | 1203.30 | 70.32 |
| Benthic Decapoda | 15.20 | 18.09 | 10.08 | 153.22 | 6.54 |
| Brachyura | 8.80 | 10.64 | 4.13 | 36.34 | 4.85 |
| Alpheidae | 0.80 | 1.06 | 1.37 | 1.10 | 0.08 |
| Munididae | 0.80 | 1.06 | 1.22 | 0.97 | 0.08 |
| Paguridae | 4.80 | 5.32 | 3.36 | 16.12 | 1.53 |
| Dendrobranchiata | 6.40 | 7.45 | 6.65 | 42.58 | 3.43 |
| (pelagic decapoda) | |||||
| Stomatopoda | |||||
| Rissoides desmaresti | 1.6 | 2.13 | 11.70 | 18.72 | 1.00 |
| CHORDATA-Teleostei | 4.00 | 5.32 | 9.23 | 36.93 | 2.48 |
| Length range (mm) | 99–186 | ||||
| N of individuals | 67 |
| PERMANOVA | Abundance p-Value |
|---|---|
| Size | 0.012 |
| Sex | 0.470 |
| Season | 0.146 |
| Sex × Size | 0.511 |
| Sex × Season | 0.474 |
| Size × Season | 0.140 |
| Sex × Size × Season | 0.516 |
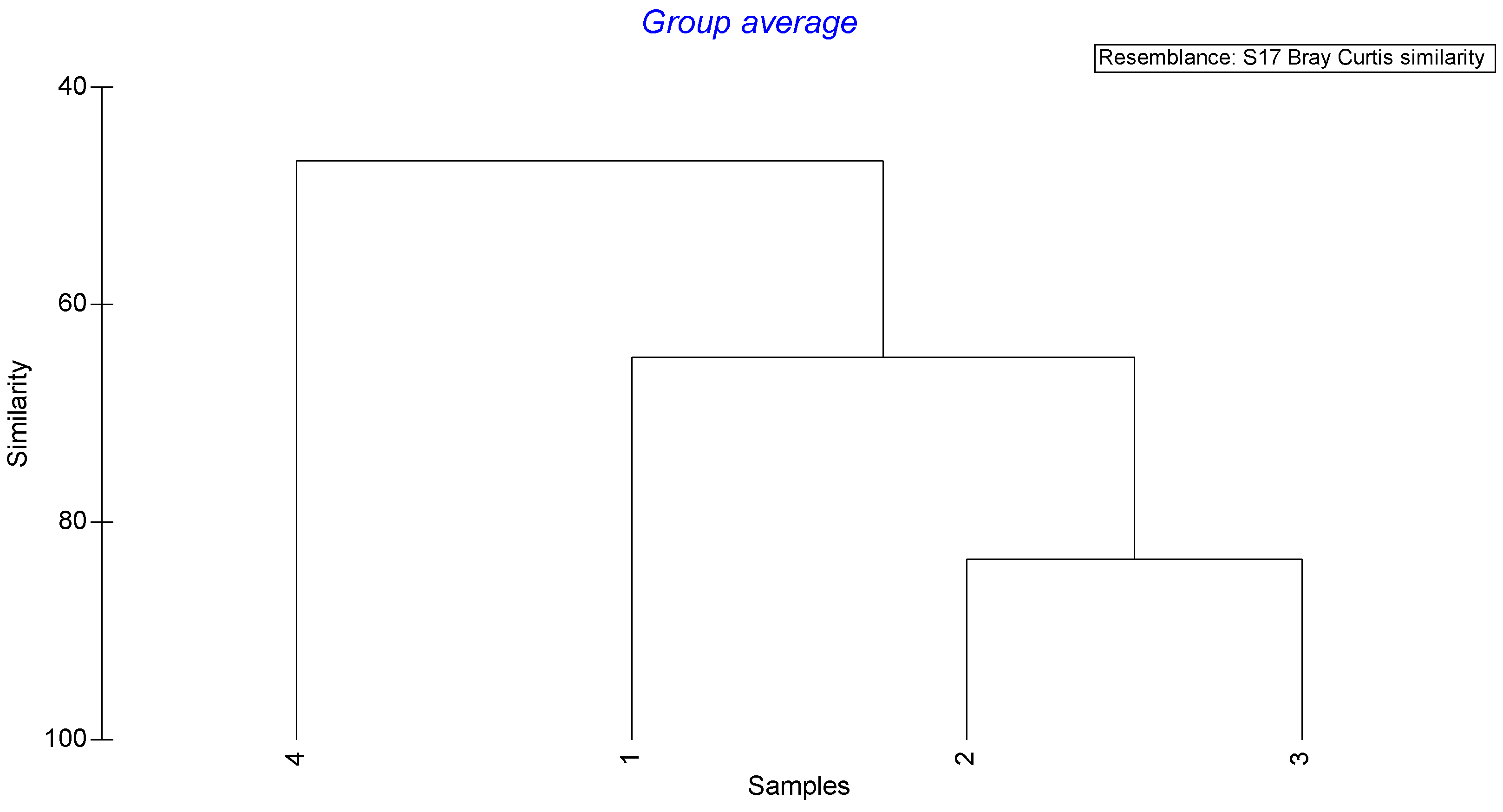
| Size Groups | Average Dissimilarity (%) | Prey Taxa | Contrib. (%) | Diet Overlap |
|---|---|---|---|---|
| 1–2 | 40.55 | Bivalvia | 28.51 | 0.55 |
| 1–3 | 29.67 | Bivalvia | 28.36 | 0.65 |
| 2–3 | 16.58 | Teleostei | 34.91 | 0.65 |
| 1–4 | 54.72 | Bivalvia | 41.38 | 0.34 |
| 2–4 | 56.80 | Bivalvia and Dendrobranchiata | 15.85 | 0.26 |
| 3–4 | 48.00 | Bivalvia | 21.66 | 0.32 |
| Size Group | H′ | B | TrL |
|---|---|---|---|
| 1 | 0.10 ± 0.09 | 0.35 | 3.70 ± 0.47 |
| 2 | 0.32 ± 0.08 | 0.46 | 3.70 ± 0.43 |
| 3 | 0.31 ± 0.07 | 0.50 | 4.10 ± 0.54 |
| 4 | 0.12 ± 0.14 | 0.90 | 4.00 ± 0.62 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mina, A.; Mytilineou, C.; Kaminas, A.; Rekleiti, A.; Siapatis, A.; Anastasopoulou, A. Feeding Habits of Dentex maroccanus and the Effect of Body Size. Animals 2023, 13, 939. https://doi.org/10.3390/ani13050939
Mina A, Mytilineou C, Kaminas A, Rekleiti A, Siapatis A, Anastasopoulou A. Feeding Habits of Dentex maroccanus and the Effect of Body Size. Animals. 2023; 13(5):939. https://doi.org/10.3390/ani13050939
Chicago/Turabian StyleMina, Amalia, Chryssi Mytilineou, Alexandros Kaminas, Anthi Rekleiti, Apostolos Siapatis, and Aikaterini Anastasopoulou. 2023. "Feeding Habits of Dentex maroccanus and the Effect of Body Size" Animals 13, no. 5: 939. https://doi.org/10.3390/ani13050939