
Anyone Can Get Old—All You Have to Do Is Live Long Enough: Understanding Mortality and Life Expectancy in European Hedgehogs (Erinaceus europaeus)



Abstract
:Simple Summary
Abstract
1. Introduction
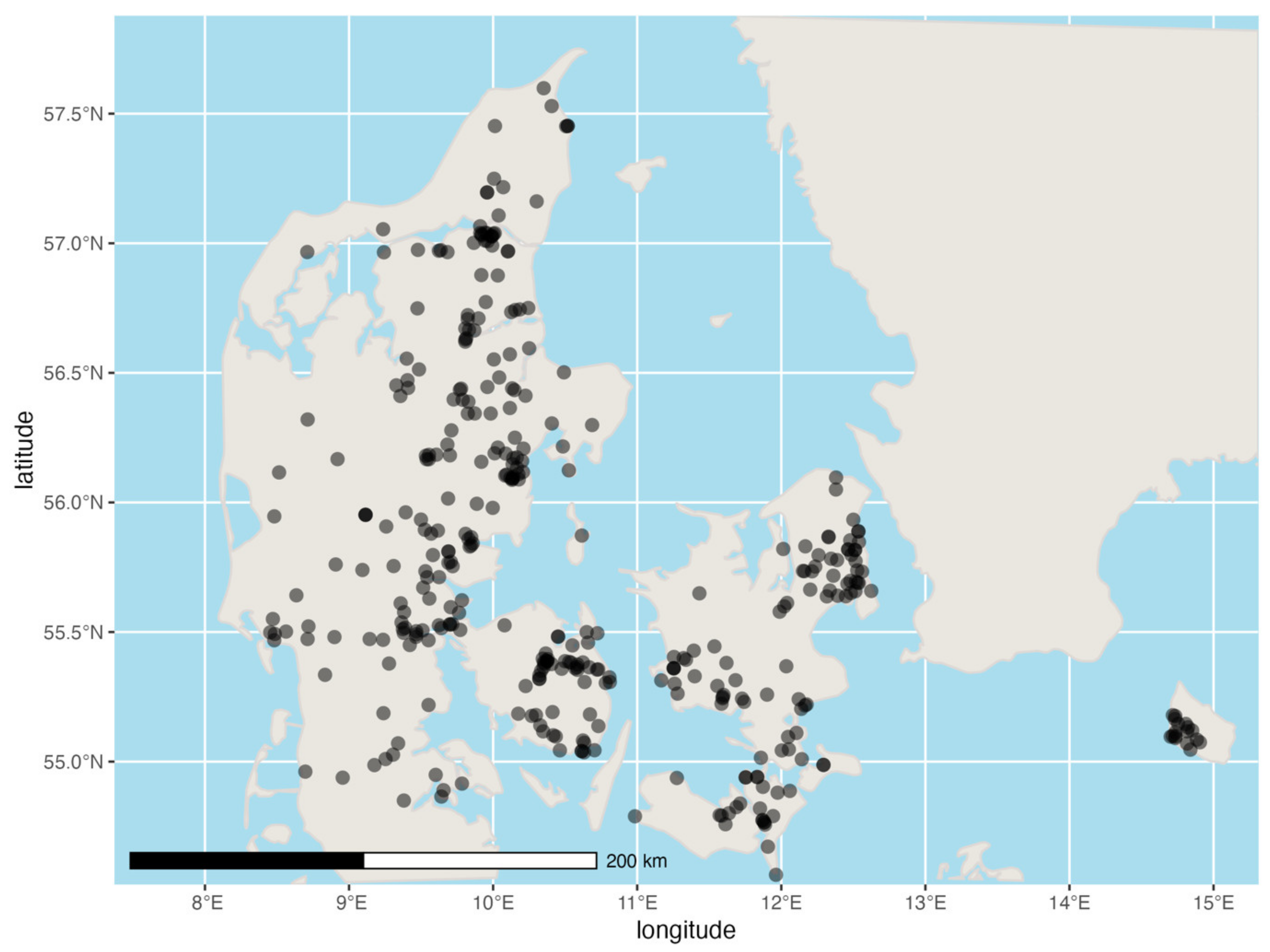
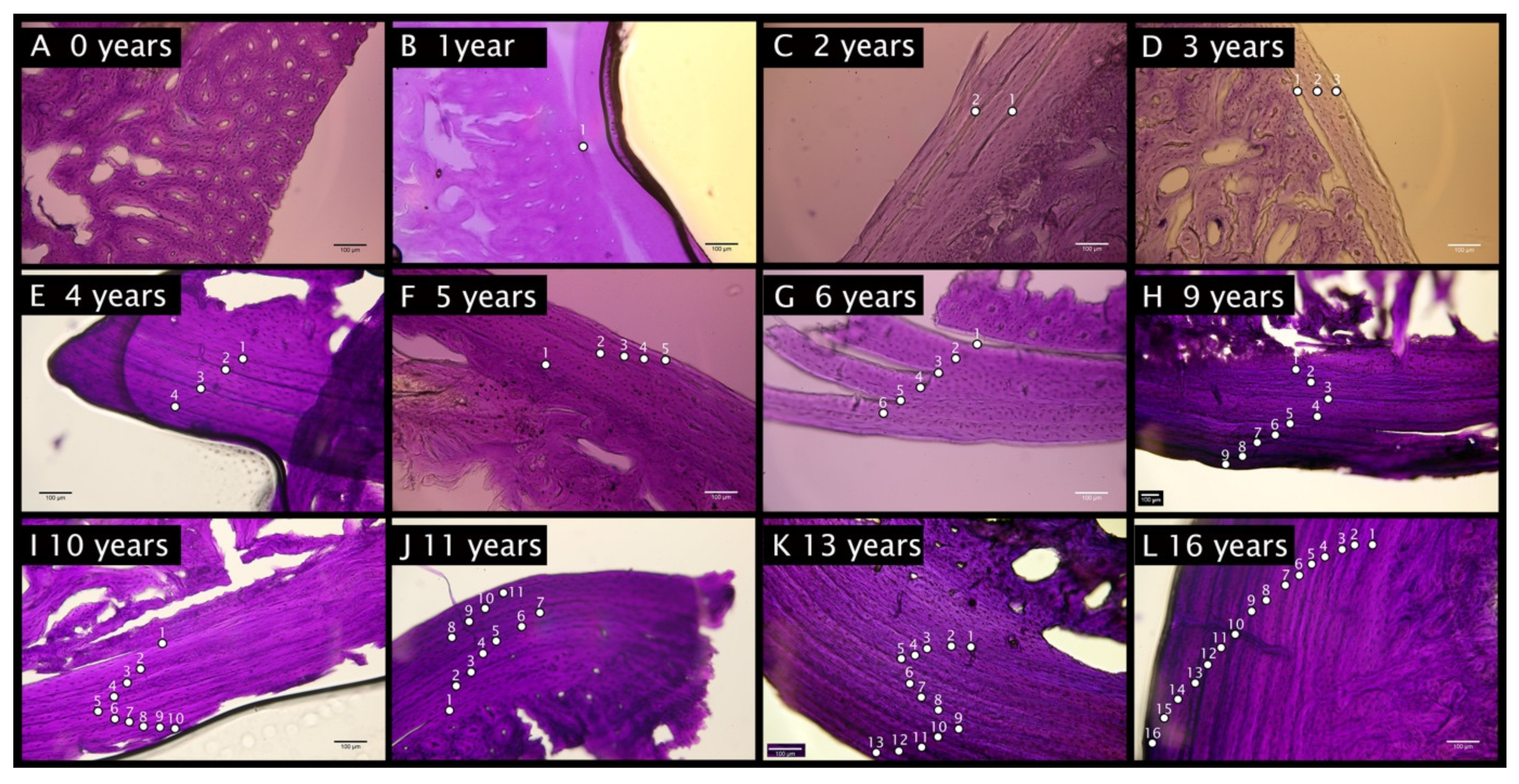
2. Materials and Methods
3. Results
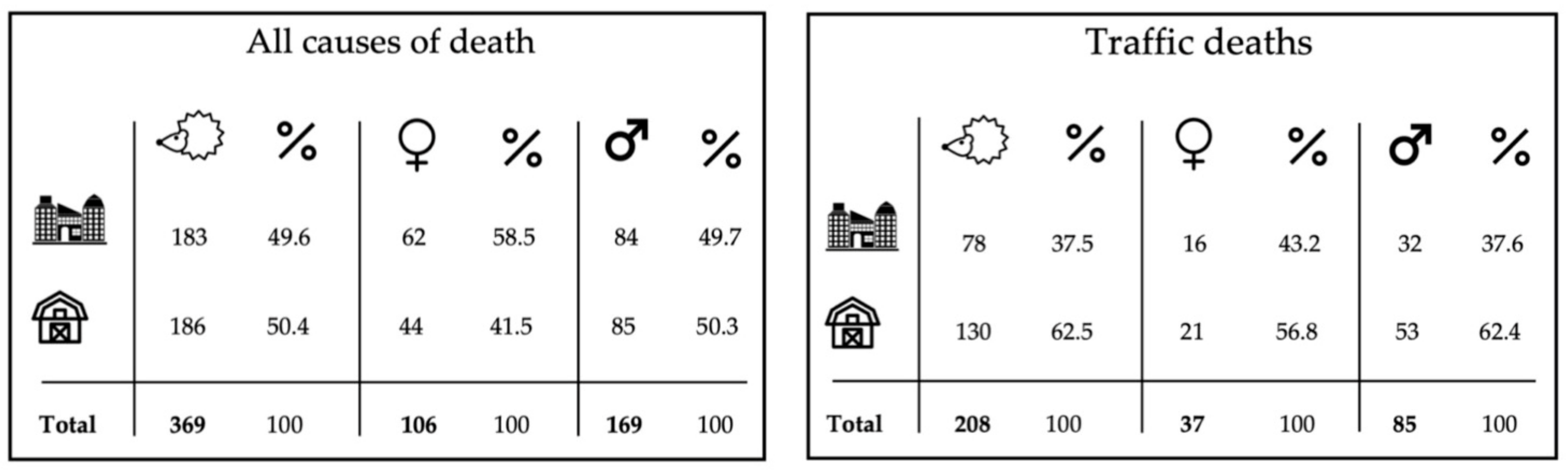
3.1. Timing and Cause of Death
3.2. The Influence of Habitat Type (Urban/Rural) and Sex on the Amount of Road-Killed Individuals
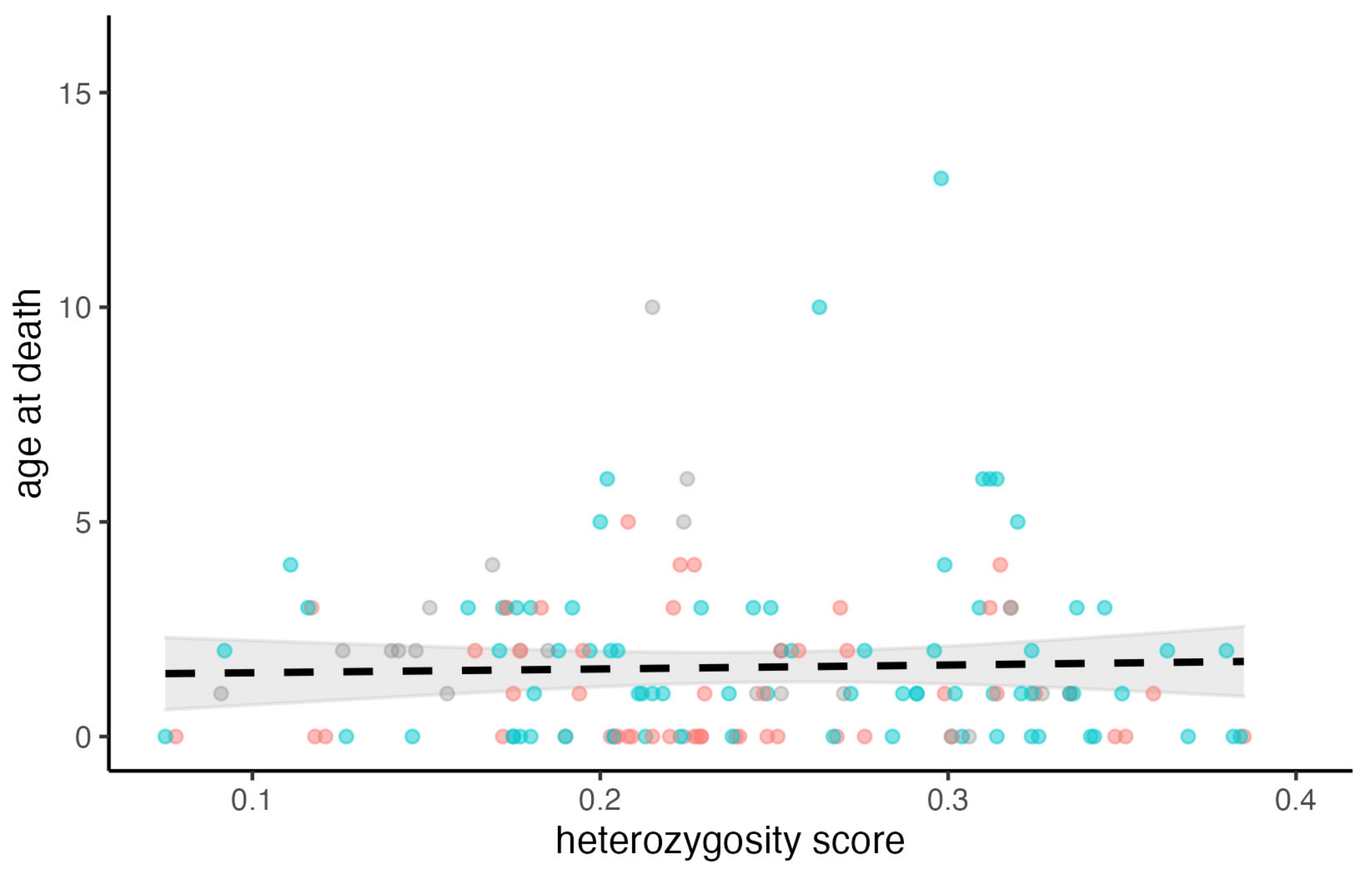
3.3. The Effects of Genetic Heterozygosity on Age-at-Death
3.4. Exploring the Association between Age-at-Death and Degree of Inbreeding, Cause of Death (Traffic/Non-Traffic), and the Interaction between Them
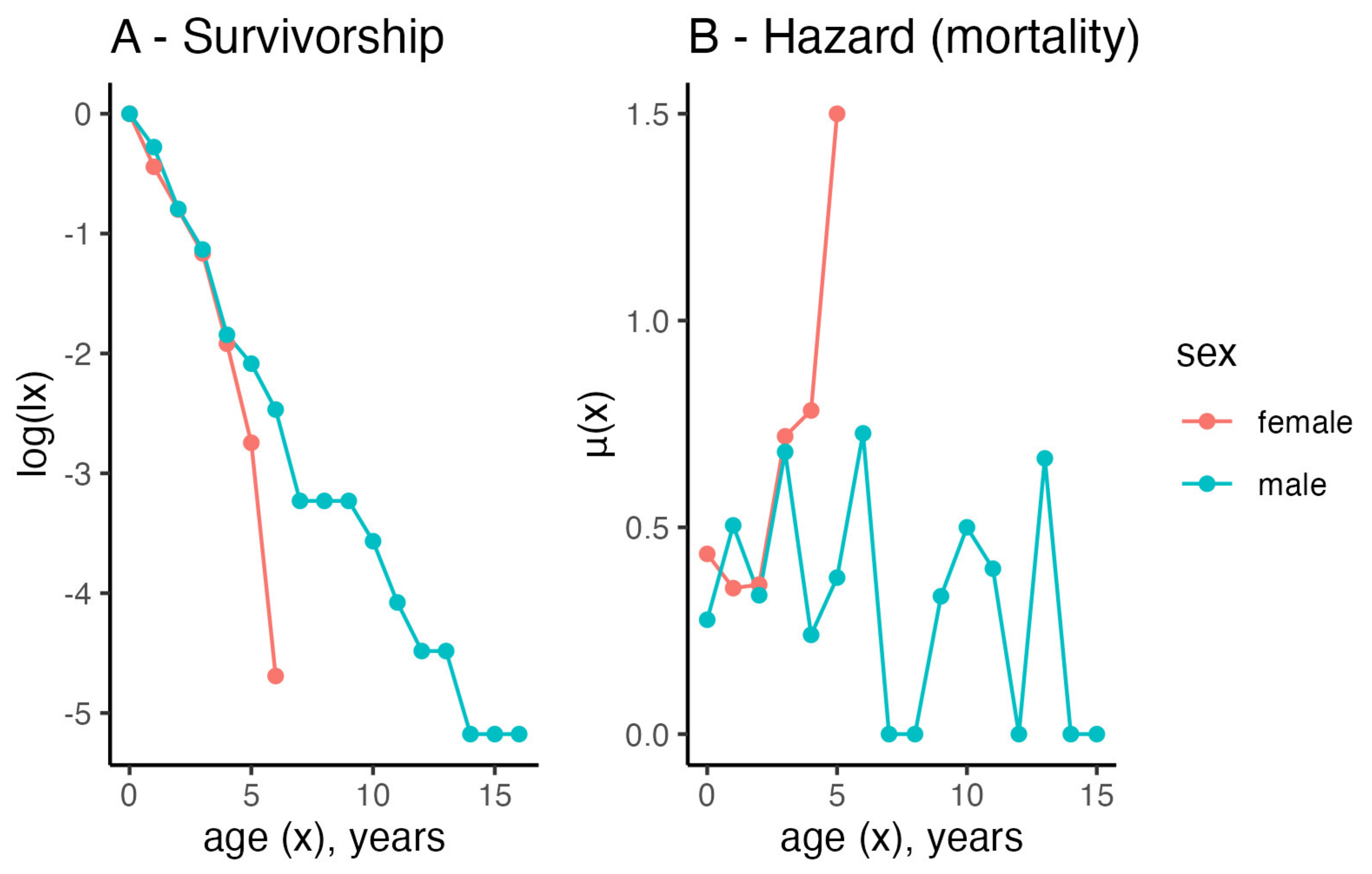
3.5. Life Table Analyses
4. Discussion
4.1. Periosteal Growth Lines
4.2. Timing and Cause of Death
4.3. Testing the Influence of Habitat Type (Urban/Rural) and Sex on the Number of Road-Killed Individuals and Using Data on Road-Killed Individuals for Population-Level Research
4.4. The Effects of Genetic Heterozygosity on Longevity
4.5. Exploring the Association between Age-at-Death and Degree of Inbreeding, Cause of Death (Traffic/Non-Traffic), and the Interaction between Them
4.6. Life Table Analyses and the Life Expectancy of Females and Males
4.7. Life Expectancy, Risks, and the Effects on Breeding
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roff, D.A. Life History Evolution; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Roff, D. Evolution of Life Histories: Theory and Analysis; Chapman and Hall: New York, NY, USA, 1992. [Google Scholar]
- Reznick, D.A.; Bryga, H.; Endler, J.A. Experimentally induced life-history evolution in a natural population. Nature 1990, 346, 357–359. [Google Scholar] [CrossRef]
- Neal, D. Introduction to Population Biology; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Spencer, W.; Rustigian-Romsos, H.; Strittholt, J.; Scheller, R.; Zielinski, W.; Truex, R. Using occupancy and population models to assess habitat conservation opportunities for an isolated carnivore population. Biol. Conserv. 2011, 144, 788–803. [Google Scholar] [CrossRef]
- Tempel, D.J.; Peery, M.; Gutierrez, R.J. Using integrated population models to improve conservation monitoring: California spotted owls as a case study. Ecol. Model. 2014, 289, 86–95. [Google Scholar] [CrossRef]
- Sierra Forest Legacy. Pacific fisher (Pekania pennanti). Available online: https://www.sierraforestlegacy.org/FC_SierraNevadaWildlifeRisk/PacificFisher.php (accessed on 10 January 2023).
- Sierra Forest Legacy. California Spotted Owl (Strix occidentalis occidentalis). Available online: https://www.sierraforestlegacy.org/FC_SierraNevadaWildlifeRisk/CaliforniaSpottedOwl.php (accessed on 10 January 2023).
- Morris, P. Hedgehogs; Whittet Books Ltd.: Stansted, Essex, UK, 2014. [Google Scholar]
- Reeve, N. Hedgehogs; Poyser: London, UK, 1994. [Google Scholar]
- Müller, F. Langzeit-Monitoring der Strassenverkehrsopfer beim Igel (Erinaceus europaeus L.) zur Indikation von Populationsdichteveränderungen entlang zweier Teststrecken im Landkreis Fulda. Beiträge Nat. Osthess. 2018, 54, 21–26. [Google Scholar]
- SoBH. The state of Britain’s Hedgehogs 2011; British Trust for Ornithology (BTO) commissioned by People’s Trust for Endangered Species (PTES) and the British Hedgehog Preservation Society (BHPS): London, UK, 2011. [Google Scholar]
- SoBH. The State of Britain’s Hedgehogs 2015; British Hedgehog Preservation Society and People’s Trust for Endangered Species: London, UK, 2015. [Google Scholar]
- SoBH. The State of Britain’s Hedgehogs 2018; British Hedgehog Preservation Society and People’s Trust for Endangered Species: London, UK, 2018. [Google Scholar]
- Hof, A.R.; Bright, P.W. Quantifying the long-term decline of the West European hedgehog in England by subsampling citizen-science datasets. Eur. J. Wildl. Res. 2016, 62, 407–413. [Google Scholar] [CrossRef]
- Krange, M. Change in the occurrence of the West European Hedgehog (Erinaceus europaeus) in western Sweden during 1950–2010. Ph.D. Thesis, Karlstad University, Karlstad, Sweden, 2015. [Google Scholar]
- van de Poel, J.L.; Dekker, J.; van Langevelde, F. Dutch hedgehogs Erinaceus europaeus are nowadays mainly found in urban areas, possibly due to the negative effects of badgers Meles meles. Wildl. Biol. 2015, 21, 51–55. [Google Scholar] [CrossRef]
- Williams, B.M.; Baker, P.J.; Thomas, E.; Wilson, G.; Judge, J.; Yarnell, R.W. Reduced occupancy of hedgehogs (Erinaceus europaeus) in rural England and Wales: The influence of habitat and an asymmetric intra-guild predator. Sci. Rep. 2018, 8, 12156. [Google Scholar] [CrossRef]
- Taucher, A.L.; Gloor, S.; Dietrich, A.; Geiger, M.; Hegglin, D.; Bontadina, F. Decline in Distribution and Abundance: Urban Hedgehogs under Pressure. Animals 2020, 10, 1606. [Google Scholar] [CrossRef]
- Wembridge, D.; Johnson, G.; Al-Fulaij, N.; Langton, S. The State of Britain’s Hedgehogs 2022; British Hedgehog Preservation Society and People’s Trust for Endangered Species: London, UK, 2022. [Google Scholar]
- Mathews, F.; Harrower, C. IUCN—Compliant Red List for Britain’s Terrestrial Mammals. Assessment by the Mammal Society under contract to Natural England, Natural Resources Wales and Scottish Natural Heritage; Natural England: Peterborough, UK.
- Brakes, C.R.; Smith, R.H. Exposure of non-target small mammals to rodenticides: Short-term effects, recovery and implications for secondary poisoning. J. Appl. Ecol. 2005, 42, 118–128. [Google Scholar] [CrossRef]
- Haigh, A.; O’Riordan, R.M.; Butler, F. Nesting behaviour and seasonal body mass changes in a rural Irish population of the Western hedgehog (Erinaceus europaeus). Acta Theriol. 2012, 57, 321–331. [Google Scholar] [CrossRef]
- Hof, A.R.; Bright, P.W. The value of agri-environment schemes for macro-invertebrate feeders: Hedgehogs on arable farms in Britain. Anim. Conserv. 2010, 13, 467–473. [Google Scholar] [CrossRef]
- Huijser, M.P.; Bergers, P.J.M. The effect of roads and traffic on hedgehog (Erinaceus europaeus) populations. Biol. Conserv. 2000, 95, 111–116. [Google Scholar] [CrossRef]
- Young, R.P.; Davison, J.; Trewby, I.D.; Wilson, G.J.; Delahay, R.J.; Doncaster, C.P. Abundance of hedgehogs (Erinaceus europaeus) in relation to the density and distribution of badgers (Meles meles). J. Zool. 2006, 269, 349–356. [Google Scholar] [CrossRef]
- Hubert, P.; Julliard, R.; Biagianti, S.; Poulle, M.-L. Ecological factors driving the higher hedgehog (Erinaceus europeaus) density in an urban area compared to the adjacent rural area. Landsc. Urban Plan. 2011, 103, 34–43. [Google Scholar] [CrossRef]
- Pettett, C.E.; Moorhouse, T.P.; Johnson, P.J.; Macdonald, D.W. Factors affecting hedgehog (Erinaceus europaeus) attraction to rural villages in arable landscapes. Eur. J. Wildl. Res. 2017, 63, 54. [Google Scholar] [CrossRef]
- Dowding, C.V.; Harris, S.; Poulton, S.; Baker, P.J. Nocturnal ranging behaviour of urban hedgehogs, Erinaceus europaeus, in relation to risk and reward. Anim. Behav. 2010, 80, 13–21. [Google Scholar] [CrossRef]
- Dowding, C.V.; Shore, R.F.; Worgan, A.; Baker, P.J.; Harris, S. Accumulation of anticoagulant rodenticides in a non-target insectivore, the European hedgehog (Erinaceus europaeus). Environ. Pollut. 2010, 158, 161–166. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Nielsen, J.L.; Jones, O.R.; Berg, T.B.; Pertoldi, C. Genetic structure of the European hedgehog (Erinaceus europaeus) in Denmark. PLoS ONE 2020, 15, e0227205. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Yashiro, E.; Sverrisdóttir, E.; Nielsen, K.L.; Lukassen, M.B.; Nielsen, J.L.; Asp, T.; Pertoldi, C. Applying the GBS Technique for the Genomic Characterization of a Danish Population of European Hedgehogs (Erinaceus europaeus). Genet. Biodivers. (GABJ) 2019, 3, 78–86. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Berg, T.B.; Dabelsteen, T.; Jones, O.R. The ecology of suburban juvenile European hedgehogs (Erinaceus europaeus) in Denmark. Ecol. Evol. 2019, 9, 13174–13187. [Google Scholar] [CrossRef] [PubMed]
- Burroughes, N.D.; Dowler, J.; Burroughes, G. Admission and Survival Trends in Hedgehogs Admitted to RSPCA Wildlife Rehabilitation Centres. Proc. Zool. Soc. 2021, 74, 198–204. [Google Scholar] [CrossRef]
- Lukešová, G.; Voslarova, E.; Vecerek, V.; Vucinic, M. Trends in intake and outcomes for European hedgehog (Erinaceus europaeus) in the Czech rescue centers. PLoS ONE 2021, 16, e0248422. [Google Scholar] [CrossRef] [PubMed]
- Morris, P. Hedgehog; Collins: London, UK, 2018; pp. 1–404. [Google Scholar]
- Rasmussen, S.L.; Kalliokoski, O.; Dabelsteen, T.; Abelson, K. An exploratory investigation of glucocorticoids, personality and survival rates in wild and rehabilitated hedgehogs (Erinaceus europaeus) in Denmark. BMC Ecol. Evol. 2021, 21, 96. [Google Scholar] [CrossRef]
- Jensen, A.B. Overwintering of European hedgehogs Erinaceus europaeus in a Danish rural area. Acta Theriol. 2004, 49, 145–155. [Google Scholar] [CrossRef]
- Park, E.A. The imprinting of nutritional disturbances on the growing bone. Pediatrics 1964, 33, 815–862. [Google Scholar] [CrossRef]
- Peabody, F.E. Annual growth zones in living and fossil vertebrates. J. Morphol. 1961, 108, 11. [Google Scholar] [CrossRef]
- Morris, P.A. A method for determining absolute age in the hedgehog. J. Zool. 1970, 161, 277–281. [Google Scholar] [CrossRef]
- Reeve, N.J.; Love, B.; Shore, R. X-ray measures of long bones as an aid to age-determination in hedgehogs (Erinaceus europaeus). In Proceedings of the 1st European Hedgehog Research Group Meeting, Arendal, Norway; 1996. [Google Scholar]
- Ohtaishi, N.; Hachiya, N.; Shibata, Y. Age determination of the hare from annual layers in the mandibular bone. Acta Theriol. 1976, 21, 168–171. [Google Scholar] [CrossRef]
- Henderson, B.A.; Bowen, H.M. Estimating the age of the European rabbit, Oryctolagus Cuniculus, by counting the adhesion lines in the periosteal zone of the lower mandible. J. Appl. Ecol. 1979, 16, 393–396. [Google Scholar] [CrossRef]
- Castanet, J.; Croci, S.; Aujard, F.; Perret, M.; Cubo, J.; de Margerie, E. Lines of arrested growth in bone and age estimation in a small primate: Microcebus murinus. J. Zool. 2004, 263, 31–39. [Google Scholar] [CrossRef]
- Sander, P.M.; Andrássy, P. Lines of arrested growth and long bone histology in Pleistocene large mammals from Germany: What do they tell us about dinosaur physiology? Palaeontogr. Abt. A 2006, 277, 143–159. [Google Scholar] [CrossRef]
- Brockie, R.E. Ecology of the hedgehog Erinaceus europaeus in New Zealand. Master’s Thesis, Victoria University of Wellington, Wellington, New Zealand, 1958. [Google Scholar]
- Dickman, C.R. Age-related dietary change in the European hedgehog, Erinaceus europaeus. J. Zool. 1988, 215, 1–14. [Google Scholar] [CrossRef]
- Döpke, C. Kasuistische Auswertung der Untersuchungen von Igeln (Erinaceus europaeus) im Einsendungsmaterial des Instituts für Pathologie von 1980 bis 2001. Ph.D. Thesis, Tierärztliche Hochschule Hannover, Hannover, Germany, 2002. [Google Scholar]
- Haigh, A.; Kelly, M.; Butler, F.; O’Riordan, R.M. Non-invasive methods of separating hedgehog (Erinaceus europaeus) age classes and an investigation into the age structure of road kill. Acta Theriol. 2014, 59, 165–171. [Google Scholar] [CrossRef]
- Heddergott, M. Zur Altersstruktur des Igels Erinaceus europaeus (L., 1758) in Thüringen anhand von Schädeln (Mammalia: Insectivora, Erinaceidae). Veröffentlichungen Naturhist. Mus. Schleus. 2003, 18, 79–82. [Google Scholar]
- Heddergott, M. Age-structure of hedgehog Erinaceus europaeus (L., 1758) in the Harz Mountains. Abh. Und Ber. Aus Dem Mus. Heine. 2004, 6, 125–130. [Google Scholar]
- Heddergott, M. Age structure of the hedgehog Erinaceus europaeus L., 1758, in the district Saalfeld-Rudolstadt (Thuringia). Hercynia 2005, 38, 113–118. [Google Scholar]
- Heddergott, M.; Müller, F. Zur Altersstruktur zweier Populationen des Braunbrustigels Erinaceus europaeus L., 1758 (Mammalia: Insectivora) in Osthessen. Beiträge Zur Nat. Osthess. 2008, 45, 63–71. [Google Scholar]
- Heddergott, M.; Steinbach, O.; Heddergott, C. Zur Altersstruktur des Braunbrustigels Erinaceus europaeus (Linnaeus, 1758) im Stadtgebiet Heilbad Heiligenstadt (Thüringen) (Mammalia: Insectivora, Erinaceidae)*. Mauritiana 2010, 21, 231–239. [Google Scholar]
- Heyne, P. Beitrag zur Altersstruktur von Erinaceus europaeus L., 1758. Populationsökologie von Kleinsäugerarten, Martin-Luther-Universitaet Halle-Wittenberg Wissenschaftliche Beitrage. SLUB 1990, 1990, 49–54. [Google Scholar]
- Kratochvil, J. Hedgehogs of the genus Erinaceus in Czechoslovakia insectivora mammalia. Zool. Listy 1975, 24, 297–312. [Google Scholar]
- Kristoffersson, R. A note on the age distribution of hedgehogs Erinaceus europaeus in Finland. Ann. Zool. Fenn. 1971, 8, 554–557. [Google Scholar]
- Morris, P.A. Some Aspects of the Ecology of the Hedgehog (Erinaceus europaeus). University of London: London, UK, 1969. [Google Scholar]
- Neuschulz, N.; Schubert, M. Altersermittlung bei Erinaceus europaeus L., 1758 an einer Igelfütterung. Popul. Von Kleinsäugerarten Wiss. Beitr. Univ. Halle 1990, 34, 42. [Google Scholar]
- Rautio, A.; Kunnasranta, M.; Valtonen, A.; Ikonen, M.; Hyvarinen, H.; Holopainen, I.J.; Kukkonen, J.V.K. Sex, Age, and Tissue Specific Accumulation of Eight Metals, Arsenic, and Selenium in the European Hedgehog (Erinaceus europaeus). Arch. Environ. Contam. Toxicol. 2010, 59, 642–651. [Google Scholar] [CrossRef]
- Skoudlin, J. On determining the age in erinaceus-europaeus and erinaceus-concolor insectivora erinaceidae. Vestn. Ceskoslovenske Spol. Zool. 1976, 40, 300–306. [Google Scholar]
- Skoudlin, J. Age structure of czechoslovak populations of Erinaceus-Europaeus and Erinaceus-Concolor Insectivora Erinaceidae. Vestn. Ceskoslovenske Spol. Zool. 1981, 45, 307–313. [Google Scholar]
- Brockie, R.E. Studies of the Hedgehog Erinaceus europaeus L. in New Zealand; Victoria University of Wellington: Wellington, New Zealand, 1974. [Google Scholar]
- Parkes, J. Some aspects of the biology of the hedgehog erinaceus-europaeus in the manawatu new-zealand. New Zealand J. Zool. 1975, 2, 463–472. [Google Scholar] [CrossRef]
- Petrides, G.A. The determination of sex and age ratios in the cottontail rabbit. Am. Midl. Nat. 1951, 46, 312–336. [Google Scholar] [CrossRef]
- Morris, P.A. Epiphyseal fusion in the forefoot as a means of age determination in the hedgehog (Erinaceus europaeus) J. Zool. 1971, 164, 254–259. [Google Scholar] [CrossRef]
- Scott-Hayward, L. Data on age determination (pers. comm. p. 224). In Hedgehogs; Morris, P.A., Ed.; Collins: London, UK, 2018. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Hijmans, R.J.; raster: Geographic Data Analysis and Modeling. R package. Available online: https://cran.r-project.org/web/packages/raster/index.html (accessed on 2 September 2022).
- Rasmussen, S.L.; Larsen, J.; van Wijk, R.E.; Jones, O.R.; Berg, T.B.; Angen, O.; Larsen, A.R. European hedgehogs (Erinaceus europaeus) as a natural reservoir of methicillin-resistant Staphylococcus aureus carrying mecC in Denmark. PLoS ONE 2019, 14, e0222031. [Google Scholar] [CrossRef]
- Samuel Preston, P.H.; Guillot, M. Demography: Measuring and Modeling Population Processes; Wiley-Blackwell: Hoboken, NJ, USA, 2020. [Google Scholar]
- Jones, O.R. Life tables: Construction and interpretation In Demographic Methods Across the Tree of Life; Oxford University Press: Oxford, UK, 2021. [Google Scholar]
- Kulus, M.J.; Dąbrowski, P. How to calculate the age at formation of Harris lines? A step-by-step review of current methods and a proposal for modifications to Byers’ formulas. Archaeol. Anthropol. Sci. 2019, 11, 1169–1185. [Google Scholar] [CrossRef]
- Foster, N.J.; Maloney, R.F.; Recio, M.R.; Seddon, P.J.; van Heezik, Y. European hedgehogs rear young and enter hibernation in New Zealand’s alpine zones. New Zealand J. Ecol. 2021, 45, 1–6. [Google Scholar] [CrossRef]
- Walhovd, H. Winter activity of Danish hedgehogs in 1973-74 with information on the size of the animals observed and location of the recordings. Flora Og Fauna 1976, 82, 35–42. [Google Scholar]
- Walhovd, H. The overwintering pattern of Danish hedgehogs in outdoor confinement, during three successive winters. Nat. Jutl. 1978, 20, 273–284. [Google Scholar]
- Hansen, L. Trafikdræbte dyr i Danmark. Dan. Ornitol. Foren. Tidsskr. 1982, 76, 96–110. [Google Scholar]
- Holsbeek, L.; Rodts, J.; Muyldermans, S. Hedgehog and other animal traffic victims in Belgium: Results of a countrywide survey. Lutra 1999, 42, 111–119. [Google Scholar]
- Schwartz, A.L.; Shilling, F.M.; Perkins, S.E. The value of monitoring wildlife roadkill. Eur. J. Wildl. Res. 2020, 66, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wembridge, D.; Newman, M.R.; Bright, P.; Morris, P. An estimate of the annual number of hedgehog (Erinaceus europaeus) road casualties in Great Britain; Mammal Society: London, UK, 2016. [Google Scholar]
- Kent, E.; Schwartz, A.L.; Perkins, S.E. Life in the fast lane: Roadkill risk along an urban–rural gradient. J. Urban Ecol. 2021, 7, juaa039. [Google Scholar] [CrossRef]
- Raynaud, J.; Schradin, C. Experimental increase of testosterone increases boldness and decreases anxiety in male African striped mouse helpers. Physiol. Behav. 2014, 129, 57–63. [Google Scholar] [CrossRef]
- Morris, P.A. A study of home range and movements in the hedgehog (Erinaceus europaeus). J. Zool. 1988, 214, 433–449. [Google Scholar] [CrossRef]
- Reeve, N.J. The home range of the hedgehog as revealed by a radio tracking study. Symp. Zool. Soc. Lond. 1982, 49, 207–230. [Google Scholar]
- Kristiansson, H. Ecology of a hedgehog (Erinaceus europaeus) population in Southern Sweden. PhD Thesis, Department of Animal Ecology, University of Lund, Lund, Sweden, 1984. [Google Scholar]
- Haigh, A.; O’Riordan, R.M.; Butler, F. Hedgehog Erinaceus europaeus mortality on Irish roads. Wildl. Biol. 2014, 20, 155–160. [Google Scholar] [CrossRef]
- Göransson, G.; Karlsson, J.; Lindgren, A. Road mortality of the hedgehog Erinaceus europaeus in southern Sweden (Igelkotten och biltrafiken). Fauna Och Flora 1976, 71, 1–6. [Google Scholar]
- Huijser, M.P. Life on the Edge: Hedgehog Traffic Victims and Mitigation Strategies in an Anthropogenic Landscape; Wageningen University and Research: Wageningen, The Netherlands, 2000. [Google Scholar]
- Vejdirektoratet. Trafik på strækninger 2021. Available online: http://vej08.vd.dk/stroemkort/nytui/kort/Stroemkort.html?id=201 (accessed on 10 January 2023).
- Rondinini, C.; Doncaster, C.P. Roads as barriers to movement for hedgehogs. Funct. Ecol. 2002, 16, 504–509. [Google Scholar] [CrossRef]
- Brockie, R.E.; Sadleir, R.M.; Linklater, W.L. Long-term wildlife road-kill counts in New Zealand. New Zealand J. Zool. 2009, 36, 123–134. [Google Scholar] [CrossRef]
- Moore, L.J.; Petrovan, S.O.; Baker, P.J.; Bates, A.J.; Hicks, H.L.; Perkins, S.E.; Yarnell, R.W. Impacts and potential mitigation of road mortality for hedgehogs in Europe. Animals 2020, 10, 1523. [Google Scholar] [CrossRef]
- Slater, F.M. An assessment of wildlife road casulties—the potential discrepancy between numbers counted and numbers killed. Web Ecol. 2002, 3, 33–42. [Google Scholar] [CrossRef]
- Ratton, P.; Secco, H.; Da Rosa, C.A. Carcass permanency time and its implications to the roadkill data. Eur. J. Wildl. Res. 2014, 60, 543–546. [Google Scholar] [CrossRef]
- Schwartz, A.L.W.; Williams, H.F.; Chadwick, E.; Thomas, R.J.; Perkins, S.E. Roadkill scavenging behaviour in an urban environment. J. Urban Ecol. 2018, 4. [Google Scholar] [CrossRef]
- Keller, L.F.; Waller, D.M. Inbreeding effects in wild populations. Trends Ecol. Evol. 2002, 17, 230–241. [Google Scholar] [CrossRef]
- Wright, P.G. , Coomber, F. G., Bellamy, C. C., Perkins, S. E., Mathews, F. Predicting hedgehog mortality risks on British roads using habitat suitability modelling. PeerJ 2020, 7, e8154. [Google Scholar] [CrossRef]
- Austad, S.N. Why women live longer than men: Sex differences in longevity. Gend. Med. 2006, 3, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Clutton-Brock, T.H.; Isvaran, K. Sex Differences in Ageing in Natural Populations of Vertebrates. Proc. Biol. Sci. 2007, 274, 3097–3104. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R.L. Parental investment and sexual selection. In Sexual Selection and the Descent of Man; Campbell, B., Ed.; Aldine-Atherton: Chicago, IL, USA, 1972; pp. 136–179. [Google Scholar]
- Beirne, C.; Delahay, R.; Young, A. Sex differences in senescence: The role of intra-sexual competition in early adulthood. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151086. [Google Scholar] [CrossRef]
- Dugdale, H.L.; Macdonald, D.W.; Pope, L.C.; Burke, T. Polygynandry, extra-group paternity and multiple-paternity litters in European badger (Meles meles) social groups. Mol. Ecol. 2007, 16, 5294–5306. [Google Scholar] [CrossRef]
- Delahay, R.; Walker, N.; Forrester, G.; Harmsen, B.; Riordan, P.; Macdonald, D.; Newman, C.; Cheeseman, C. Demographic correlates of bite wounding in Eurasian badgers, Meles meles L., in stable and perturbed populations. Anim. Behav. 2006, 71, 1047–1055. [Google Scholar] [CrossRef]
- Macdonald, D.W.; Newman, C.; Nouvellet, P.M.; Buesching, C.D. An Analysis of Eurasian Badger (Meles meles) Population Dynamics: Implications for Regulatory Mechanisms. J. Mammal. 2009, 90, 1392–1403. [Google Scholar] [CrossRef]
- Sugianto, N.A.; Newman, C.; Macdonald, D.W.; Buesching, C.D. Reproductive and somatic senescence in the European badger (Meles meles): Evidence from lifetime sex-steroid profiles. Zoology 2020, 141, 125803. [Google Scholar] [CrossRef] [PubMed]
- Woodroffe, R. Alloparental behaviour in the European badger. Anim. Behav. 1993, 46, 413–415. [Google Scholar] [CrossRef]
- Rasmussen, S.L.; Hallig, J.; van Wijk, R.E.; Petersen, H.H. An investigation of endoparasites and the determinants of parasite infection in European hedgehogs (Erinaceus europaeus) from Denmark. Int. J. Parasitol. Parasites Wildl. 2021, 16, 217–227. [Google Scholar] [CrossRef]
- Marshall, H.H.; Vitikainen, E.I.; Mwanguhya, F.; Businge, R.; Kyabulima, S.; Hares, M.C.; Inzani, E.; Kalema-Zikusoka, G.; Mwesige, K.; Nichols, H.J. Lifetime fitness consequences of early-life ecological hardship in a wild mammal population. Ecol. Evol. 2017, 7, 1712–1724. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Country | Technique | Period | Sample Size | Mean Age | Age Span | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Female | Male | Total | Female | Male | Total | ||||||
| Brockie [47] | New Zealand | Tooth abrasions | 143 | 0.4 (see table text) | 0–2 | ||||||
| Brockie [64] | New Zealand | Mandible (periosteal growth lines) | 83 | <2.7* or 1.8 (see table text) | 0–7 | ||||||
| Dickman [48] | United Kingdom | Mandible (periosteal growth lines) | 1982–84 | 87 | 3.3* | 1–8 | |||||
| Döpke [49] | Germany | Mandible (periosteal growth lines) | 1980–2001 | 32 | 34 | 66 | 0.8 | 1.7 | 1.2 | 0–6 | |
| Haigh, Kelly, Butler and O’Riordan [50] | Ireland | Mandible (periosteal growth lines) | 2008–2011 | 31 | 46 | 83 (6 of unknown sex) | 2.1 | 1.9 | 1.9 | 0–9 | |
| Heddergott [51] | Thüringen, Germany | Tooth growth lines | 1993–2002 | 81 | 99 | 185 (5 of unknown sex) | 2.5 | 2.1 | 2.3* | 1–7 | |
| Heddergott [52] | Harz, Germany | Tooth growth lines | 1997–2002 | 49 | 57 | 106 | 1.6 | 1.5 | 1.5* | 1–4 | |
| Heddergott [53] | Germany, Saalfeld-Rudolstadt (Thüringen) | Tooth growth lines | 1992–2003 | 44 | 37 | 81 | 1.9 | 2.5 | 2.2* | 1–8 | |
| Heddergott and Müller [54] | Germany, Osthessen | Tooth growth lines | 1980–2006 | 62 | 61 | 124 | 1.9 | 1.7 | 1.8* | 1–6 | |
| Heddergott, Steinbach and Heddergott [55] | Germany, Heilbad Heiligenstadt (Thüringen) | Tooth growth lines | 1998–2004 | 62 | 51 | 113 | 2.2 | 2.5 | 2.3* | 1–7 | |
| Heyne [56] | Germany | Tooth abrasions+ Tooth growth lines | 1981–89 | 62 | 2* | 1–5 | |||||
| Kristoffersson [58] | Finland | Mandible (periosteal growth lines) | 1960–61 | 67 | 2.7* (individuals < 500 g excluded) | 1–7 | |||||
| Scott-Hayward [68] | Uists, Outer Hebrides, UK | Mandible (periosteal growth lines) | 66 | 2.6* | 1–6 | ||||||
| Morris [36], Morris [41], Morris [60] | Southern England, United Kingdom | Mandible (periosteal growth lines) | 1965–68 | 102 | 127 | 244 (15 of unknown sex) | 1.7 | 0–6 | |||
| Neuschulz and Schubert [60] | Dresden-Bühlau, Germany | Capture-mark-recapture | 1984–89 | 125 (60 adults and 65 juveniles marked, 53 recaptured over 5 years) | 2.1* | 1–6 | |||||
| Parkes [65] | Manawatu, New Zealand | Capture-mark-recapture + calculation using a formula [66] | 1970–1971 | 144 (73 juveniles) | 2.0 | ||||||
| Rasmussen et al. 2023 (present study) | Denmark | Mandible (periosteal growth lines) | 2016 | 109 | 177 | 388 (101 of unknown sex) | 1.6 | 2.2 (1.6 for unknown sex) | 1.8 | 0–16 | |
| Rautio, Kunnasranta, Valtonen, Ikonen, Hyvarinen, Holopainen and Kukkonen [61] | Finland | Mandible (periosteal growth lines) | 2004–2005 | 31 | 34 | 65 | 1.5 | 1.7 | 1.6 | 0–7 | |
| Reeve, Love and Shore [42] | United Kingdom | Mandible (periosteal growth lines) | 1988–89 | 68 | 1.6 | 0–7 | |||||
| Skoudlin [62] | Czech republic | Tooth abrasions | 43 | 63 | 106 | 2.5* | 1–5 | ||||
| Skoudlin [63] | Czech republic, Bohemina and Moravia | Tooth abrasions | 215 | 2.1 | 0–6 | ||||||
| Age in Years | Number of Individuals | Sex | Distribution by Sex |
|---|---|---|---|
| 0 | 109 | ||
| Male | 43 | ||
| Female | 39 | ||
| Unknown | 27 | ||
| 1 | 115 | ||
| Male | 54 | ||
| Female | 21 | ||
| Unknown | 40 | ||
| 2 | 54 | ||
| Male | 23 | ||
| Female | 15 | ||
| Unknown | 16 | ||
| 3 | 53 | ||
| Male | 29 | ||
| Female | 18 | ||
| Unknown | 6 | ||
| 4 | 22 | ||
| Male | 6 | ||
| Female | 9 | ||
| Unknown | 7 | ||
| 5 | 15 | ||
| Male | 7 | ||
| Female | 6 | ||
| Unknown | 2 | ||
| 6 | 11 | ||
| Male | 8 | ||
| Female | 1 | ||
| Unknown | 2 | ||
| 9 | 2 | ||
| Male | 2 | ||
| Female | 0 | ||
| Unknown | 0 | ||
| 10 | 4 | ||
| Male | 2 | ||
| Female | 0 | ||
| Unknown | 2 | ||
| 11 | 1 | ||
| Male | 1 | ||
| Female | 0 | ||
| Unknown | 0 | ||
| 13 | 1 | ||
| Male | 1 | ||
| Female | 0 | ||
| Unknown | 0 | ||
| 16 | 1 | ||
| Male | 1 | ||
| Female | 0 | ||
| Unknown | 0 | ||
| Total individuals | 388 |
| x | lx | ndx | nqx | npx | nLx | Tx | ex | nmx | nax |
|---|---|---|---|---|---|---|---|---|---|
| 0 | 177 | 43 | 0.243 | 0.757 | 155.5 | 460.5 | 2.602 | 0.277 | 0.5 |
| 1 | 134 | 54 | 0.403 | 0.597 | 107.0 | 305.0 | 2.276 | 0.505 | 0.5 |
| 2 | 80 | 23 | 0.287 | 0.713 | 68.5 | 198.0 | 2.475 | 0.336 | 0.5 |
| 3 | 57 | 29 | 0.509 | 0.491 | 42.5 | 129.5 | 2.272 | 0.682 | 0.5 |
| 4 | 28 | 6 | 0.214 | 0.786 | 25.0 | 87.0 | 3.107 | 0.240 | 0.5 |
| 5 | 22 | 7 | 0.318 | 0.682 | 18.5 | 62.0 | 2.818 | 0.378 | 0.5 |
| 6 | 15 | 8 | 0.533 | 0.467 | 11.0 | 43.5 | 2.900 | 0.727 | 0.5 |
| 7 | 7 | 0 | 0.000 | 1.000 | 7.0 | 32.5 | 4.643 | 0.000 | 0.5 |
| 8 | 7 | 0 | 0.000 | 1.000 | 7.0 | 25.5 | 3.643 | 0.000 | 0.5 |
| 9 | 7 | 2 | 0.286 | 0.714 | 6.0 | 18.5 | 2.643 | 0.333 | 0.5 |
| 10 | 5 | 2 | 0.400 | 0.600 | 4.0 | 12.5 | 2.500 | 0.500 | 0.5 |
| 11 | 3 | 1 | 0.333 | 0.667 | 2.5 | 8.5 | 2.833 | 0.400 | 0.5 |
| 12 | 2 | 0 | 0.000 | 1.000 | 2.0 | 6.0 | 3.000 | 0.000 | 0.5 |
| 13 | 2 | 1 | 0.500 | 0.500 | 1.5 | 4.0 | 2.000 | 0.667 | 0.5 |
| 14 | 1 | 0 | 0.000 | 1.000 | 1.0 | 2.5 | 2.500 | 0.000 | 0.5 |
| 15 | 1 | 0 | 0.000 | 1.000 | 1.0 | 1.5 | 1.500 | 0.000 | 0.5 |
| 16 | 1 | 1 | 1.000 | 0.000 | 0.5 | 0.5 | 0.500 | 2.000 | 0.5 |
| 17 | 0 | 0 | - | - | - | 0.0 | - | - | 0.5 |
| x | lx | ndx | nqx | npx | nLx | Tx | ex | nmx | nax |
|---|---|---|---|---|---|---|---|---|---|
| 0 | 109 | 39 | 0.358 | 0.642 | 89.5 | 231.5 | 2.124 | 0.436 | 0.5 |
| 1 | 70 | 21 | 0.300 | 0.700 | 59.5 | 142.0 | 2.029 | 0.353 | 0.5 |
| 2 | 49 | 15 | 0.306 | 0.694 | 41.5 | 82.5 | 1.684 | 0.361 | 0.5 |
| 3 | 34 | 18 | 0.529 | 0.471 | 25.0 | 41.0 | 1.206 | 0.720 | 0.5 |
| 4 | 16 | 9 | 0.562 | 0.438 | 11.5 | 16.0 | 1.000 | 0.783 | 0.5 |
| 5 | 7 | 6 | 0.857 | 0.143 | 4.0 | 4.5 | 0.643 | 1.500 | 0.5 |
| 6 | 1 | 1 | 1.000 | 0.000 | 0.5 | 0.5 | 0.500 | 2.000 | 0.5 |
| 7 | 0 | 0 | - | - | - | 0.0 | - | - | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasmussen, S.L.; Berg, T.B.; Martens, H.J.; Jones, O.R. Anyone Can Get Old—All You Have to Do Is Live Long Enough: Understanding Mortality and Life Expectancy in European Hedgehogs (Erinaceus europaeus). Animals 2023, 13, 626. https://doi.org/10.3390/ani13040626
Rasmussen SL, Berg TB, Martens HJ, Jones OR. Anyone Can Get Old—All You Have to Do Is Live Long Enough: Understanding Mortality and Life Expectancy in European Hedgehogs (Erinaceus europaeus). Animals. 2023; 13(4):626. https://doi.org/10.3390/ani13040626
Chicago/Turabian StyleRasmussen, Sophie Lund, Thomas B. Berg, Helle Jakobe Martens, and Owen R. Jones. 2023. "Anyone Can Get Old—All You Have to Do Is Live Long Enough: Understanding Mortality and Life Expectancy in European Hedgehogs (Erinaceus europaeus)" Animals 13, no. 4: 626. https://doi.org/10.3390/ani13040626