

Urban Bird Community Assembly Mechanisms and Driving Factors in University Campuses in Nanjing, China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
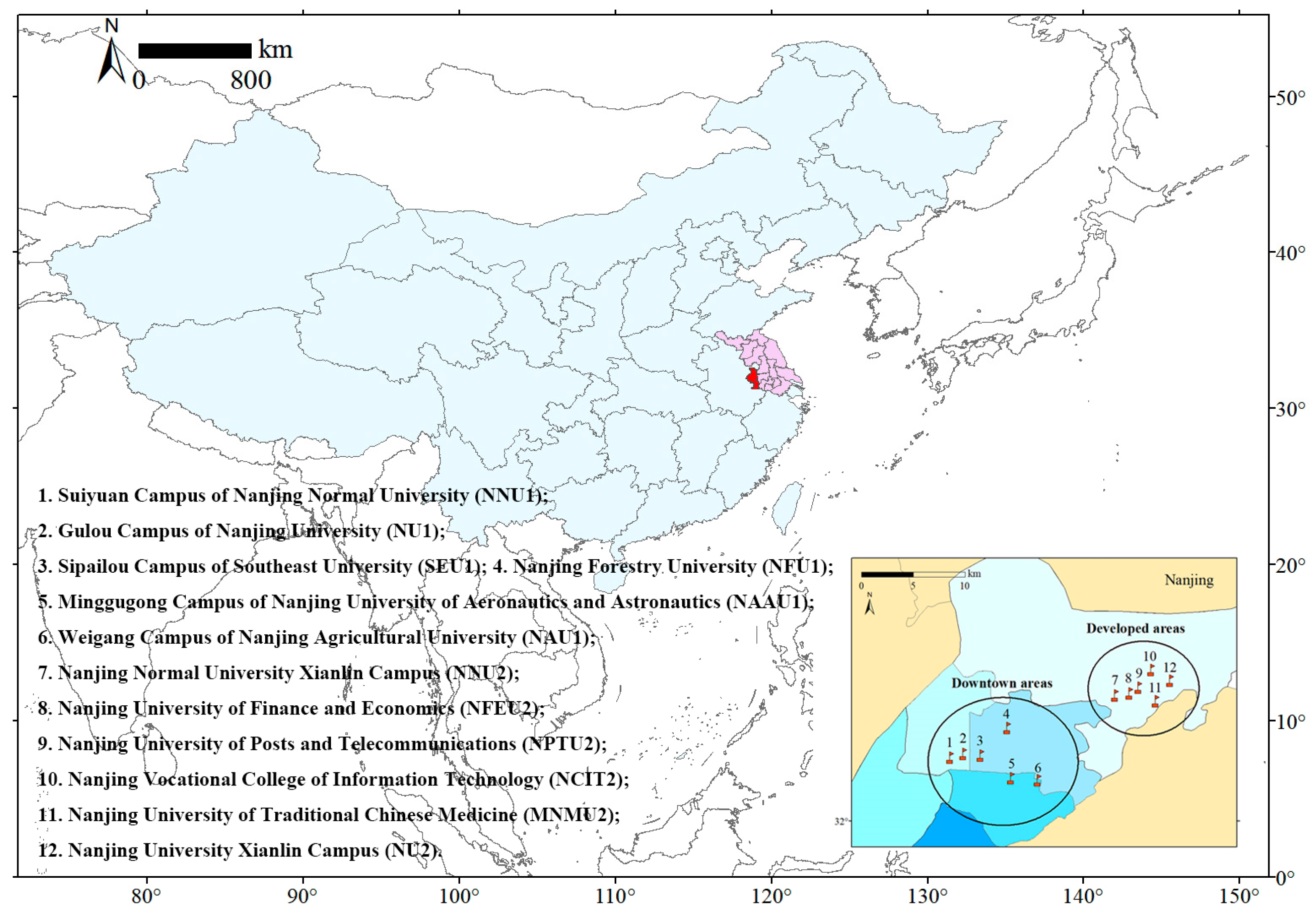
2.1. Study Area
2.2. Data Collection
2.2.1. Landscape Configuration
2.2.2. Bird Counts
2.2.3. Diversity Metrics of Bird Assemblage
2.3. Statistical Analyses
3. Results
3.1. Landscape Metrics
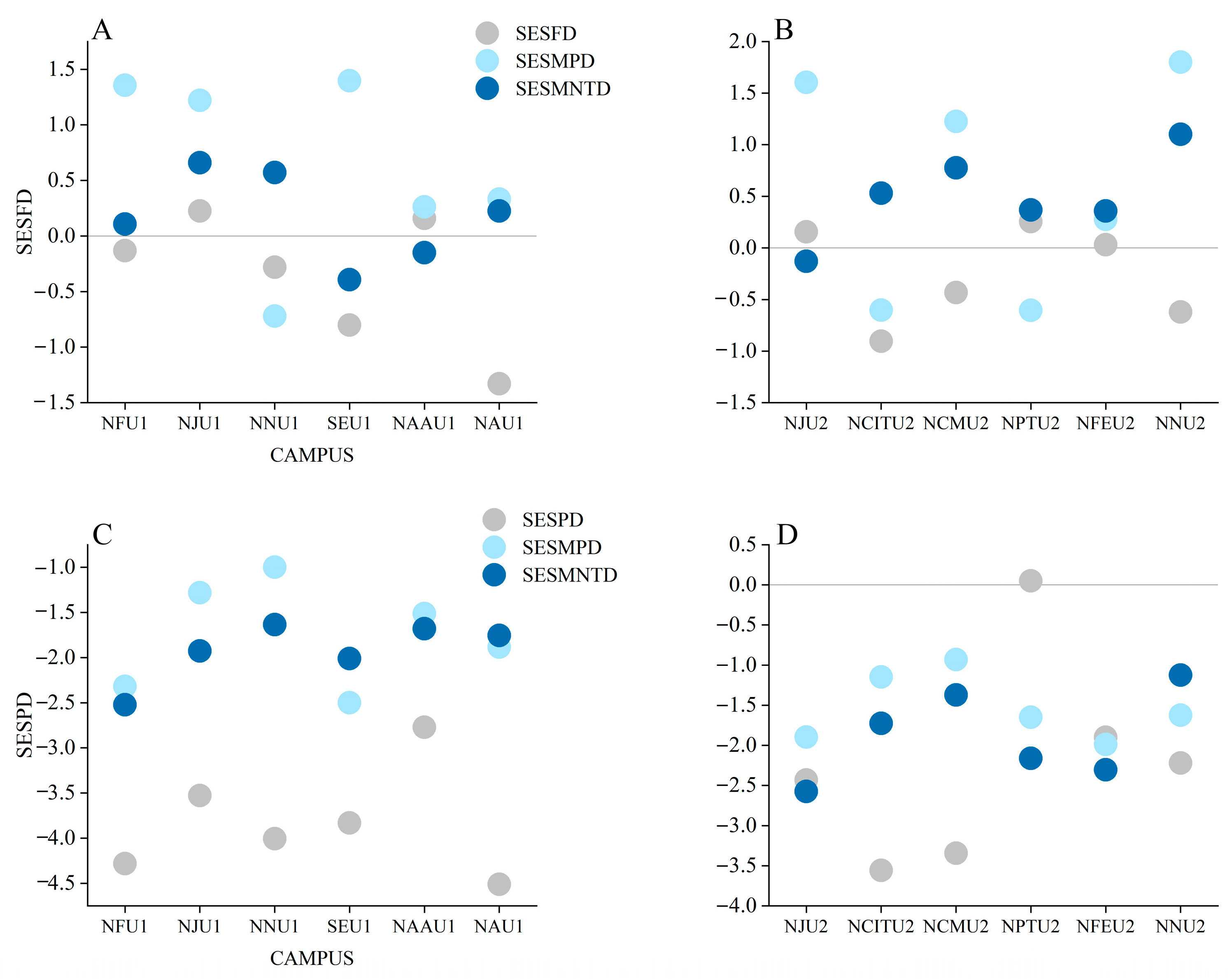
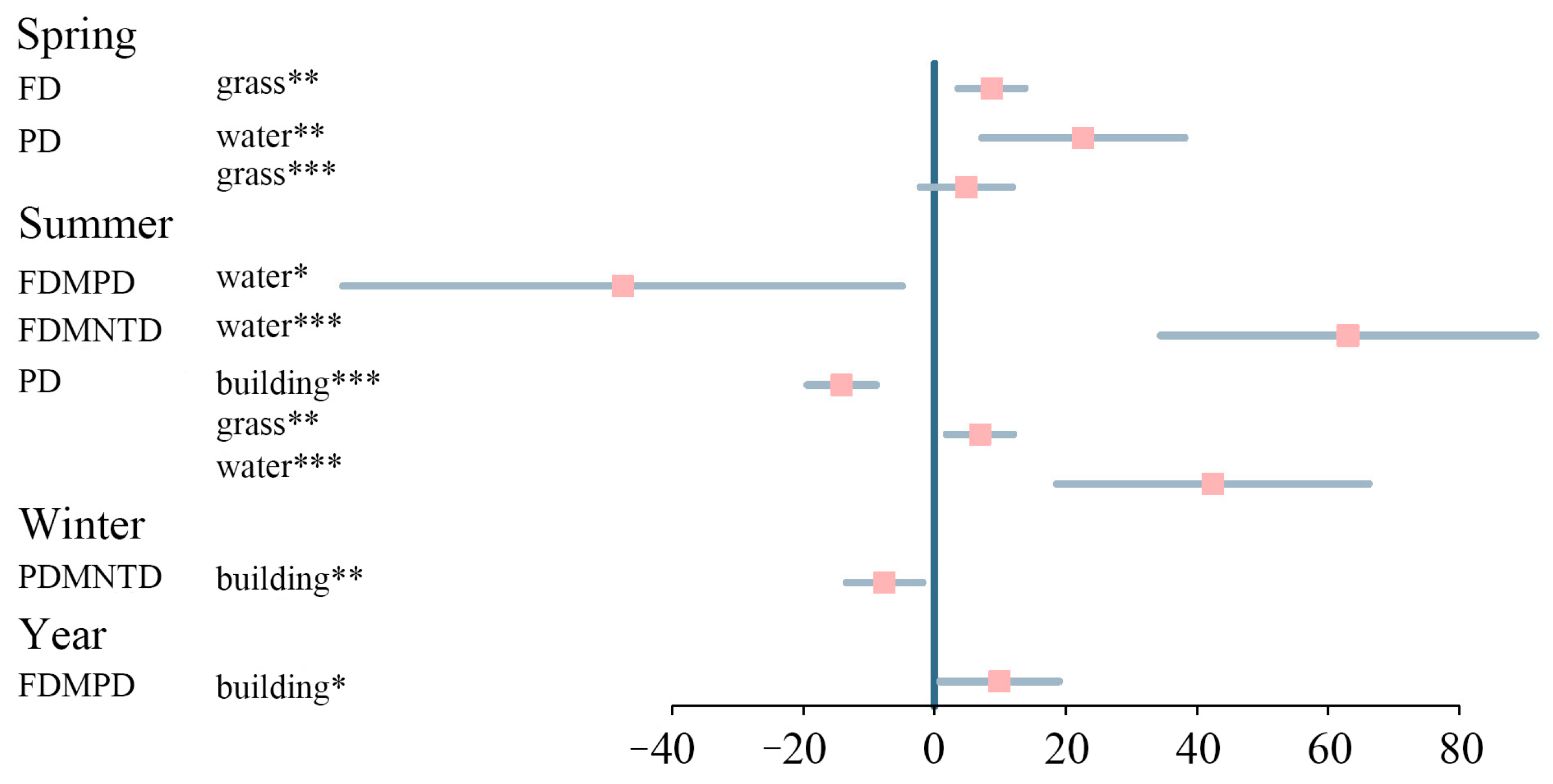
3.2. Bird Community Composition, Assembly Processes, and Biodiversity
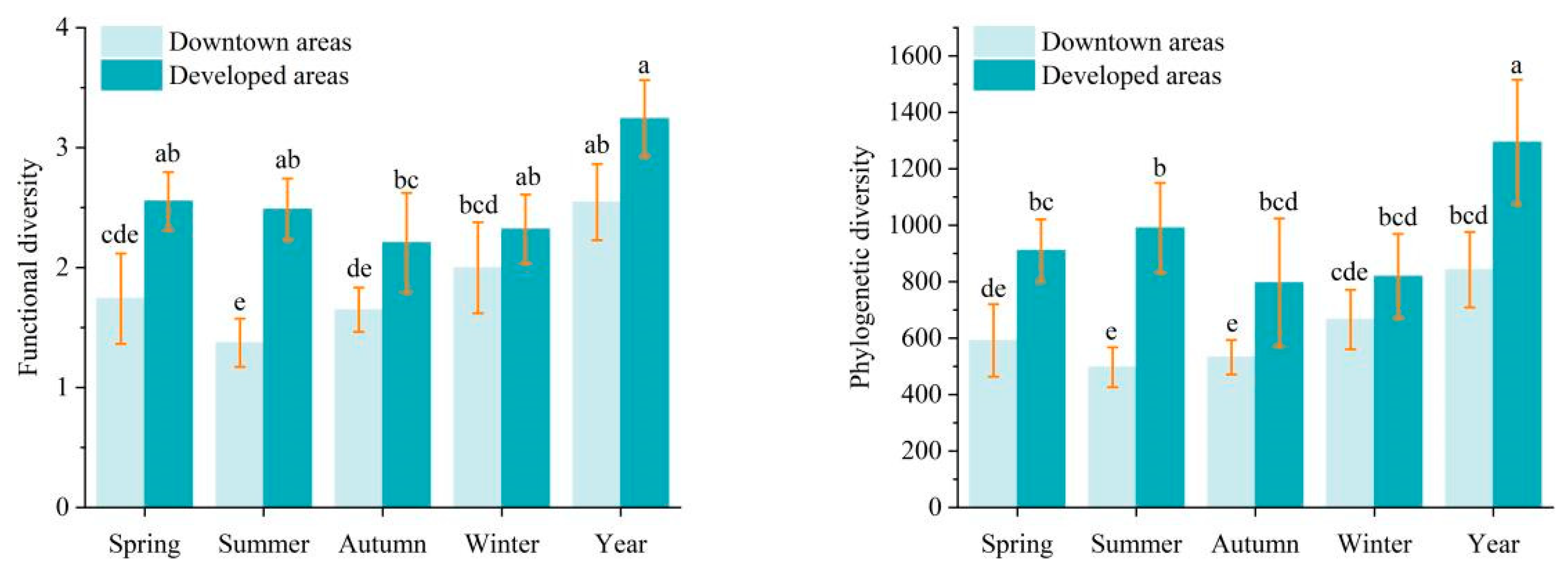
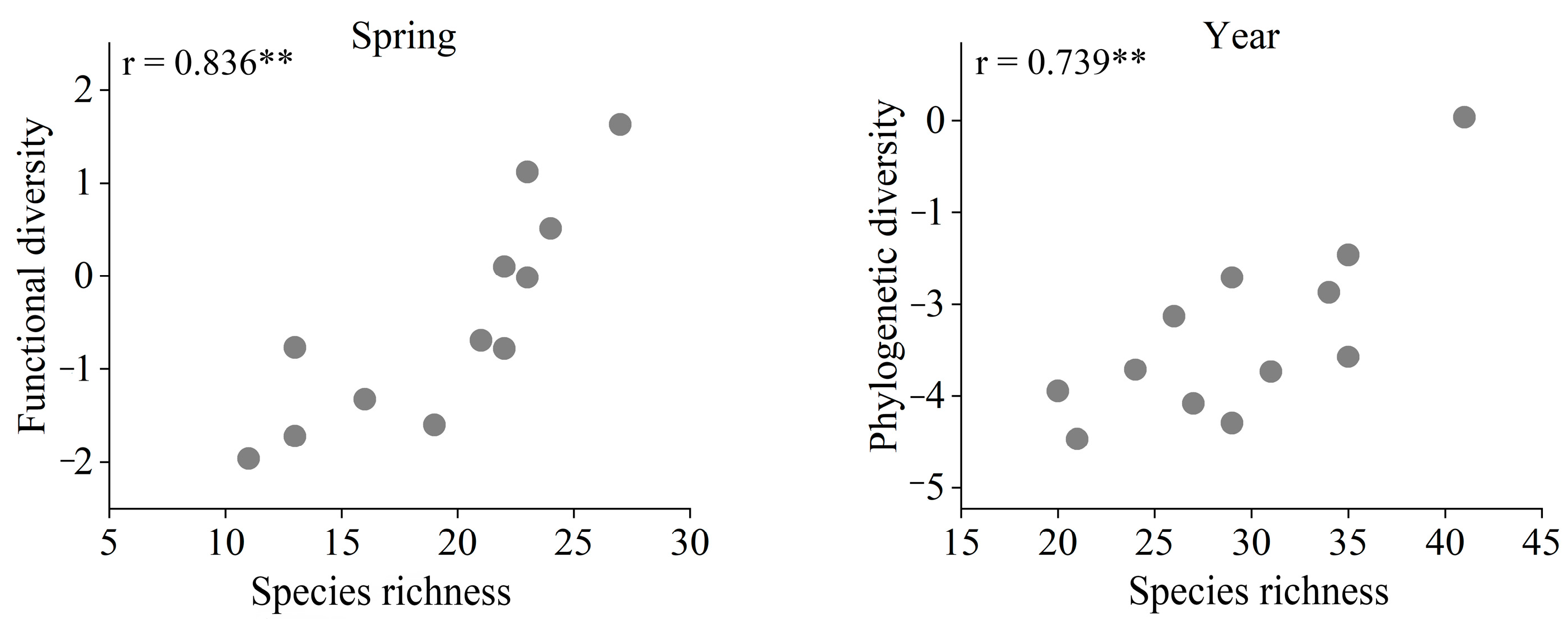
3.3. Bird Communities Functional and Phylogenetic Diversity
4. Discussion
4.1. Landscape Metrics
4.2. Bird Diversity and Residential Types
4.3. Differences in Bird Functional and Phylogenetic Diversity in Newly Developed and Downtown Areas
4.4. Relationship between Landscape Metrics and Diversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cowie, R.; Bouchet, P.; Fontaine, B. The Sixth Mass Extinction: Fact, fiction or speculation? Biol. Rev. 2022, 97, 640–663. [Google Scholar] [CrossRef]
- García, F.C.; Bestion, E.; Warfield, R.; Yvon-Durocher, G. Changes in temperature alter the relationship between biodiversity and ecosystem functioning. Proc. Natl. Acad. Sci. USA 2018, 115, 10989–10994. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves Souza, D.; Verburg, P.H.; Dobrovolski, R. Habitat loss, extinction predictability and conservation efforts in the terrestrial ecoregions. Biol. Conserv. 2020, 246, 108579. [Google Scholar] [CrossRef]
- Cavender Bares, J.; Kozak, K.H.; Fine, P.V.; Kembel, S.W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 2009, 12, 693–715. [Google Scholar] [CrossRef]
- Maitner, B.S.; Rudgers, J.A.; Dunham, A.E.; Whitney, K.D. Patterns of bird invasion are consistent with environmental filtering. Ecography 2012, 35, 614–623. [Google Scholar] [CrossRef]
- Mayfield, M.M.; Levine, J.M. Opposing effects of competitive exclusion on the phylogenetic structure of communities. Ecol. Lett. 2010, 13, 1085–1093. [Google Scholar] [CrossRef]
- Hubbell, S.P. Neutral theory and the evolution of ecological equivalence. Ecology 2006, 87, 1387–1398. [Google Scholar] [CrossRef] [Green Version]
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. B 2011, 366, 2351–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebrija Trejos, E.; Pérez-García, E.A.; Meave, J.A.; Bongers, F.; Poorter, L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology 2010, 91, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Si, X.; Baselga, A.; Leprieur, F.; Song, X.; Ding, P. Selective extinction drives taxonomic and functional alpha and beta diversities in island bird assemblages. J. Anim. Ecol. 2016, 85, 409–418. [Google Scholar] [CrossRef]
- Wosnick, N.; Leite, R.D.; Giareta, E.P.; Nunes, A.R.O.; Nunes, J.L.; Charvet, P.; Monteiro-Filho, E.L. Evaluating conservation status and governmental efforts towards regional flagship species in Brazil. J. Environ. Manag. 2021, 292, 112732. [Google Scholar] [CrossRef]
- Swan, C.M.; Brown, B.; Borowy, D.; Cavender-Bares, J.; Jeliazkov, A.; Knapp, S.; Lososová, Z.; Padullés Cubino, J.; Pavoine, S.; Ricotta, C. A framework for understanding how biodiversity patterns unfold across multiple spatial scales in urban ecosystems. Ecosphere 2021, 12, e03650. [Google Scholar] [CrossRef]
- Sandström, U.; Angelstam, P.; Mikusiński, G. Ecological diversity of birds in relation to the structure of urban green space. Landsc. Urban Plan 2006, 77, 39–53. [Google Scholar] [CrossRef]
- De Groot, M.; Flajšman, K.; Mihelič, T.; Vilhar, U.; Simončič, P.; Verlič, A. Green space area and type affect bird communities in a South-eastern European city. Urban For. Urban Green. 2021, 63, 127212. [Google Scholar] [CrossRef]
- Xu, H.; Liu, K.; Ning, T.; Huang, G.; Zhang, Q.; Li, Y.; Wang, M.; Fan, Y.; An, W.; Ji, L. Environmental remediation promotes the restoration of biodiversity in the Shenzhen Bay Estuary, South China. Ecosyst. Health Sustain. 2022, 8, 2026250. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, Q.-k.; Gao, L.; Li, X.-y.; Meng, L.-l.; Peng, K.-k. A multiobjective evolutionary algorithm based on decomposition for hybrid flowshop green scheduling problem. Comput. Ind. Eng. 2019, 136, 325–344. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Ibáñez-Álamo, J.D.; Tryjanowski, P.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Suhonen, J.; Díaz, M.; Møller, A.P.; Moravec, D. Effects of urbanization on taxonomic, functional and phylogenetic avian diversity in Europe. Sci. Total. Environ. 2021, 795, 148874. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Bino, G.; Major, R.E.; Martin, J.M.; Lyons, M.B.; Kingsford, R.T. Heterogeneous urban green areas are bird diversity hotspots: Insights using continental-scale citizen science data. Landsc. Ecol. 2019, 34, 1231–1246. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, C.; Chen, S.; Zhang, Y.; Shi, H.; Chen, B.; Mao, L. Effects of Landscape Attributes on Campuses Bird Species Richness and Diversity, Implications for Eco-Friendly Urban Planning. Sustainability 2021, 13, 5558. [Google Scholar] [CrossRef]
- Zhang, W.; Liang, C.; Liu, J.; Si, X.; Feng, G. Species richness, phylogenetic and functional structure of bird communities in Chinese university campuses are associated with divergent variables. Urban Ecosyst. 2018, 21, 1213–1225. [Google Scholar] [CrossRef]
- Chong, K.Y.; Teo, S.; Kurukulasuriya, B.; Chung, Y.F.; Rajathurai, S.; Tan, H.T.W. Not all green is as good: Different effects of the natural and cultivated components of urban vegetation on bird and butterfly diversity. Biol. Conserv. 2014, 171, 299–309. [Google Scholar] [CrossRef]
- Pedersen, C.; Krøgli, S.O. The effect of land type diversity and spatial heterogeneity on farmland birds in Norway. Ecol. Indic. 2017, 75, 155–163. [Google Scholar] [CrossRef]
- Fan, Z.; Duan, J.; Lu, Y.; Zou, W.; Lan, W. A geographical detector study on factors influencing urban park use in Nanjing, China. Urban For. Urban Green. 2021, 59, 126996. [Google Scholar] [CrossRef]
- Ye, C.; Chen, M.; Chen, R.; Guo, Z. Multi-scalar separations: Land use and production of space in Xianlin, a university town in Nanjing, China. Habitat Int. 2014, 42, 264–272. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Y.; Zha, D.; Yang, S.; Huang, Z.Y.; de Boer, W.F. Assembly processes of waterbird communities across subsidence wetlands in China: A functional and phylogenetic approach. Divers. Distrib. 2019, 25, 1118–1129. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Song, Y.; Chen, C.; Zhao, Y.; Zeng, D.; Wu, Y.; Ding, P. A dataset on life-history and ecological traits of Chinese birds. Biodivers. Sci. 2021, 29, 1149–1153. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity (FD), species richness and community composition. Ecol. Lett. 2002, 5, 402–411. [Google Scholar] [CrossRef]
- Srivastava, D.S.; Cadotte, M.W.; MacDonald, A.A.M.; Marushia, R.G.; Mirotchnick, N. Phylogenetic diversity and the functioning of ecosystems. Ecol. Lett. 2012, 15, 637–648. [Google Scholar] [CrossRef]
- Revell, L.J.; Harmon, L.J.; Collar, D.C. Phylogenetic signal, evolutionary process, and rate. Syst. Biol. 2008, 57, 591–601. [Google Scholar] [CrossRef]
- MacArthur, R.; Levins, R. The limiting similarity, convergence, and divergence of coexisting species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Gomez, J.P.; Bravo, G.A.; Brumfield, R.T.; Tello, J.G.; Cadena, C.D. A phylogenetic approach to disentangling the role of competition and habitat filtering in community assembly of Neotropical forest birds. J. Anim. Ecol. 2010, 79, 1181–1192. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, C.T.; Major, R.E.; Lyons, M.B.; Martin, J.M.; Kingsford, R.T. The effects of local and landscape habitat attributes on bird diversity in urban greenspaces. Ecosphere 2018, 9, e02347. [Google Scholar] [CrossRef] [Green Version]
- Barton, K.; Barton, M.K. Package ‘mumin’. Version 2015, 1, 439. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- Aronson, M.F.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.; Cilliers, S.; Clarkson, B. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. Roy. Soc. B-Biol. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef]
- Taylor, J.J.; Lepczyk, C.A.; Brown, D.G. Patch and matrix level influences on forest birds at the rural–urban interface. Landsc. Ecol. 2016, 31, 1005–1020. [Google Scholar] [CrossRef]
- Strohbach, M.W.; Lerman, S.B.; Warren, P.S. Are small greening areas enhancing bird diversity? Insights from community-driven greening projects in Boston. Landsc. Urban Plan. 2013, 114, 69–79. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin III, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.; Loreau, M.; Naeem, S. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Perna, P.; Santolini, R. Associations among taxonomic diversity, functional diversity and evolutionary distinctiveness vary among environments. Ecol. Indic. 2018, 88, 8–16. [Google Scholar] [CrossRef]
- Farias, A.A.; Jaksic, F.M. Low functional richness and redundancy of a predator assemblage in native forest fragments of Chiloe Island, Chile. J. Anim. Ecol. 2011, 80, 809–817. [Google Scholar] [CrossRef]
- Pacheco-Muñoz, R.; Aguilar-Gómez, M.Á.; Schondube, J.E. Overwintering in a megacity: Urban green areas and migratory birds in Mexico City. Urban For. Urban Green. 2022, 73, 127614. [Google Scholar] [CrossRef]
- Violle, C.; Nemergut, D.R.; Pu, Z.; Jiang, L. Phylogenetic limiting similarity and competitive exclusion. Ecol. Lett. 2011, 14, 782–787. [Google Scholar] [CrossRef]
- Gábor, L.; Moudrý, V.; Barták, V.; Lecours, V. How do species and data characteristics affect species distribution models and when to use environmental filtering? Int. J. Geogr. Inf. Sci. 2020, 34, 1567–1584. [Google Scholar] [CrossRef]
- Freeman, B.G.; Weeks, T.; Schluter, D.; Tobias, J.A. The latitudinal gradient in rates of evolution for bird beaks, a species interaction trait. Ecol. Lett. 2022, 25, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Edelaar, P.; Siepielski, A.M.; Clobert, J. Matching habitat choice causes directed gene flow: A neglected dimension in evolution and ecology. Evolution 2008, 62, 2462–2472. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, Y.; Si, X.; Feng, G.; Slik, F.; Zhang, J. University campuses as valuable resources for urban biodiversity research and conservation. Urban For. Urban Green. 2021, 64, 127255. [Google Scholar] [CrossRef]
- Klingbeil, B.T.; Willig, M.R. Community assembly in temperate forest birds: Habitat filtering, interspecific interactions and priority effects. Evol. Ecol. 2016, 30, 703–722. [Google Scholar] [CrossRef]
- Xu, C.; Huang, Z.Y.; Chi, T.; Chen, B.J.; Zhang, M.; Liu, M. Can local landscape attributes explain species richness patterns at macroecological scales? Global Ecol. Biogeogr. 2014, 23, 436–445. [Google Scholar] [CrossRef]
- Jakobsson, S.; Wood, H.; Ekroos, J.; Lindborg, R. Contrasting multi-taxa functional diversity patterns along vegetation structure gradients of woody pastures. Biodivers. Conserv. 2020, 29, 3551–3572. [Google Scholar] [CrossRef]
- Osorio-Olvera, L.; Yañez-Arenas, C.; Martínez-Meyer, E.; Peterson, A.T. Relationships between population densities and niche-centroid distances in North American birds. Ecol. Lett. 2020, 23, 555–564. [Google Scholar] [CrossRef]
- Curzel, F.E.; Bellocq, M.I.; Leveau, L.M. Local and landscape features of wooded streets influenced bird taxonomic and functional diversity. Urban For. Urban Green. 2021, 66, 127369. [Google Scholar] [CrossRef]
- Hager, S.B.; Cosentino, B.J.; Aguilar-Gómez, M.A.; Anderson, M.L.; Bakermans, M.; Boves, T.J.; Brandes, D.; Butler, M.W.; Butler, E.M.; Cagle, N.L. Continent-wide analysis of how urbanization affects bird-window collision mortality in North America. Biol. Conserv. 2017, 212, 209–215. [Google Scholar] [CrossRef]
- Xu, Y.; Si, Y.; Wang, Y.; Zhang, Y.; Prins, H.H.; Cao, L.; de Boer, W.F. Loss of functional connectivity in migration networks induces population decline in migratory birds. Ecol. Appl. 2019, 29, e01960. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landscape Metrics | Abbreviations | Brief Description |
|---|---|---|
| Total area | Ta | The total area of the campus |
| Forest area | Forest | Woody plant area including trees and shrubs |
| Grass area | Grass | Grass and lawn area |
| Building area | Building | Building and structure area |
| Impervious surface area | Isurf | Impervious surface area, including parking areas, playgrounds, and squares |
| Water body area | Water | Water body area, including ponds, rivers, and urban lakes |
| Patch density | Pd | The density of a patch in the landscape |
| Largest patch index | Lpi | Dominant patch types in the landscape |
| Edge density | Ed | Landscape fragmentation degree |
| Mean patch area | Marea | Average area of a patch in the landscape |
| Proportion of similar adjacency | Pladj | Patch proximity and landscape fragmentation |
| Connectance index | Connect | Patch connectivity |
| Splitting index | Split | Separation degree of patch distribution |
| Aggregation index | Ai | Connectivity between patches of landscape types |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Borzée, A.; Li, J.; Chen, S.; Shi, H.; Zhang, Y. Urban Bird Community Assembly Mechanisms and Driving Factors in University Campuses in Nanjing, China. Animals 2023, 13, 673. https://doi.org/10.3390/ani13040673
Zhao Z, Borzée A, Li J, Chen S, Shi H, Zhang Y. Urban Bird Community Assembly Mechanisms and Driving Factors in University Campuses in Nanjing, China. Animals. 2023; 13(4):673. https://doi.org/10.3390/ani13040673
Chicago/Turabian StyleZhao, Zixi, Amaël Borzée, Jinghao Li, Sheng Chen, Hui Shi, and Yong Zhang. 2023. "Urban Bird Community Assembly Mechanisms and Driving Factors in University Campuses in Nanjing, China" Animals 13, no. 4: 673. https://doi.org/10.3390/ani13040673