
Treatment of Inactive Ovaries of Holstein Dairy Cows by Epidural Injection of GnRH Analogue (Receptal) and Its Impact on the Reproductive Hormones, Oxidant/Antioxidant Profile and Micro and Macro-Elements Profile
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. Blood Samples and Experimental Protocol
- -
- Group Repid (treated group): 30 cows were administered GnRH agonist by epidural route in the sacrococcygeal space (5 mL Receptal per head, each 1 mL contained 0.004 mg Buserelin, Intervet, Holland).
- -
- Group Cepid (control group): 30 cows were administered 5 mL of normal saline solution by epidural route in the sacrococcygeal space (0.9% NaCl).
2.3. Determination of Hormonal Profile in Blood
2.4. Determinations of Oxidant/Antioxidant Levels and the Mineral Profile in Blood
2.5. Statistical Evaluation
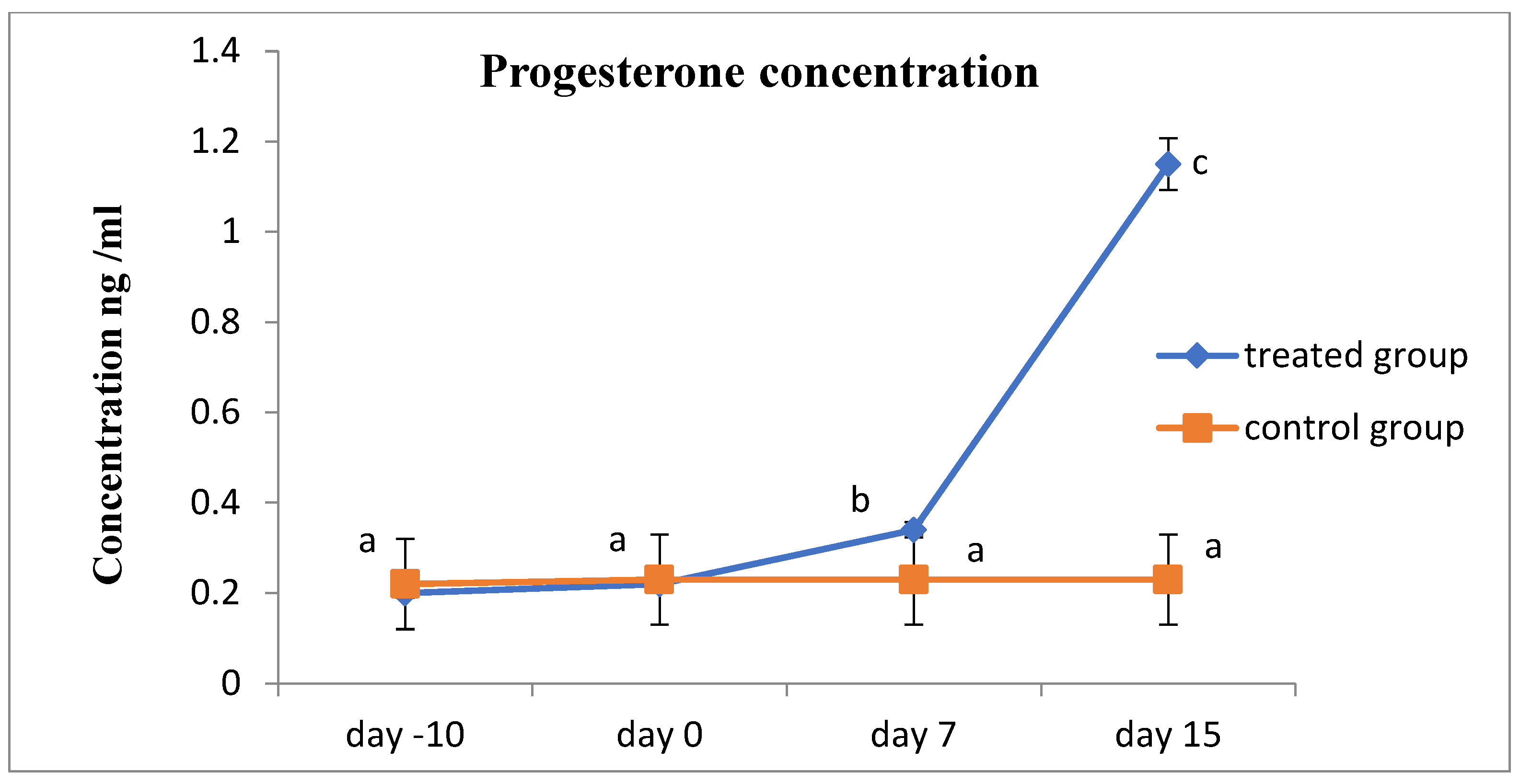
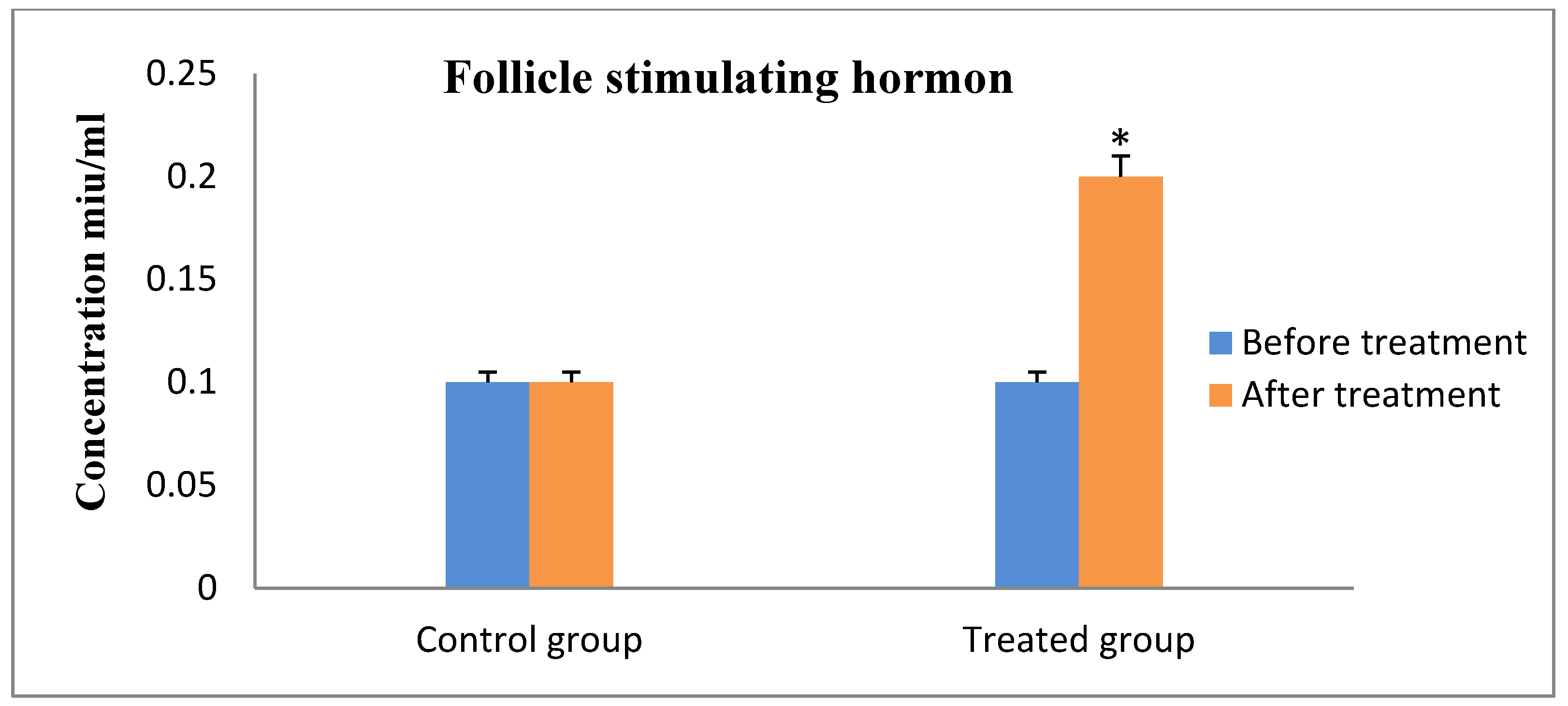
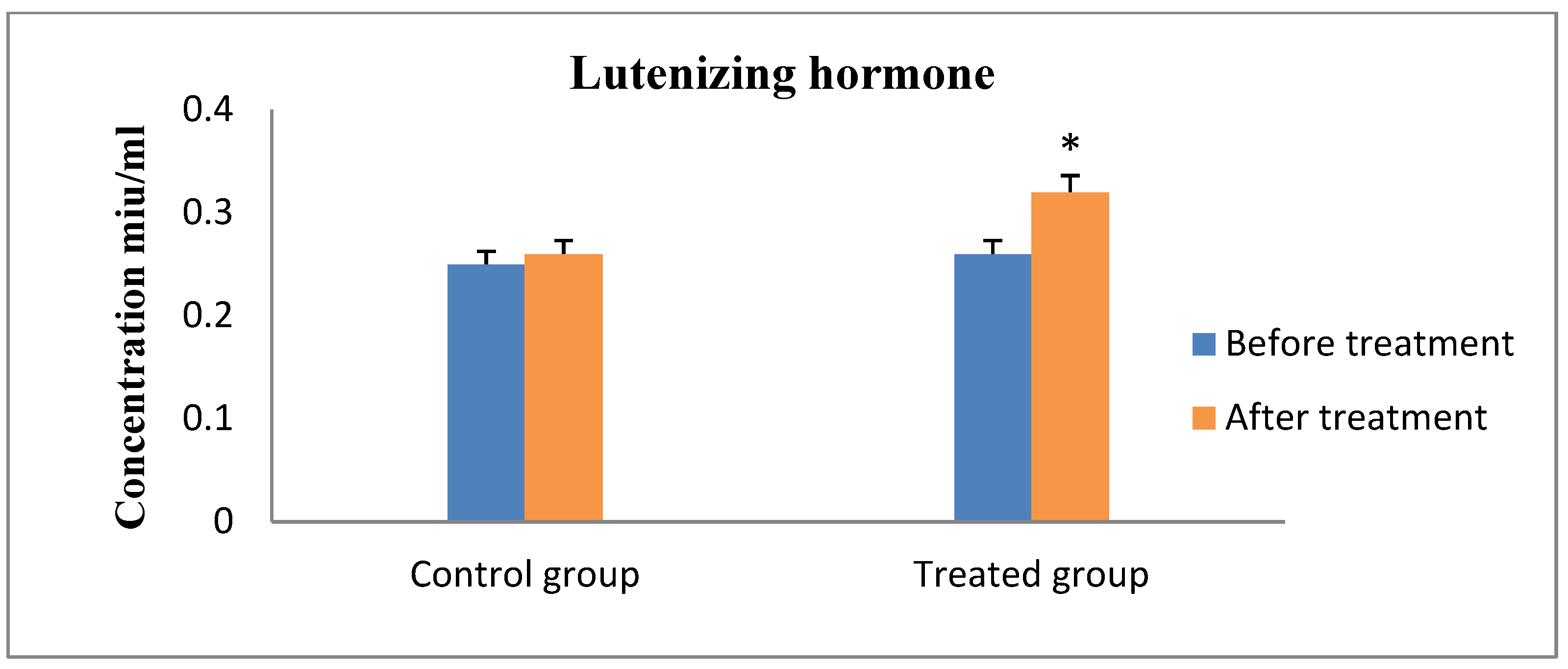
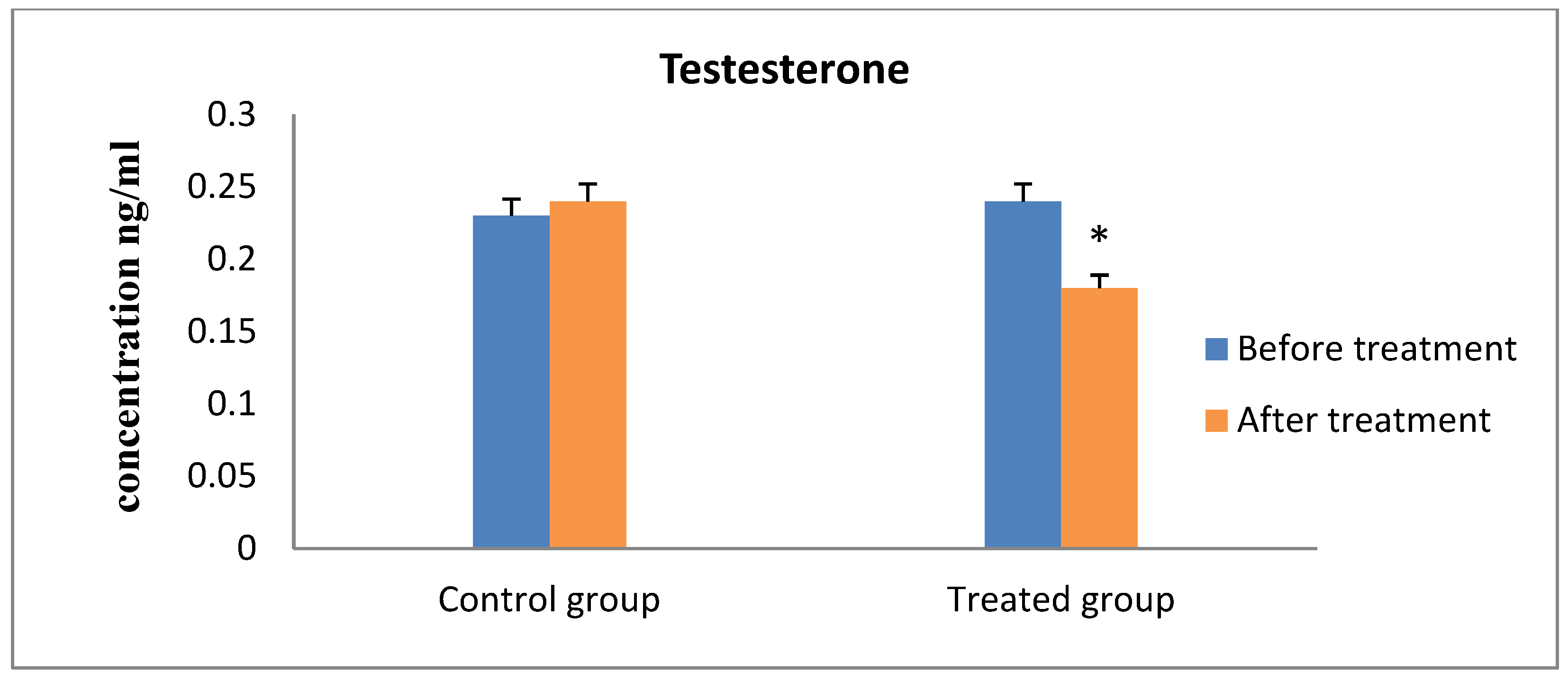
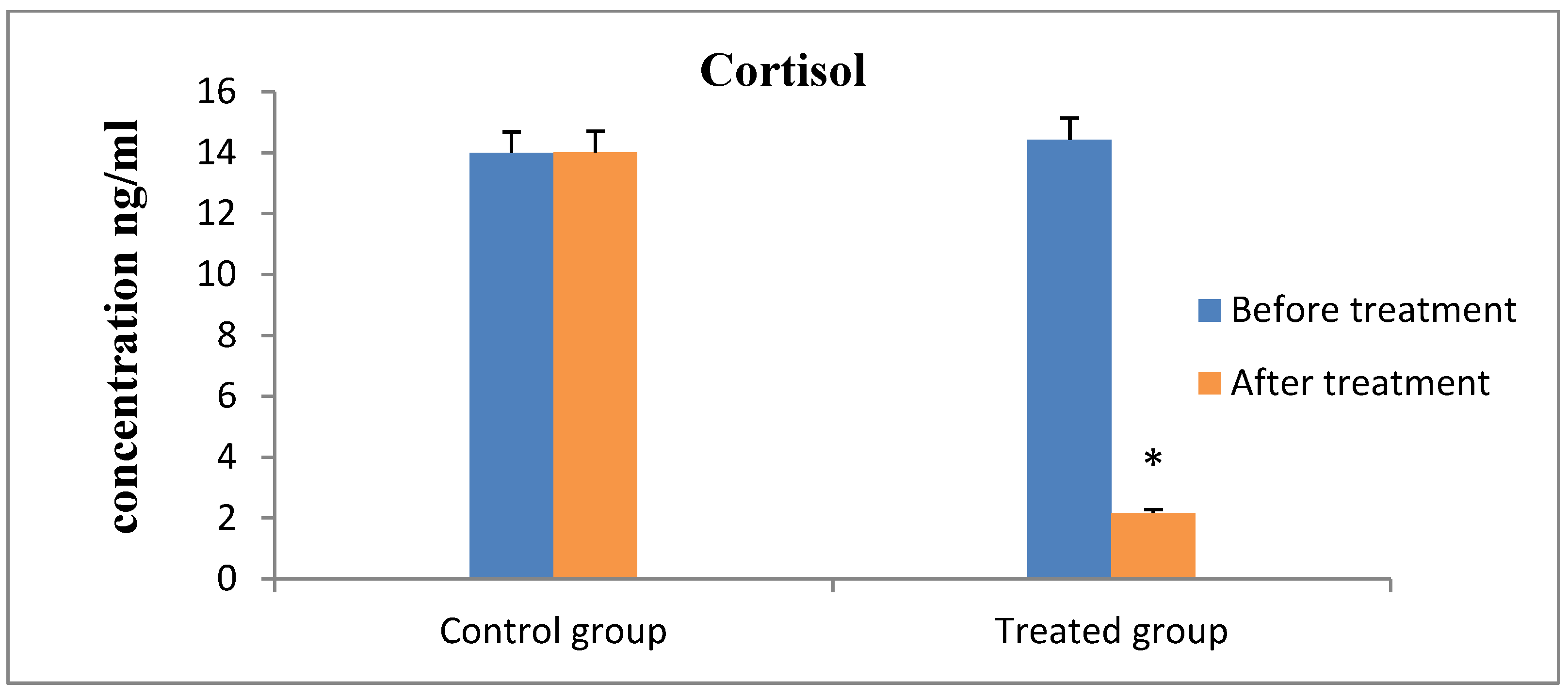
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borş, S.-I.; Borş, A. Ovarian cysts, an anovulatory condition in dairy cattle. J. Vet. Med. Sci. 2020, 82, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Cheng, J.; Yu, H.; Wang, Z.; Bai, Y.; Xia, C.; Xu, C. Early Warning for Ovarian Diseases Based on Plasma Non-esterified Fatty Acid and Calcium Concentrations in Dairy Cows. Front. Vet. Sci. 2021, 8, 792498. [Google Scholar] [CrossRef] [PubMed]
- Azarbayejani, R.; Mohammadsadegh, M. Glucose, insulin, and cortisol concentrations and glucose tolerance test in Holstein cows with inactive ovaries. Trop. Anim. Health Prod. 2021, 53, 41. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.M.; Gareis, N.C.; Hein, G.J.; Salvetti, N.R.; Amweg, A.N.; Huber, E.; Stassi, A.F.; Ortega, H.H.; Rey, F. Role of components of the insulin-like growth factor system in the early stages of ovarian follicular persistence in cattle. J. Comp. Pathol. 2017, 157, 201–214. [Google Scholar] [CrossRef]
- Zhao, C.; Bai, Y.; Fu, S.; Wu, L.; Xia, C.; Xu, C. Comparison of metabolic alterations in serum and milk whey between inactive ovaries and estrus dairy cows. Front. Vet. Sci. 2021, 7, 609391. [Google Scholar] [CrossRef]
- Xu, C.; Xia, C.; Sun, Y.; Xiao, X.; Wang, G.; Fan, Z.; Shu, S.; Zhang, H.; Xu, C.; Yang, W. Metabolic profiles using 1H-nuclear magnetic resonance spectroscopy in postpartum dairy cows with ovarian inactivity. Theriogenology 2016, 86, 1475–1481. [Google Scholar] [CrossRef]
- Rhodes, F.; McDougall, S.; Burke, C.; Verkerk, G.; Macmillan, K. Invited review: Treatment of cows with an extended postpartum anestrous interval. J. Dairy Sci. 2003, 86, 1876–1894. [Google Scholar] [CrossRef]
- Santos, J.; Juchem, S.; Cerri, R.; Galvao, K.; Chebel, R.; Thatcher, W.; Dei, C.; Bilby, C. Effect of bST and reproductive management on reproductive performance of Holstein dairy cows. J. Dairy Sci. 2004, 87, 868–881. [Google Scholar] [CrossRef]
- Peter, A.; Vos, P.; Ambrose, D. Postpartum anestrus in dairy cattle. Theriogenology 2009, 71, 1333–1342. [Google Scholar] [CrossRef]
- Mohammadsadegh, M. The impacts of eCG administration, 3 days before OVSYNCH on the treatment of inactive ovary of dairy cows. Rev. Médecine Vétérinaire 2019, 170, 110–116. [Google Scholar]
- Amin, Y.; Yousef, N.; Mahmoud, A.; Salah, M.; Khalil, A.; Shanab, O.; Hassaneen, A. Impact of polyherbal formulation oral administration on the estrus response, luteal activity, and oxidative stress in postpartum dairy cows with ovarian subfunction. Vet. World 2022, 15, 360–367. [Google Scholar] [CrossRef]
- Amin, Y.A.; El-Naga, E.M.A.; Noseer, E.A.; Fouad, S.S.; Ali, R.A. Synchronization with controlled internal drug release (CIDR) and prostaglandin F2α (PGF2α) influences oxidant/antioxidant biomarkers and mineral profile in summer-stressed anoestrous buffalo (Bubalus bubalis). Theriogenology 2019, 134, 34–41. [Google Scholar] [CrossRef]
- Amin, Y.; Said, A. The addition of chitosan to GnRH analog induces ovarian resumption and improves conception rates in buffaloes. Trop. Anim. Sci. J. 2021, 44, 1–9. [Google Scholar] [CrossRef]
- Mohammadzadeh, S.; Moradian, F.; Yeganeh, S.; Falahatkar, B.; Milla, S. Design, production and purification of a novel recombinant gonadotropin-releasing hormone associated peptide as a spawning inducing agent for fish. Protein Expr. Purif. 2020, 166, 105510. [Google Scholar] [CrossRef]
- Schneider, F.; Tomek, W.; Gründker, C. Gonadotropin-releasing hormone (GnRH) and its natural analogues: A review. Theriogenology 2006, 66, 691–709. [Google Scholar] [CrossRef]
- Ramakrishnappa, N.; Rajamahendran, R.; Lin, Y.-M.; Leung, P. GnRH in non-hypothalamic reproductive tissues. Anim. Reprod. Sci. 2005, 88, 95–113. [Google Scholar] [CrossRef]
- Ramakrishnappa, N.; Merwe, G.K.V.D.; Rajamahendran, R. Gonadotropin-releasing hormone receptor messenger ribonucleic acid expression in bovine ovary. (Abstract). Biol. Reprod. 2001, 64, 229. [Google Scholar]
- Dolan, S.; Evans, N.P.; Richter, T.A.; Nolan, A.M. Expression of gonadotropin-releasing hormone and gonadotropin-releasing hormone receptor in sheep spinal cord. Neurosci. Lett. 2003, 346, 120–122. [Google Scholar] [CrossRef]
- Annalisa, R.; Debora, C.; Maddalena, M.; Giuseppe, M.; Massimo, S.; Luigi, S.R. Epidural vs. intramuscular administration of lecirelin, a GnRH analogue, for the resolution of follicular cysts in dairy cows. Anim. Reprod. Sci. 2011, 126, 19–22. [Google Scholar] [CrossRef]
- Thurmon, J.C.; Tranquilli, W.J.; Benson, G.J.; Lumb, W.V. Lumb & Jones’ Veterinary Anesthesia, 3rd ed.; Williams & Wilkins: Wantage, UK; Lea & Febiger: Baltimore, MD, USA, 1996; pp. 494–495. [Google Scholar]
- Dissen, G.A.; Dees, W.L.; Ojeda, S.R. Neural and neurotrophic control of ovarian development. In The Ovary; Adashi, E.Y., Leung, P.C.K., Eds.; Raven Press: New York, NY, USA, 1993; pp. 1–19. [Google Scholar]
- Pelagalli, G.V.; Botte, V. Anatomia Veterinaria Sistematica e Comparata; Edi-Ermes: Milan, Italy, 2003. [Google Scholar]
- Ahmed, W. Overview on some factors negatively affecting ovarian activity in large farm animals. Glob. Vet 2007, 1, 53–66. [Google Scholar]
- Peters, A.E.; Mihalas, B.P.; Bromfield, E.G.; Roman, S.D.; Nixon, B.; Sutherland, J.M. Autophagy in female fertility: A role in oxidative stress and aging. Antioxid. Redox Signal. 2020, 32, 550–568. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Tiwari, M.; Pandey, A.N.; Shrivastav, T.G.; Chaube, S.K. Impact of stress on oocyte quality and reproductive outcome. J. Biomed. Sci. 2016, 23, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-N.; Zhang, C.-J.; Diao, H.-L.; Zhang, Y. Protective effects of curcumin against sodium arsenite-induced ovarian oxidative injury in a mouse model. Chin. Med. J. 2017, 130, 1026–1032. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xie, Y.; Yang, D.; Ren, D. Oxidative stress-induced apoptosis in granulosa cells involves JNK, p53 and Puma. Oncotarget 2017, 8, 25310. [Google Scholar] [CrossRef]
- Sohel, M.M.H.; Akyuz, B.; Konca, Y.; Arslan, K.; Sariozkan, S.; Cinar, M.U. Oxidative stress modulates the expression of apoptosis-associated microRNAs in bovine granulosa cells in vitro. Cell Tissue Res. 2019, 376, 295–308. [Google Scholar] [CrossRef]
- Cajas, Y.N.; Cañón-Beltrán, K.; Ladrón de Guevara, M.; Millán de la Blanca, M.G.; Ramos-Ibeas, P.; Gutiérrez-Adán, A.; Rizos, D.; González, E.M. Antioxidant nobiletin enhances oocyte maturation and subsequent embryo development and quality. Int. J. Mol. Sci. 2020, 21, 5340. [Google Scholar] [CrossRef]
- Edrey, Y.H.; Salmon, A.B. Revisiting an age-old question regarding oxidative stress. Free. Radic. Biol. Med. 2014, 71, 368–378. [Google Scholar] [CrossRef]
- Maclaran, K.; Nikolaou, D. Early ovarian ageing. Obstet. Gynaecol. 2019, 21, 107–116. [Google Scholar] [CrossRef]
- Yildiz, H. Serum Progesterone, Vitamin A, E, C and-Carotene Levels in Pregnant and Nonpregnant Cows Post-Mating. J. Anim. Vet. Adv. 2005, 4, 381–384. [Google Scholar]
- Steeneveld, W.; Amuta, P.; van Soest, F.J.; Jorritsma, R.; Hogeveen, H. Estimating the combined costs of clinical and subclinical ketosis in dairy cows. PloS ONE 2020, 15, e0230448. [Google Scholar] [CrossRef]
- Fiore, E.; Piccione, G.; Rizzo, M.; Morgante, M.; Barberio, A.; Giudice, E.; Gianesella, M. Adaptation of some energetic parameters during transition period in dairy cows. J. Appl. Anim. Res. 2018, 46, 402–405. [Google Scholar] [CrossRef]
- Xu, W.; Vervoort, J.; Saccenti, E.; Kemp, B.; van Hoeij, R.J.; van Knegsel, A.T. Relationship between energy balance and metabolic profiles in plasma and milk of dairy cows in early lactation. J. Dairy Sci. 2020, 103, 4795–4805. [Google Scholar] [CrossRef]
- Manspeaker, J.E.; Robl, M.G. The use of amino acid chelates in bovine fertility and embryonic viability. In The Roles of Amino Acid Chelates in Animal Nutrition; Noyes Publications: Westwood, NJ, USA, 1993; pp. 140–153. [Google Scholar]
- Campbell, M.; Miller, J.; Schrick, F. Effect of additional cobalt, copper, manganese, and zinc on reproduction and milk yield of lactating dairy cows receiving bovine somatotropin. J. Dairy Sci. 1999, 82, 1019–1025. [Google Scholar] [CrossRef]
- Roberts, T.; Chapinal, N.; LeBlanc, S.; Kelton, D.; Dubuc, J.; Duffield, T. Metabolic parameters in transition cows as indicators for early-lactation culling risk. J. Dairy Sci. 2012, 95, 3057–3063. [Google Scholar] [CrossRef]
- LeBlanc, S. Monitoring metabolic health of dairy cattle in the transition period. J. Reprod. Dev. 2010, 56, S29–S35. [Google Scholar] [CrossRef]
- Ferguson, J.D.; Galligan, D.T.; Thomsen, N. Principal descriptors of body condition score in Holstein cows. J. Dairy Sci. 1994, 77, 2695–2703. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Dairy Cattle: 2001, 7th ed.; The National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Bakr, A.; Ramoun, A. Effect of dietary energy on response of buffaloes with smooth inactive ovaries to gonadotrophin releasing hormone injection. Assiut Vet. Med. J. 2000, 42, 13–23. [Google Scholar]
- Ramoun, A.; Darwish, S. Ultrasound guided hormonal treatment of true anestrum in buffalo. Zag. Vet. J. 2006, 34, 51e62. [Google Scholar]
- Sakaguchi, K.; Yanagawa, Y.; Yoshioka, K.; Suda, T.; Katagiri, S.; Nagano, M. Relationships between the antral follicle count, steroidogenesis, and secretion of follicle-stimulating hormone and anti-Müllerian hormone during follicular growth in cattle. Reprod. Biol. Endocrinol. 2019, 17, 88. [Google Scholar] [CrossRef]
- Rose, M.P. Follicle stimulating hormone: International standards and reference preparations for the calibration of immunoassays and bioassays. Clin. Chim. Acta 1998, 273, 103–117. [Google Scholar] [CrossRef]
- Rebar, R.W.; Erickson, G.F.; Yen, S.S. Idiopathic premature ovarian failure: Clinical and endocrine characteristics. Fertil. Steril. 1982, 37, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Pickering, A.a.; Pottinger, T. Seasonal and diel changes in plasma cortisol levels of the brown trout, Salmo trutta L. Gen. Comp. Endocrinol. 1983, 49, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Kingston, H.; Jassie, L. Microwave acid sample decomposition for elemental analysis. J. Res. Natl. Bur. Stand. 1988, 93, 269. [Google Scholar] [CrossRef]
- Sakaguchi, K.; Suda, T.; Ninpetch, N.; Kawano, K.; Yanagawa, Y.; Katagiri, S.; Yoshioka, K.; Nagano, M. Plasma profile of follicle-stimulating hormone and sex steroid hormones after a single epidural administration of follicle-stimulating hormone via caudal vertebrae in Holstein dry cows. Anim. Sci. J. 2022, 93, e13696. [Google Scholar] [CrossRef]
- Monaco, D.; El-Bahrawy, K.A.; Lacalandra, G.M. Epidural administration of Lecirelin for inducing ovulation in female dromedary camels (Camelus dromedarius): A preliminary study. Small Rumin. Res. 2017, 146, 69–71. [Google Scholar] [CrossRef]
- Espey, L.L. Ovarian contractility and its relationship to ovulation: A review. Biol. Reprod. 1978, 19, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Niswender, G.D.; Juengel, J.L.; Silva, P.J.; Rollyson, M.K.; McIntush, E.W. Mechanisms controlling the function and life span of the corpus luteum. Physiol. Rev. 2000, 80, 1–29. [Google Scholar] [CrossRef]
- Morimoto, K.; Okamura, H.; Kanzaki, H.; Okuda, Y.; Takenaka, A.; Nishimura, T. The adrenergic nerve supply to bovine ovarian follicles. Int. J. Fertil. 1981, 26, 14–19. [Google Scholar]
- Gajewski, Z.; Thun, R.; Faundez, R.; Pawliński, B. The influence of alpha-adrenergic receptors stimulator and blockers and beta-blocker on the ovary and endocrinological activity in heifers during superovulation. J. Physiol. Pharmacol. 2006, 57, 173–188. [Google Scholar]
- Xu, C.; Roepke, T.A.; Zhang, C.; Rønnekleiv, O.K.; Kelly, M.J. Gonadotropin-releasing hormone (GnRH) activates the m-current in GnRH neurons: An autoregulatory negative feedback mechanism? Endocrinology 2008, 149, 2459–2466. [Google Scholar] [CrossRef]
- Skinner, D.C.; Albertson, A.J.; Navratil, A.; Smith, A.; Mignot, M.; Talbott, H.; Scanlan-Blake, N. Effects of gonadotrophin-releasing hormone outside the hypothalamic-pituitary-reproductive axis. J. Neuroendocrinol. 2009, 21, 282–292. [Google Scholar] [CrossRef]
- Viana, J.; Dorea, M.; Siqueira, L.; Arashiro, E.; Camargo, L.; Fernandes, C.; Palhão, M. Occurrence and characteristics of residual follicles formed after transvaginal ultrasound-guided follicle aspiration in cattle. Theriogenology 2013, 79, 267–273. [Google Scholar] [CrossRef]
- El-Razek, A.; Emad, M.; Allam, T.S. Some Biochemical Parameters and Hematological Picture in Cases of Smooth Inactive Ovaries in Buffalo-Cows. Alex. J. Vet. Sci. 2019, 61, 1. [Google Scholar]
- Ahmad, I.; Lodhi, L.; Qureshi, Z.; Younis, M. Studies on blood glucose, total proteins, urea and cholesterol levels in cyclic, non-cyclic and endometritic crossbred cows. Pak. Vet. J. 2004, 24, 92–94. [Google Scholar]
- Akhtar, M.S.; AsimFarooq, A.; AkbarLodhi, L.; Muhammad, S.; MazharAyaz, M.; Lashari, M.H.; Murtaza, S.; Hussain, I.; Irshad, M.; Hussain, M. Studies on serum macro and micro minerals status in repeat breeder and normal cyclic Nili-Ravi buffaloes and their treatment strategies. Afr. J. Biotechnol. 2014, 13, 1143–1146. [Google Scholar]
- Chumchai, R.; Ratsiri, T.; Ratchamak, R.; Vongpralub, T.; Boonkum, W.; Chankitisakul, V. Ovarian responses and FSH profiles at superovulation with a single epidural administration of gonadotropin in the Thai-Holstein crossbreed. Anim. Reprod. 2021, 18. [Google Scholar] [CrossRef]
- Taya, K. Inhibin and secretion of FSH in oestrous cycles of cows and pigs. J. Reprod. Fertil. 1991, 43, 151–162. [Google Scholar] [CrossRef]
- Saleh, N.; Mahmud, E.; Waded, E. Interactions between insulin like growth factor 1, thyroid hormones and blood energy metabolites in cattle with postpartum inactive ovaries. Nat. Sci. 2011, 9, 56–63. [Google Scholar]
- Jaiswal, R.; Singh, J.; Adams, G. Developmental pattern of small antral follicles in the bovine ovary. Biol. Reprod. 2004, 71, 1244–1251. [Google Scholar] [CrossRef]
- McNatty, K.P.; Juengel, J.L.; Pitman, J.L. Oocyte-somatic cell interactions and ovulation rate: Effects on oocyte quality and embryo yield. Reprod. Biol. Insights 2014, 7, 1–8. [Google Scholar]
- Webb, R.; England, B.G. Identification of the ovulatory follicle in the ewe: Associated changes in follicular size, thecal and granulosa cell luteinizing hormone receptors, antral fluid steroids, and circulating hormones during the preovulatory period. Endocrinology 1982, 110, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Mehlmann, L.M. Stops and starts in mammalian oocytes: Recent advances in understanding the regulation of meiotic arrest and oocyte maturation. Reproduction 2005, 130, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.D. Models of luteinization. Biol. Reprod. 2000, 63, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Spicer, L.; Alonso, J.; Chamberlain, C. Effects of thyroid hormones on bovine granulosa and thecal cell function in vitro: Dependence on insulin and gonadotropins. J. Dairy Sci. 2001, 84, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Dobson, H.; Ghuman, S.; Prabhakar, S.; Smith, R. A conceptual model of the influence of stress on female reproduction. Reprod. Camb. 2003, 125, 151–163. [Google Scholar] [CrossRef]
- Smith, R.; Ghuman, S.; Evans, N.; Karsch, F.; Dobson, H. Stress and the control of LH secretion in the ewe. Reprod. Camb. Suppl. 2003, 61, 267–282. [Google Scholar] [CrossRef]
- Constable, P.D.; Hinchcliff, K.W.; Done, S.H.; Grünberg, W. Veterinary Medicine: A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats, 11th ed.; Elsevier: Saint Louis, MO, USA, 2017; p. 2356. ISBN 2978-2350-7020-7057-2350. [Google Scholar]
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Sharma, R. Oxidative stress and its implications in female infertility–A clinician’s perspective. Reprod. Biomed. Online 2005, 11, 641–650. [Google Scholar] [CrossRef]
- Tiwari, M.; Prasad, S.; Tripathi, A.; Pandey, A.N.; Singh, A.K.; Shrivastav, T.G.; Chaube, S.K. Involvement of reactive oxygen species in meiotic cell cycle regulation and apoptosis in mammalian oocytes. React. Oxyg. Species 2016, 1, 110–116. [Google Scholar] [CrossRef]
- Brookes, P.S. Mitochondrial H+ leak and ROS generation: An odd couple. Free. Radic. Biol. Med. 2005, 38, 12–23. [Google Scholar] [CrossRef]
- Ghosh, M.; Gupta, M.; Kumar, R.; Kumar, S.; Balhara, A.; Singh, I. Relation between antioxidant status and postpartum anestrous condition in Murrah buffalo. Vet. World 2015, 8, 1163. [Google Scholar] [CrossRef]
- Amin, Y.A.; Mahmoudand, H.Y.; Megahed, G.A. Relation between Oxidant/Antioxidant status and postpartum anestrous Conditions. World J. Res. Rev. 2017, 9, 10. [Google Scholar]
- El-Amrawi, G.; El-Karim, D.G.; El-Sayed, Y. Antioxidant/oxidant status, metabolic profile status and concentration of some inflammatory cytokines in Egyptian buffalo-cows suffered from ovarian inactivity. Buffalo Bull. 2021, 40, 39–45. [Google Scholar]
- Jan, M.; Das, G.; Khan, F.; Singh, J.; Bashir, S.; Khan, S.; Prasad, J.; Mehrotra, S.; Pathak, M.; Singh, G. Evaluation of follicular oxidant-antioxidant balance and oxidative damage during reproductive acyclicity in water buffalo (Bubalus bubalis. Asian Pac. J. Reprod. 2014, 3, 35–40. [Google Scholar] [CrossRef]
- Cotgreave, I.A. Recent trends in glutathione biochemistry glutathione-protein interactions: A molecular link between oxidative stress and cell proliferation. Biochem. Biophys. Res. Commun. 1998, 242, 1–9. [Google Scholar] [CrossRef]
- Michalkova, V.; Benoit, J.B.; Attardo, G.M.; Medlock, J.; Aksoy, S. Amelioration of reproduction-associated oxidative stress in a viviparous insect is critical to prevent reproductive senescence. PLoS ONE 2014, 9, e87554. [Google Scholar] [CrossRef]
- Amin, B.Y.; Dar, R.R.; Ali, A.; Malla, J.A.; Shubeena, S. Role of micro-nutrients in bovine reproduction. Insight-Int. J. Reprod. All Anim. 2016, 6, 57–65. [Google Scholar] [CrossRef]
- Balamurugan, B.; Ramamoorthy, M.; Mandal, R.S.K.; Keerthana, J.; Gopalakrishnan, G.; Kavya, K.; Katiyar, R. Mineral an important nutrient for efficient reproductive health in dairy cattle. Int. J. Environ. Sci. Technol. 2017, 6, 694–701. [Google Scholar]
- Wang, Z.; Song, Y.; Sun, S.; Zhao, C.; Fu, S.; Xia, C.; Bai, Y. Metabolite Comparison between Serum and Follicular Fluid of Dairy Cows with Inactive Ovaries Postpartum. Animals 2022, 12, 285. [Google Scholar] [CrossRef]
- Karen, A.; Heleil, B.; Nasef, A.; Serur, B. Ovarian Inactivity in Dairy Cows: The Incidence, Causes and A Trial Of Treatment. Kafrelsheikh Vet. Med. J. 2010, 8, 1–21. [Google Scholar] [CrossRef]
- Virmani, M.; Malik, R.; Singh, P.; Dalal, S. Studies on blood biochemical and mineral profiles with the treatment of acyclicity in post-partum anestrus sahiwal cows. Haryana Vet. 2011, 50, 77–79. [Google Scholar]
- Singh, M.; Pant, H. Blood biochemical profile of normal and repeat breeder cows in Himachal Pradesh. Indian J. Anim. Reprod. 1998, 19, 156–157. [Google Scholar]
- Chaurasia, R.; Kushwaha, H.; Chaurasia, D.; Gendley, M.; Santra, A. Comparative studies of certain macro minerals during various reproductive states in buffaloes. Buffalo Bull. 2010, 29, 291–298. [Google Scholar]
- Sharad, K.; Atul, S. Comparative studies on metabolic profile of anestrous and normal cyclic Murrah buffaloes. Buffalo Bull. 2010, 29, 7–11. [Google Scholar]
- Devkota, B.; Nakao, T.; Kobayashi, K.; Sato, H.; Sah, S.K.; Singh, D.K.; Dhakal, I.P.; Yamagishi, N. Effects of treatment for anestrus in water buffaloes with PGF2α and GnRH in comparison with vitamin-mineral supplement, and some factors influencing treatment effects. J. Vet. Med. Sci. 2013, 75, 1623–1627. [Google Scholar] [CrossRef]
- Montiel, F.; Ahuja, C. Body condition and suckling as factors influencing the duration of postpartum anestrus in cattle: A review. Anim. Reprod. Sci. 2005, 85, 1–26. [Google Scholar] [CrossRef]
- Castro, N.; Kawashima, C.; Van Dorland, H.; Morel, I.; Miyamoto, A.; Bruckmaier, R. Metabolic and energy status during the dry period is crucial for the resumption of ovarian activity postpartum in dairy cows. J. Dairy Sci. 2012, 95, 5804–5812. [Google Scholar] [CrossRef]
- Andrieu, S. Is there a role for organic trace element supplements in transition cow health? Vet. J. 2008, 176, 77–83. [Google Scholar] [CrossRef]
- Kumar, S. Management of infertility due to mineral deficiency in dairy animals. In Proceedings of the ICAR Summer School on “Advance Diagnostic Techniques and Therapeutic Approaches to Metabolic and Deficiency Diseases in Dairy Animals”, Izatnagar, India, 15 July–4 August 2003; pp. 128–137. [Google Scholar]
- Derar, D.; Ali, A.; Almundarij, T.; Abd-Elmoniem, E.; Alhassun, T.; Zeitoun, M. Association between Serum Trace Elements Levels, Steroid Concentrations, and Reproductive Disorders in Ewes and Does. Vet. Med. Int. 2022, 2022, 8525089. [Google Scholar] [CrossRef]
- Keen, C.L.; Ensunsa, J.L.; Lönnerdal, B.A.; Zidenberg-Cherr, S. Manganese Encyclopedia of Human Nutrition, 3rd ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 148–154. [Google Scholar]
- Wilson, J.G. Bovine functional infertility in Devon and Cornwall: Response to manganese therapy. Vet. Rec. 1966, 79, 562–566. [Google Scholar]
- Corah, L. Trace mineral requirements of grazing cattle. Anim. Feed. Sci. Technol. 1996, 59, 61–70. [Google Scholar] [CrossRef]
- Patterson, H.; Adams, D.C.; Klopfenstein, T.J.; Clark, R.; Teichert, B. Supplementation to meet metabolizable protein requirements of primiparous beef heifers: II. Pregnancy and economics. J. Anim. Sci. 2003, 81, 563–570. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | No. of Cows | Estrus | Pregnant Cows after 1st Serving | Pregnant Cows after 2nd Serving | Total Pregnancies |
|---|---|---|---|---|---|
| Treated group | 30 | 26 (86%) | 18 (69%) | 5 (63%) | 23 (88%) |
| Control group | 30 | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Parameters | Groups | Before Treatment | After Treatment |
|---|---|---|---|
| Malondialdehyde (nmol) | Treated group | 4.8 ± 0.12 | 5.2 ± 0.11 |
| Control group | 4.7 ± 0.11 | 5.1 ± 0.13 | |
| Super Oxide Dismutase (U/mL) | Treated group | 386.5 ± 0.1 | 401.7 ± 0.12 |
| Control group | 387.5 ± 0.13 | 400.1 ± 0.14 | |
| Glutathione peroxidase (U/L) | Treated group | 1335 ± 0.11 | 1382 ± 0.15 * |
| Control group | 1336 ± 0.12 | 1337 ± 0.14 | |
| Total Antioxidant Capacity (mM) | Treated group | 3.6 ± 0.23 | 4.6 ± 0.25 * |
| Control group | 3.5 ± 0.21 | 3.7 ± 0.26 |
| Parameters | Groups | Before Treatment | After Treatment |
|---|---|---|---|
| Calcium (mg/dL) | Treated group | 4.73 ± 0.07 | 3.64 ± 0.12 |
| Control group | 4.51 ± 0.19 | 4.22 ± 0.17 | |
| Phosphorus (mg/dL) | Treated group | 10.20 ± 0.12 | 9.51 ± 0.11 |
| Control group | 10.21 ± 0.23 | 10.01 ± 0.25 | |
| Manganese (μg/L) | Treated group | 3.22 ± 0.24 | 3.61 ± 0.23 |
| Control group | 3.11 ± 0.16 | 3.22 ± 0.12 | |
| Magnesium (mg/dL) | Treated group | 1.43 ± 0.09 | 2.23 ± 0.08 * |
| Control group | 1.44 ± 0.01 | 1.45 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amin, Y.A.; Mahmoud, A.E.Z.; Ali, R.A.; Fouad, S.S.; Shanab, O.; Ibrahim, R.M.; Farrag, F.; Shukry, M.; Ibrahim, S.F.; Fericean, L.; et al. Treatment of Inactive Ovaries of Holstein Dairy Cows by Epidural Injection of GnRH Analogue (Receptal) and Its Impact on the Reproductive Hormones, Oxidant/Antioxidant Profile and Micro and Macro-Elements Profile. Animals 2023, 13, 653. https://doi.org/10.3390/ani13040653
Amin YA, Mahmoud AEZ, Ali RA, Fouad SS, Shanab O, Ibrahim RM, Farrag F, Shukry M, Ibrahim SF, Fericean L, et al. Treatment of Inactive Ovaries of Holstein Dairy Cows by Epidural Injection of GnRH Analogue (Receptal) and Its Impact on the Reproductive Hormones, Oxidant/Antioxidant Profile and Micro and Macro-Elements Profile. Animals. 2023; 13(4):653. https://doi.org/10.3390/ani13040653
Chicago/Turabian StyleAmin, Yahia A., Alaa Eldin Z. Mahmoud, Rana A. Ali, Samer S. Fouad, Obeid Shanab, Rawia M. Ibrahim, Foad Farrag, Mustafa Shukry, Samah F. Ibrahim, Liana Fericean, and et al. 2023. "Treatment of Inactive Ovaries of Holstein Dairy Cows by Epidural Injection of GnRH Analogue (Receptal) and Its Impact on the Reproductive Hormones, Oxidant/Antioxidant Profile and Micro and Macro-Elements Profile" Animals 13, no. 4: 653. https://doi.org/10.3390/ani13040653