
Serological and Molecular Survey on Domestic Dog Hepadnavirus in Household Dogs, Italy
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
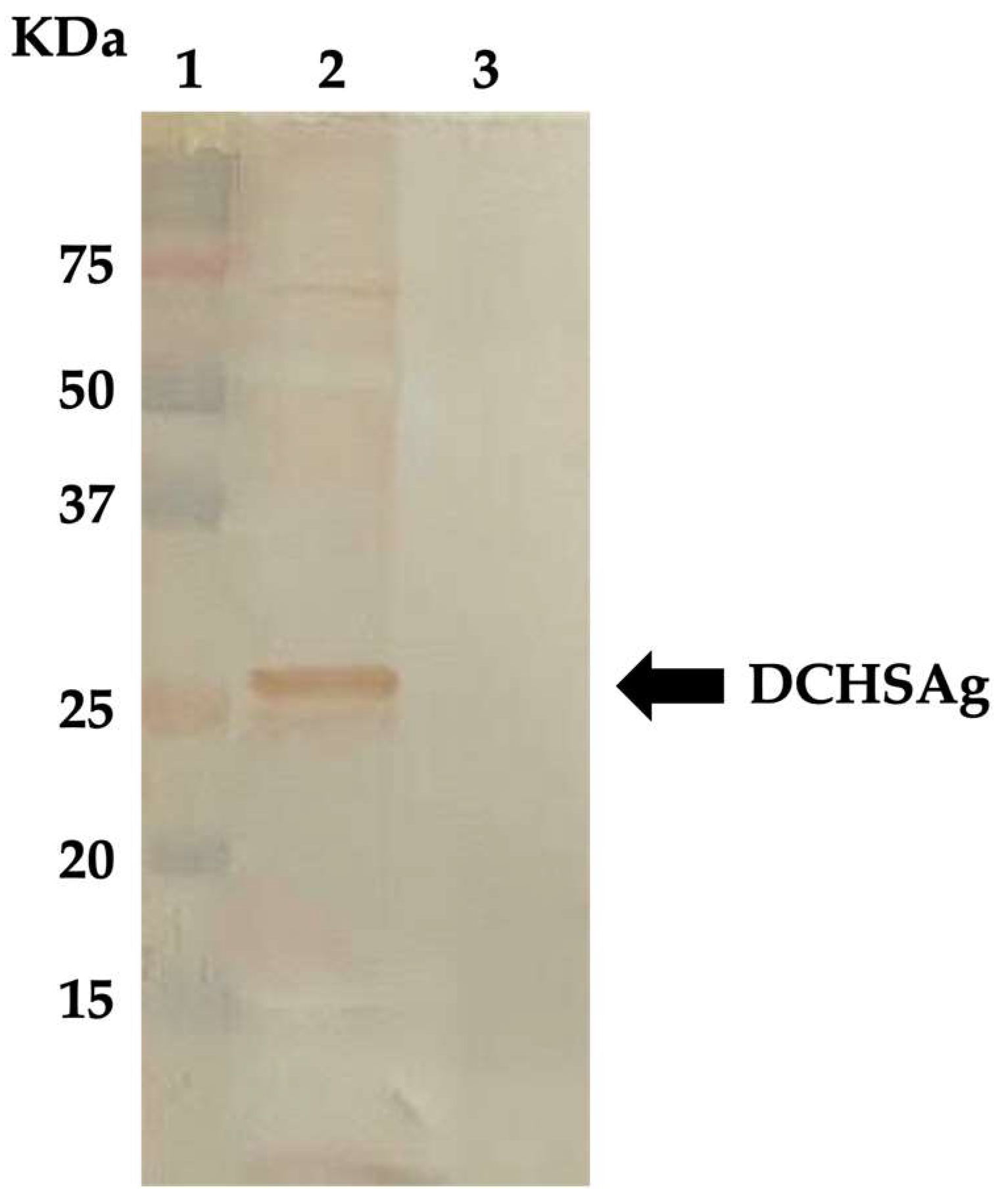
2.2. Expression of DCHSAg in the Baculovirus System and Development of Antibody Detection Enzyme-Linked Immunosorbent Assay (ELISA)
2.3. Serological Screening for Anti-DCHCAg and Anti-DCHSAg
2.4. Molecular Screening for Hepadnavirus
3. Results
3.1. Detection of Antibodies Anti-DCHCAg (DCHCAbs)
3.2. Molecular Detection and Sequence Analysis
3.3. Detection of Antibodies Anti-DCHSAg
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seeger, C.; Mason, W.S. Molecular biology of hepatitis B virus infection. Virology 2015, 479–480, 672–686. [Google Scholar] [CrossRef] [Green Version]
- Magnius, L.; Mason, W.S.; Taylor, J.; Kann, M.; Glebe, D.; Dény, P.; Sureau, C.; Norder, H.; Ictv Report Consortium. ICTV Virus Taxonomy Profile: Hepadnaviridae. J. Gen. Virol. 2020, 101, 571–572. [Google Scholar] [CrossRef]
- WHO. Hepatitis B. Available online: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b (accessed on 15 November 2022).
- Drexler, J.F.; Geipel, A.; König, A.; Corman, V.M.; van Riel, D.; Leijten, L.M.; Bremer, C.M.; Rasche, A.; Cottontail, V.M.; Maganga, G.D.; et al. Bats carry pathogenic hepadnaviruses antigenically related to hepatitis B virus and capable of infecting human hepatocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 16151–16156. [Google Scholar] [CrossRef] [Green Version]
- He, B.; Fan, Q.; Yang, F.; Hu, T.; Qiu, W.; Feng, Y.; Li, Z.; Li, Y.; Zhang, F.; Guo, H.; et al. Hepatitis virus in long-fingered bats, Myanmar. Emerg. Infect. Dis. 2013, 19, 638–640. [Google Scholar] [CrossRef]
- Wang, B.; Yang, X.L.; Li, W.; Zhu, Y.; Ge, X.Y.; Zhang, L.B.; Zhang, Y.Z.; Bock, C.T.; Shi, Z.L. Detection and genome characterization of four novel bat hepadnaviruses and a hepevirus in China. Virol. J. 2017, 14, 40. [Google Scholar] [CrossRef] [Green Version]
- Nie, F.Y.; Lin, X.D.; Hao, Z.Y.; Chen, X.N.; Wang, Z.X.; Wang, M.R.; Wu, J.; Wang, H.W.; Zhao, G.; Ma, R.Z.; et al. Extensive diversity and evolution of hepadnaviruses in bats in China. Virology 2018, 514, 88–97. [Google Scholar] [CrossRef]
- Gogarten, J.F.; Ulrich, M.; Bhuva, N.; Garcia, J.; Jain, K.; Lee, B.; Löhrich, T.; Oleynik, A.; Couacy-Hymann, E.; Fuh Neba, T.; et al. A Novel Orthohepadnavirus Identified in a dead Maxwell’s Duiker (Philantomba maxwellii) in Taï National Park, Côte d’Ivoire. Viruses 2019, 11, 279. [Google Scholar] [CrossRef] [Green Version]
- Rasche, A.; Lehmann, F.; Goldmann, N.; Nagel, M.; Moreira-Soto, A.; Nobach, D.; de Oliveira Carneiro, I.; Osterrieder, N.; Greenwood, A.D.; Steinmann, E.; et al. A hepatitis B virus causes chronic infections in equids worldwide. Proc. Natl. Acad. Sci. USA 2021, 118, e2013982118. [Google Scholar] [CrossRef]
- Aghazadeh, M.; Shi, M.; Barrs, V.R.; McLuckie, A.J.; Lindsay, S.A.; Jameson, B.; Hampson, B.; Holmes, E.C.; Beatty, J.A. A Novel Hepadnavirus identified in an immunocompromised domestic cat in Australia. Viruses 2018, 10, 269. [Google Scholar] [CrossRef] [Green Version]
- Vieira, Y.R.; Portilho, M.M.; Oliveira, F.F.; Guterres, A.; Dos Santos, D.R.L.; Villar, L.M.; Mirazo, S.; Arbiza, J.; Dimache, L.A.G.; Almeida, F.Q.; et al. Evaluation of HBV-Like circulation in wild and farm animals from Brazil and Uruguay. Int. J. Environ. Res. Public Health 2019, 16, 2679. [Google Scholar] [CrossRef] [Green Version]
- Diakoudi, G.; Capozza, P.; Lanave, G.; Pellegrini, F.; Di Martino, B.; Elia, G.; Decaro, N.; Camero, M.; Ghergo, P.; Stasi, F.; et al. A novel hepadnavirus in domestic dogs. Sci. Rep. 2022, 12, 2864. [Google Scholar] [CrossRef]
- Lanave, G.; Capozza, P.; Diakoudi, G.; Catella, C.; Catucci, L.; Ghergo, P.; Stasi, F.; Barrs, V.; Beatty, J.; Decaro, N.; et al. Identification of hepadnavirus in the sera of cats. Sci. Rep. 2019, 9, 10668. [Google Scholar] [CrossRef] [Green Version]
- Fruci, P.; Di Profio, F.; Palombieri, A.; Massirio, I.; Lanave, G.; Diakoudi, G.; Pellegrini, F.; Marsilio, F.; Martella, V.; Di Martino, B. Detection of antibodies against domestic cat hepadnavirus using baculovirus-expressed core protein. Transbound. Emerg. Dis. 2022, 69, 2980–2986. [Google Scholar] [CrossRef]
- Piewbang, C.; Wardhani, S.W.; Chaiyasak, S.; Yostawonkul, J.; Chai-In, P.; Boonrungsiman, S.; Kasantikul, T.; Techangamsuwan, S. Insights into the genetic diversity, recombination, and systemic infections with evidence of intracellular maturation of hepadnavirus in cats. PLoS ONE 2020, 15, e0241212. [Google Scholar] [CrossRef]
- Anpuanandam, K.; Selvarajah, G.T.; Choy, M.M.K.; Ng, S.W.; Kumar, K.; Ali, R.M.; Rajendran, S.K.; Ho, K.L.; Tan, W.S. Molecular detection and characterisation of Domestic Cat Hepadnavirus (DCH) from blood and liver tissues of cats in Malaysia. BMC Vet. Res. 2021, 17, 9. [Google Scholar] [CrossRef]
- Takahashi, K.; Kaneko, Y.; Shibanai, A.; Yamamoto, S.; Katagiri, A.; Osuga, T.; Inoue, Y.; Kuroda, K.; Tanabe, M.; Okabayashi, T.; et al. Identification of domestic cat hepadnavirus from a cat blood sample in Japan. J. Vet. Med. Sci. 2022, 84, 648–652. [Google Scholar] [CrossRef]
- Stone, C.; Petch, R.; Gagne, R.B.; Nehring, M.; Tu, T.; Beatty, J.A.; VandeWoude, S. Prevalence and genomic sequence analysis of Domestic Cat Hepadnavirus in the United States. Viruses 2022, 14, 2091. [Google Scholar] [CrossRef]
- Pesavento, P.A.; Jackson, K.; Hampson, T.S.T.T.B.; Munday, J.S.; Barrs, V.R.; Beatty, J.A. A novel hepadnavirus is associated with chronic hepatitis and hepatocellular carcinoma in cats. Viruses 2019, 11, 969. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.R.; Chen, M.C.; Carrai, M.; Rizzo, F.; Chai, Y.; Tse, M.; Jackson, K.; Martella, V.; Steiner, J.; Pesavento, P.A.; et al. Hepadnavirus DNA is detected in canine blood samples in Hong Kong but not in liver biopsies of chronic hepatitis or hepatocellular carcinoma. Viruses 2022, 14, 1543. [Google Scholar] [CrossRef]
- Kao, J.H. Diagnosis of hepatitis B virus infection through serological and virological markers. Expert Rev. Gastroenterol. Hepatol. 2008, 2, 553–562. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, H.S. Current laboratory tests for diagnosis of hepatitis B virus infection. Int. J. Clin. Pract. 2021, 75, e14812. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for the Prevention, Care and Treatment of Persons with Chronic Hepatitis B Infection. Available online: https://www.who.int/publications/i/item/9789241549059 (accessed on 15 November 2022).
- Wang, Q.; Klenerman, P.; Semmo, N. Significance of anti-HBc alone serological status in clinical practice. Lancet Gastroenterol. Hepatol. 2017, 2, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Hoofnagle, J.H.; Schafer, D.F.; Ferenci, P.; Waggoner, J.G.; Vergalla, J.; April, M.; Phillips, L. Antibody to hepatitis B surface antigen in nonprimate animal species. Gastroenterology 1983, 84, 1478–1482. [Google Scholar] [CrossRef]
- Chen, D.S.; Sung, J.L.; Lai, M.Y.; Liu, M.R. Antibody to hepatitis B surface antigen in dogs in Taiwan. Gastroenterology 1984, 86, 1001–1003. [Google Scholar]
- Al-Jumaa, Z.M.; Ajaj, E.A.; Dahl, M.O. Seroprevalence of canine hepatitis in stray dogs in Nineveh Province, Iraq. Vet. World 2020, 13, 2326–2329. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| DCHCAbs | Positive | Negative |
|---|---|---|
| IgM anti-DCHCAg | 13 (2.2%) | 587 (97.8%) |
| IgG anti-DCHCAg | 45 (7.5%) | 555 (92.5%) |
| IgM and IgG anti-DCHCAg | 2 (0.3%) | 598 (99.7%) |
| Total | 60 (10.0%) | 540 (90.0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fruci, P.; Palombieri, A.; Sarchese, V.; Aste, G.; Friedrich, K.G.; Martella, V.; Di Martino, B.; Di Profio, F. Serological and Molecular Survey on Domestic Dog Hepadnavirus in Household Dogs, Italy. Animals 2023, 13, 729. https://doi.org/10.3390/ani13040729
Fruci P, Palombieri A, Sarchese V, Aste G, Friedrich KG, Martella V, Di Martino B, Di Profio F. Serological and Molecular Survey on Domestic Dog Hepadnavirus in Household Dogs, Italy. Animals. 2023; 13(4):729. https://doi.org/10.3390/ani13040729
Chicago/Turabian StyleFruci, Paola, Andrea Palombieri, Vittorio Sarchese, Giovanni Aste, Klaus G. Friedrich, Vito Martella, Barbara Di Martino, and Federica Di Profio. 2023. "Serological and Molecular Survey on Domestic Dog Hepadnavirus in Household Dogs, Italy" Animals 13, no. 4: 729. https://doi.org/10.3390/ani13040729