
Metabolomic Analysis of Multiple Biological Specimens (Feces, Serum, and Urine) by 1H-NMR Spectroscopy from Dairy Cows with Clinical Mastitis
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Metabolomic Analysis
2.3. Statistical Analysis
3. Results
3.1. Characterization of Molecules in Feces, Serum, and Urine
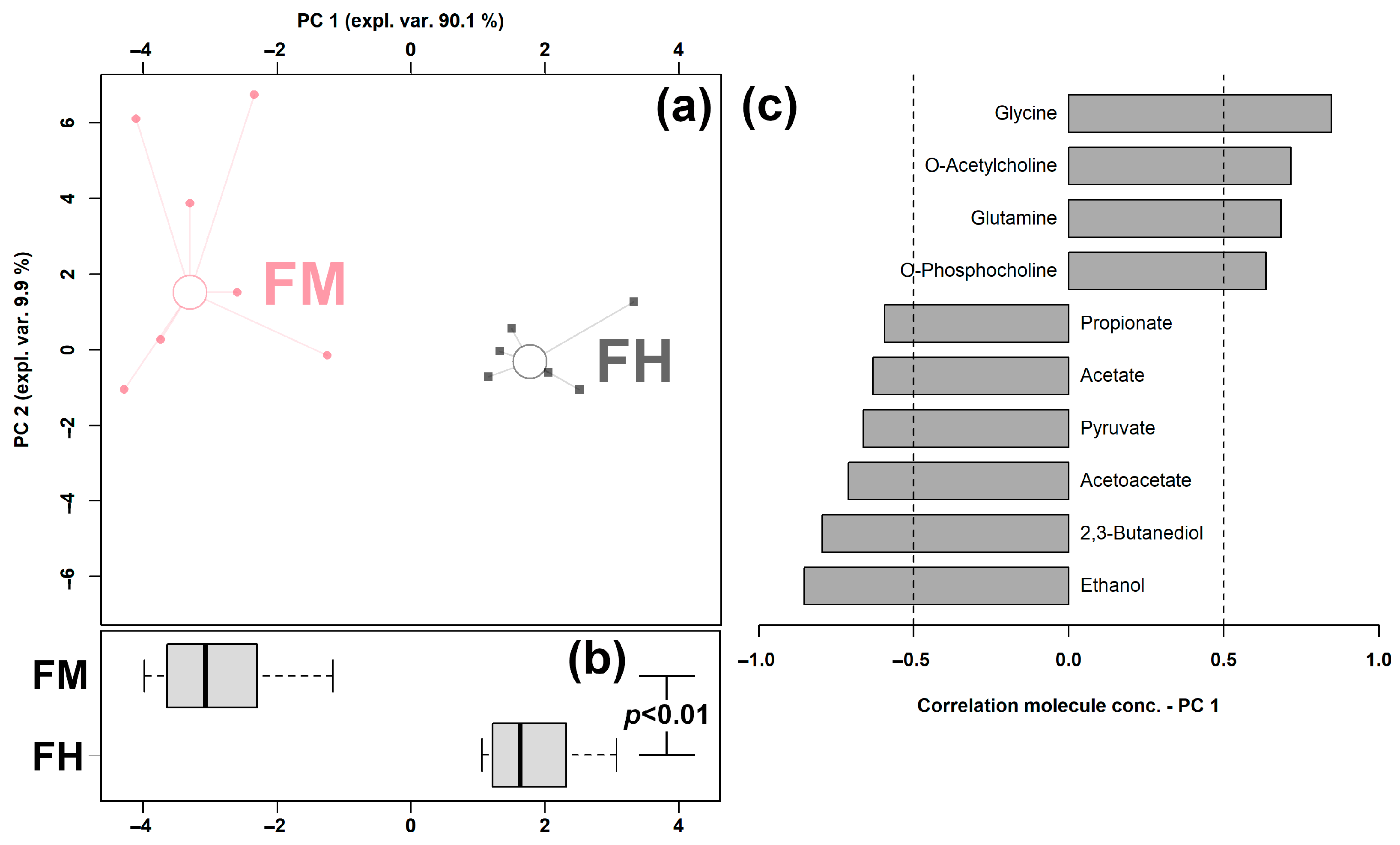
3.2. Feces Metabolome Affected by Clinical Mastitis
3.3. Serum Metabolomic Features Affected by Clinical Mastitis
3.4. Urine Metabolome Affected by Clinical Mastitis
3.5. Pathway Analysis in Relation to Clinical Mastitis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ezzat Alnakip, M.; Quintela-Baluja, M.; Böhme, K.; Fernández-No, I.; Caamaño-Antelo, S.; Calo-Mata, P.; Barros-Velázquez, J. The Immunology of Mammary Gland of Dairy Ruminants between Healthy and Inflammatory Conditions. J. Vet. Med. 2014, 2014, 659801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamari, I.; Mimoune, N.; Khelef, D. Effect of Feed Additive Supplementation on Bovine Subclinical Mastitis. Vet. Stanica 2021, 52, 445–460. [Google Scholar] [CrossRef]
- Mimoune, N.; Saidi, R.; Benadjel, O.; Khelef, D.; Kaidi, R. Alternative Treatment of Bovine Mastitis. Vet. Stanica 2021, 52, 639–649. [Google Scholar] [CrossRef]
- Nyman, A.K.; Persson Waller, K.; Bennedsgaard, T.W.; Larsen, T.; Emanuelson, U. Associations of Udder-Health Indicators with Cow Factors and with Intramammary Infection in Dairy Cows. J. Dairy Sci. 2014, 97, 5459–5473. [Google Scholar] [CrossRef] [Green Version]
- Cvetnić, L.; Špičić, S.; Kompes, G.; Habrun, B.; Katalinić-Janković, V.; Cvetnić, M.; Zdelar-Tuk, M.; Reil, I.; Duvnjak, S.; Cvetnić, Ž.; et al. Bovine Mastitis Caused by Rapid-Growth Environmental Mycobacteria. Vet. Stanica 2022, 53, 493–501. [Google Scholar] [CrossRef]
- Saidi, R.; Cantekin, Z.; Mimoune, N.; Ergun, Y.; Solmaz, H.; Khelef, D.; Kaidi, R. Investigation of the Presence of Slime Production, VanA Gene and Antiseptic Resistance Genes in Staphylococci Isolated from Bovine Mastitis in Algeria. Vet. Stanica 2021, 52, 57–63. [Google Scholar] [CrossRef]
- Pavliček, D.; Lugomer, M.D.; Novosel, T. Halogenated Salicylanilides—Flukicide Drugs. Vet. Stanica 2022, 53, 723–734. [Google Scholar] [CrossRef]
- Cvetnić, L.; Samardžija, M.; Duvnjak, S.; Habrun, B.; Cvetnić, M.; Tkalec, V.J.; Đuričić, D.; Benić, M. Multi Locus Sequence Typing and Spa Typing of Staphylococcus Aureus Isolated from the Milk of Cows with Subclinical Mastitis in Croatia. Microorganisms 2021, 9, 725. [Google Scholar] [CrossRef]
- Luangwilai, M.; Duangmal, K.; Chantaprasarn, N.; Settachaimongkon, S. Comparative Metabolite Profiling of Raw Milk from Subclinical and Clinical Mastitis Cows Using 1H-NMR Combined with Chemometric Analysis. Int. J. Food Sci. Technol. 2021, 56, 493–503. [Google Scholar] [CrossRef]
- Sargeant, J.M.; Leslie, K.E.; Shirley, J.E.; Pulkrabek, B.J.; Lim, G.H. Sensitivity and Specificity of Somatic Cell Count and California Mastitis Test for Identifying Intramammary Infection in Early Lactation. J. Dairy Sci. 2001, 84, 2018–2024. [Google Scholar] [CrossRef]
- Kneževic, K.; Dobranic, V.; Samardžija, M.; Getz, I.; Efendic, M.; Šavoric, J.; Butkovic, I.; Cvetnic, M.; Macešic, N.; Duricic, D.; et al. Use of Somatic Cell Count in the Diagnosis of Mastitis and Its Impacts on Milk Quality. Vet. Stanica 2021, 52, 751–764. [Google Scholar]
- Zhang, J.; Li, W.; Tang, Y.; Liu, X.; Zhang, H.; Zhou, Y.; Wang, Y.; Xiao, W.; Yu, Y. Testing Two Somatic Cell Count Cutoff Values for Bovine Subclinical Mastitis Detection Based on Milk Microbiota and Peripheral Blood Leukocyte Transcriptome Profile. Animals 2022, 12, 1694. [Google Scholar] [CrossRef]
- Laghi, L.; Picone, G.; Capozzi, F. Nuclear Magnetic Resonance for Foodomics beyond Food Analysis. TrAC Trends Anal. Chem. 2014, 59, 93–102. [Google Scholar] [CrossRef]
- Wishart, D.S. NMR Metabolomics: A Look Ahead. J. Magn. Reson. 2019, 306, 155–161. [Google Scholar] [CrossRef]
- Zhu, C.; Faillace, V.; Laus, F.; Bazzano, M.; Laghi, L. Characterization of Trotter Horses Urine Metabolome by Means of Proton Nuclear Magnetic Resonance Spectroscopy. Metabolomics 2018, 14, 106. [Google Scholar] [CrossRef]
- Zampiga, M.; Laghi, L.; Petracci, M.; Zhu, C.; Meluzzi, A.; Dridi, S.; Sirri, F. Effect of Dietary Arginine to Lysine Ratios on Productive Performance, Meat Quality, Plasma and Muscle Metabolomics Profile in Fast-Growing Broiler Chickens. J. Anim. Sci. Biotechnol. 2018, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Laghi, L.; Zhang, Z.; He, Y.; Wu, D.; Zhang, H.; Huang, Y.; Li, C.; Zou, L. First Steps toward the Giant Panda Metabolome Database: Untargeted Metabolomics of Feces, Urine, Serum, and Saliva by 1H NMR. J. Proteome Res. 2020, 19, 1052–1059. [Google Scholar] [CrossRef]
- Zhu, C.; Li, C.; Wang, Y.; Laghi, L. Characterization of Yak Common Biofluids Metabolome by Means of Proton Nuclear Magnetic Resonance Spectroscopy. Metabolites 2019, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Bazzano, M.; Laghi, L.; Zhu, C.; Magi, G.E.; Serri, E.; Spaterna, A.; Tesei, B.; Laus, F. Metabolomics of Tracheal Wash Samples and Exhaled Breath Condensates in Healthy Horses and Horses Affected by Equine Asthma. J. Breath Res. 2018, 12, 46015. [Google Scholar] [CrossRef]
- Zhu, C.; Jin, L.; Luo, B.; Zhou, Q.; Dong, L.; Li, X.; Zhang, H.; Huang, Y.; Li, C.; Zou, L.; et al. Dominant Components of the Giant Panda Seminal Plasma Metabolome, Characterized By1 H-NMR Spectroscopy. Animals 2022, 12, 1536. [Google Scholar] [CrossRef]
- Bobbo, T.; Meoni, G.; Niero, G.; Tenori, L.; Luchinat, C.; Cassandro, M.; Penasa, M. Nuclear Magnetic Resonance Spectroscopy to Investigate the Association between Milk Metabolites and Udder Quarter Health Status in Dairy Cows. J. Dairy Sci. 2022, 105, 535–548. [Google Scholar] [CrossRef]
- De Paepe, E.; van Meulebroek, L.; Rombouts, C.; Huysman, S.; Verplanken, K.; Lapauw, B.; Wauters, J.; Hemeryck, L.Y.; Vanhaecke, L. A Validated Multi-Matrix Platform for Metabolomic Fingerprinting of Human Urine, Feces and Plasma Using Ultra-High Performance Liquid-Chromatography Coupled to Hybrid Orbitrap High-Resolution Mass Spectrometry. Anal. Chim. Acta 2018, 1033, 108–118. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, A.; Han, Y.; Wang, P.; Sun, H.; Song, G.; Dong, T.; Yuan, Y.; Yuan, X.; Zhang, M.; et al. Urine Metabolomics Analysis for Biomarker Discovery and Detection of Jaundice Syndrome in Patients with Liver Disease. Mol. Cell. Proteom. 2012, 11, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Sun, Z.; Zeng, B.; Huang, S.; Zhao, J.; Zhang, Y.; Su, X.; Xu, J.; Wei, H.; Zhang, H. Cow-to-Mouse Fecal Transplantations Suggest Intestinal Microbiome as One Cause of Mastitis. Microbiome 2018, 6, 200. [Google Scholar] [CrossRef]
- Hu, X.; Guo, J.; Zhao, C.; Jiang, P.; Maimai, T.; Yanyi, L.; Cao, Y.; Fu, Y.; Zhang, N. The Gut Microbiota Contributes to the Development of Staphylococcus Aureus-Induced Mastitis in Mice. ISME J. 2020, 14, 1897–1910. [Google Scholar] [CrossRef]
- Zhu, C.; Petracci, M.; Li, C.; Fiore, E.; Laghi, L. An Untargeted Metabolomics Investigation of Jiulong Yak (Bos Grunniens) Meat by 1H-NMR. Foods 2020, 9, 481. [Google Scholar] [CrossRef]
- Laghi, L.; Zhu, C.; Campagna, G.; Rossi, G.; Bazzano, M.; Laus, F. Probiotic Supplementation in Trained Trotter Horses: Effect on Blood Clinical Pathology Data and Urine Metabolomic Assessed in Field. J. Appl. Physiol. 2018, 125, 654–660. [Google Scholar] [CrossRef]
- Kneen, M.A.; Annegarn, H.J. Algorithm for Fitting XRF, SEM and PIXE X-Ray Spectra Backgrounds. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 1996, 109–110, 209–213. [Google Scholar] [CrossRef]
- Liland, K.H.; Almøy, T.; Mevik, B.H. Optimal Choice of Baseline Correction for Multivariate Calibration of Spectra. Appl. Spectrosc. 2010, 64, 1007–1016. [Google Scholar] [CrossRef]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic Quotient Normalization as Robust Method to Account for Dilution of Complex Biological Mixtures. Application In1H NMR Metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef]
- RR Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; Volume 1, ISBN 3900051070. [Google Scholar]
- Box, G.E.P.; Cox, D.R. An Analysis of Transformations. J. R. Stat. Soc. Ser. B (Methodol.) 2018, 26, 211–243. [Google Scholar] [CrossRef]
- Hubert, M.; Rousseeuw, P.J.; Vanden Branden, K. ROBPCA: A New Approach to Robust Principal Component Analysis. Technometrics 2005, 47, 64–79. [Google Scholar] [CrossRef]
- Li, C.; Soufan, O.; Chong, J.; Xia, J.; Bourque, G.; Li, S.; Caraus, I.; Wishart, D.S. MetaboAnalyst 4.0: Towards More Transparent and Integrative Metabolomics Analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Jiang, H.; He, Q.; Wang, M.; Xue, J.; Liu, H.; Shi, K.; Wei, M.; Liang, S.; Zhang, L. Liquid Chromatography/Mass Spectrometry Reveals the Effect of Lactobacillus Treatment on the Faecal Metabolite Profile of Rats with Chronic Renal Failure. Nephron 2017, 135, 156–166. [Google Scholar] [CrossRef]
- Zhu, C.; Yang, Z.; He, L.; Lu, X.; Tang, J.; Laghi, L. The Longer the Storage Time, the Higher the Price, the Better the Quality? A 1H-NMR Based Metabolomic Investigation of Aged Ya’an Tibetan Tea (Camellia Sinensis). Foods 2022, 11, 2986. [Google Scholar] [CrossRef]
- Halasa, T.; Huijps, K.; Østerås, O.; Hogeveen, H. Economic Effects of Bovine Mastitis and Mastitis Management: A Review. Vet. Q. 2007, 29, 18–31. [Google Scholar] [CrossRef]
- Wang, Y.; Nan, X.; Zhao, Y.; Wang, H.; Wang, M.; Jiang, L.; Zhang, F.; Xue, F.; Hua, D.; Li, K.; et al. Coupling 16S RDNA Sequencing and Untargeted Mass Spectrometry for Milk Microbial Composition and Metabolites from Dairy Cows with Clinical and Subclinical Mastitis. J. Agric. Food Chem. 2020, 68, 8496–8508. [Google Scholar] [CrossRef]
- Wang, Y.; Nan, X.; Zhao, Y.; Jiang, L.; Wang, H.; Zhang, F.; Hua, D.; Liu, J.; Yang, L.; Yao, J.; et al. Discrepancies among Healthy, Subclinical Mastitic, and Clinical Mastitic Cows in Fecal Microbiome and Metabolome and Serum Metabolome. J. Dairy Sci. 2022, 105, 7668–7688. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC-MS Metabolomics Identifies Metabolite Alterations That Precede Subclinical Mastitis in the Blood of Transition Dairy Cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef]
- Zhang, G.; Tobolski, D.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Identification of Serum-Predictive Biomarkers for Subclinical Mastitis in Dairy Cows and New Insights into the Pathobiology of the Disease. J. Agric. Food Chem. 2022, 70, 1724–1746. [Google Scholar] [CrossRef]
- Zwierzchowski, G.; Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Mass-Spec-Based Urinary Metabotyping around Parturition Identifies Screening Biomarkers for Subclinical Mastitis in Dairy Cows. Res. Vet. Sci. 2020, 129, 39–52. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, E.T.; Eom, J.S.; Choi, Y.Y.; Lee, S.J.; Lee, S.S.; Chung, C.D.; Lee, S.S. Exploration of Metabolite Profiles in the Biofluids of Dairy Cows by Proton Nuclear Magnetic Resonance Analysis. PLoS ONE 2021, 16, e0246290. [Google Scholar] [CrossRef]
- Eom, J.S.; Lee, S.J.; Kim, H.S.; Choi, Y.Y.; Kim, S.H.; Lee, Y.G.; Lee, S.S. Metabolomics Comparison of Hanwoo (Bos Taurus Coreanae) Biofluids Using Proton Nuclear Magnetic Resonance Spectroscopy. Metabolites 2020, 10, 333. [Google Scholar] [CrossRef]
- Sun, H.Z.; Shi, K.; Wu, X.H.; Xue, M.Y.; Wei, Z.H.; Liu, J.X.; Liu, H.Y. Lactation-Related Metabolic Mechanism Investigated Based on Mammary Gland Metabolomics and 4 Biofluids’ Metabolomics Relationships in Dairy Cows. BMC Genom. 2017, 18, 936. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Koser, S.L.; Bequette, B.J.; Donkin, S.S. Effect of Propionate on MRNA Expression of Key Genes for Gluconeogenesis in Liver of Dairy Cattle. J. Dairy Sci. 2015, 98, 8698–8709. [Google Scholar] [CrossRef] [Green Version]
- Denton, R.M.; Halestrap, A.P. Regulation of Pyruvate Metabolism in Mammalian Tissues. Essays Biochem. 1979, 15, 37–77. [Google Scholar]
- Jeyanathan, J.; Martin, C.; Morgavi, D.P. The Use of Direct-Fed Microbials for Mitigation of Ruminant Methane Emissions: A Review. Animal 2014, 8, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Grassian, A.R.; Parker, S.J.; Davidson, S.M.; Divakaruni, A.S.; Green, C.R.; Zhang, X.; Slocum, K.L.; Pu, M.; Lin, F.; Vickers, C.; et al. IDH1 Mutations Alter Citric Acid Cycle Metabolism and Increase Dependence on Oxidative Mitochondrial Metabolism. Cancer Res. 2014, 74, 3317–3331. [Google Scholar] [CrossRef] [Green Version]
- Antunes-Fernandes, E.C.; van Gastelen, S.; Dijkstra, J.; Hettinga, K.A.; Vervoort, J. Milk Metabolome Relates Enteric Methane Emission to Milk Synthesis and Energy Metabolism Pathways. J. Dairy Sci. 2016, 99, 6251–6262. [Google Scholar] [CrossRef]
- Bravo, D.M.; Wall, E.H. The Rumen and beyond: Nutritional Physiology of the Modern Dairy Cow. J. Dairy Sci. 2016, 99, 4939–4940. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, G.; Noor, S.O.; Ridgway, K.; Scovell, L.; Jamieson, C.; Johnson, I.T.; Colquhoun, I.J.; Kemsley, E.K.; Narbad, A. Metabolomics of Fecal Extracts Detects Altered Metabolic Activity of Gut Microbiota in Ulcerative Colitis and Irritable Bowel Syndrome. J. Proteome Res. 2011, 10, 4208–4218. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, T.; Han, L.; Zhao, L.; Niu, Y.; Chen, H. L-Glutamine Supplementation Alleviates Constipation during Late Gestation of Mini Sows by Modifying the Microbiota Composition in Feces. BioMed. Res. Int. 2017, 2017, 4862861. [Google Scholar] [CrossRef] [Green Version]
- Johnzon, C.F.; Dahlberg, J.; Gustafson, A.M.; Waern, I.; Moazzami, A.A.; Östensson, K.; Pejler, G. The Effect of Lipopolysaccharide-Induced Experimental Bovine Mastitis on Clinical Parameters, Inflammatory Markers, and the Metabolome: A Kinetic Approach. Front. Immunol. 2018, 9, 1487. [Google Scholar] [CrossRef]
- Haxhiaj, K.; Li, Z.; Johnson, M.; Dunn, S.M.; Wishart, D.S.; Ametaj, B.N. Blood Metabolomic Phenotyping of Dry Cows Could Predict the High Milk Somatic Cells in Early Lactation—Preliminary Results. Dairy 2022, 3, 59–77. [Google Scholar] [CrossRef]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine Transport and Fatty Acid Oxidation. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 2422–2435. [Google Scholar] [CrossRef]
- Keen, P.M.; Waterman, A.E.; Bourne, F.J. Oxygen Concentration in Milk of Healthy and Mastitic Cows and Implications of Low Oxygen Tension for the Killing of Staphylococcus Aureus by Bovine Neutrophils. J. Dairy Res. 1988, 55, 513–519. [Google Scholar] [CrossRef]
- Kalhan, S.C.; Hanson, R.W. Resurgence of Serine: An Often Neglected but Indispensable Amino Acid. J. Biol. Chem. 2012, 287, 19786–19791. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Heegaard, C.W.; Rasmussen, J.T.; Gilbert, G.E. Lactadherin Binds Selectively to Membranes Containing Phosphatidyl-L-Serine and Increased Curvature. Biochim. Biophys. Acta (BBA)-Biomembr. 2004, 1667, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Konashi, S.; Takahashi, K.; Akiba, Y. Effects of Dietary Essential Amino Acid Deficiencies on Immunological Variables in Broiler Chickens. Br. J. Nutr. 2000, 83, 449–456. [Google Scholar]
- Waisbren, S.E.; Noel, K.; Fahrbach, K.; Cella, C.; Frame, D.; Dorenbaum, A.; Levy, H. Phenylalanine Blood Levels and Clinical Outcomes in Phenylketonuria: A Systematic Literature Review and Meta-Analysis. Mol. Genet. Metab. 2007, 92, 63–70. [Google Scholar] [CrossRef]
- Brown, B. Basic Concepts in Biochemistry: A Student’s Survival Guide. Biochem. Educ. 1992, 20, 186. [Google Scholar] [CrossRef]
- Sun, H.Z.; Wang, D.M.; Wang, B.; Wang, J.K.; Liu, H.Y.; Guan, L.L.; Liu, J.X. Metabolomics of Four Biofluids from Dairy Cows: Potential Biomarkers for Milk Production and Quality. J. Proteome Res. 2015, 14, 1287–1298. [Google Scholar] [CrossRef]
- Fetherston, C.M.; Ching, T.L.; Mitoulas, L.R.; Hartmann, P.E. Excretion of Lactose in Urine as a Measure of Increased Permeability of the Lactating Breast during Inflammation. Acta Obstet. Gynecol. Scand. 2006, 85, 20–25. [Google Scholar] [CrossRef]
- Shuster, D.E.; Harmon, R.J.; Jackson, J.A.; Hemken, R.W. Suppression of Milk Production During Endotoxin-Induced Mastitis. J. Dairy Sci. 1991, 74, 3763–3774. [Google Scholar] [CrossRef]
- Miao, J.; Zhang, J.; Zheng, L.; Yu, X.; Zhu, W.; Zou, S. Taurine Attenuates Streptococcus Uberis-Induced Mastitis in Rats by Increasing T Regulatory Cells. Amino Acids 2012, 42, 2417–2428. [Google Scholar] [CrossRef]
- Moffett, J.R.; Ariyannur, P.; Arun, P.; Namboodiri, A.M.A. N-Acetylaspartate and N-Acetylaspartylglutamate in Central Nervous System Health and Disease. In Magnetic Resonance Spectroscopy: Tools for Neuroscience Research and Emerging Clinical Applications; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Urine Metabolic Fingerprinting Can Be Used to Predict the Risk of Metritis and Highlight the Pathobiology of the Disease in Dairy Cows. Metabolomics 2018, 14, 83. [Google Scholar] [CrossRef]
- Taylor, E.N.; Han, J.; Fan, C.; Beckmann, M.; Hewinson, G.; Rooke, D.; Koets, A.P.; Mur, L.A.J. Defining Fatty Acid Changes Linked to Rumen Development, Weaning and Growth in Holstein-Friesian Heifers. Metabolites 2022, 12, 374. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy (FH) | Clinical Mastitis (FM) | p-Value | Trend | |
|---|---|---|---|---|
| 1,3-Dihydroxyacetone | 4.87 × 10−5 ± 1.25 × 10−5 | 6.63 × 10−5 ± 1.48 × 10−5 | # 0.040 * | ↑ |
| 2,3-Butanediol | 4.81 × 10−5 ± 1.87 × 10−5 | 1.39 × 10−4 ± 6.12 × 10−5 | 0.001 ** | ↑ |
| Acetate | 4.61 × 10−2 ± 4.71 × 10−3 | 6.14 × 10−2 ± 1.55 × 10−2 | 0.046 * | ↑ |
| Acetoacetate | 2.44 × 10−5 ± 5.81 × 10−6 | 3.47 × 10−5 ± 7.09 × 10−6 | 0.015 * | ↑ |
| Benzoate | 5.91 × 10−4 ± 4.19 × 10−4 | 2.16 × 10−4 ± 1.97 × 10−4 | 0.011 * | ↓ |
| Creatine | 4.68 × 10−5 ± 9.63 × 10−6 | 4.20 × 10−5 ± 7.00 × 10−5 | 0.031 * | ↓ |
| Ethanol | 1.01 × 10−4 ± 2.20 × 10−5 | 1.60 × 10−4 ± 3.84 × 10−5 | 0.006 ** | ↑ |
| Glutamine | 1.95 × 10−4 ± 6.22 × 10−5 | 1.28 × 10−4 ± 2.61 × 10−5 | 0.047 * | ↓ |
| Glycine | 4.14 × 10−4 ± 8.73 × 10−5 | 1.70 × 10−4 ± 2.69 × 10−5 | 0.000 *** | ↓ |
| Methanol | 1.20 × 10−4 ± 1.75 × 10−5 | 2.11 × 10−4 ± 1.58 × 10−4 | 0.028 * | ↑ |
| O-Acetylcholine | 1.02 × 10−5 ± 1.87 × 10−6 | 6.45 × 10−6 ± 2.33 × 10−6 | 0.008 ** | ↓ |
| O-Phosphocholine | 9.84 × 10−6 ± 2.16 × 10−6 | 6.75 × 10−6 ± 2.51 × 10−6 | 0.036 * | ↓ |
| Propionate | 8.74 × 10−3 ± 9.20 × 10−4 | 1.27 × 10−2 ± 3.77 × 10−3 | 0.020 * | ↑ |
| Pyruvate | 8.19 × 10−6 ± 2.51 × 10−6 | 1.26 × 10−5 ± 3.15 × 10−6 | 0.016 * | ↑ |
| Tyrosine | 9.88 × 10−5 ± 1.52 × 10−5 | 7.95 × 10−5 ± 1.62 × 10−5 | 0.049 * | ↓ |
| Valerate | 8.92 × 10−4 ± 1.15 × 10−4 | 1.41 × 10−3 ± 4.98 × 10−4 | 0.009 ** | ↑ |
| Healthy (SH) | Clinical Mastitis (SM) | p-Value | Trend | |
|---|---|---|---|---|
| 3-Methylhistidine | 2.81 × 10−1 ± 3.28 × 10−2 | 3.49 × 10−1 ± 3.59 × 10−2 | # 0.014 * | ↑ |
| Asparagine | 1.64 × 10−1 ± 1.98 × 10−2 | 1.35 × 10−1 ± 1.49 × 10−2 | 0.036 * | ↓ |
| Citrate | 5.29 × 10−1 ± 5.32 × 10−2 | 2.80 × 10−1 ± 1.35 × 10−1 | 0.011 * | ↓ |
| Formate | 1.06 × 10−1 ± 2.89 × 10−2 | 1.54 × 10−1 ± 4.68 × 10−2 | 0.048 * | ↑ |
| Lactate | 5.31 ± 9.79 × 10−1 | 12.10 ± 7.25 | 0.029 * | ↑ |
| Phenylalanine | 6.09 × 10−1 ± 1.02 × 10−1 | 8.70 × 10−1 ± 2.32 × 10−1 | 0.035 * | ↑ |
| Serine | 3.99 × 10−1 ± 2.63 × 10−2 | 4.97 × 10−1 ± 1.21 × 10−1 | 0.045 * | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Zhang, Q.; Zhao, X.; Yang, Z.; Yang, F.; Yang, Y.; Tang, J.; Laghi, L. Metabolomic Analysis of Multiple Biological Specimens (Feces, Serum, and Urine) by 1H-NMR Spectroscopy from Dairy Cows with Clinical Mastitis. Animals 2023, 13, 741. https://doi.org/10.3390/ani13040741
Zhu C, Zhang Q, Zhao X, Yang Z, Yang F, Yang Y, Tang J, Laghi L. Metabolomic Analysis of Multiple Biological Specimens (Feces, Serum, and Urine) by 1H-NMR Spectroscopy from Dairy Cows with Clinical Mastitis. Animals. 2023; 13(4):741. https://doi.org/10.3390/ani13040741
Chicago/Turabian StyleZhu, Chenglin, Qian Zhang, Xin Zhao, Zhibo Yang, Falong Yang, Yang Yang, Junni Tang, and Luca Laghi. 2023. "Metabolomic Analysis of Multiple Biological Specimens (Feces, Serum, and Urine) by 1H-NMR Spectroscopy from Dairy Cows with Clinical Mastitis" Animals 13, no. 4: 741. https://doi.org/10.3390/ani13040741