
Evaluation of Muscle Proteins for Estimating the Post-Mortem Interval in Veterinary Forensic Pathology

 ,
,  , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Muscle Sample Collection
2.2. Sampling Procedure
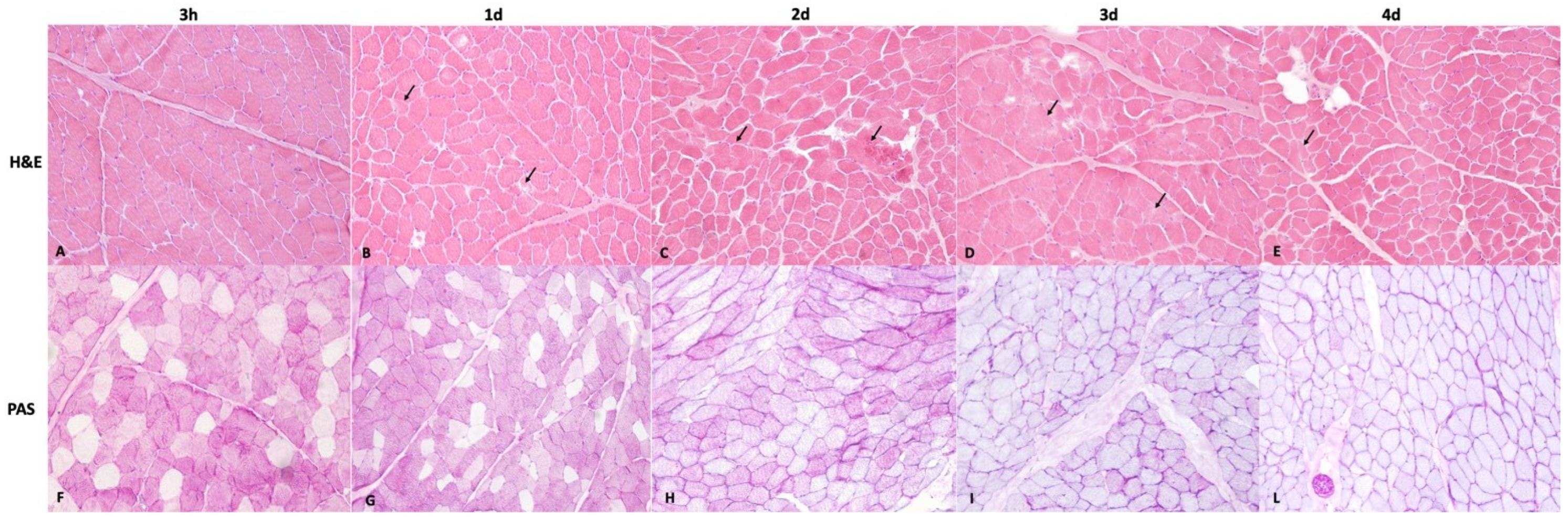
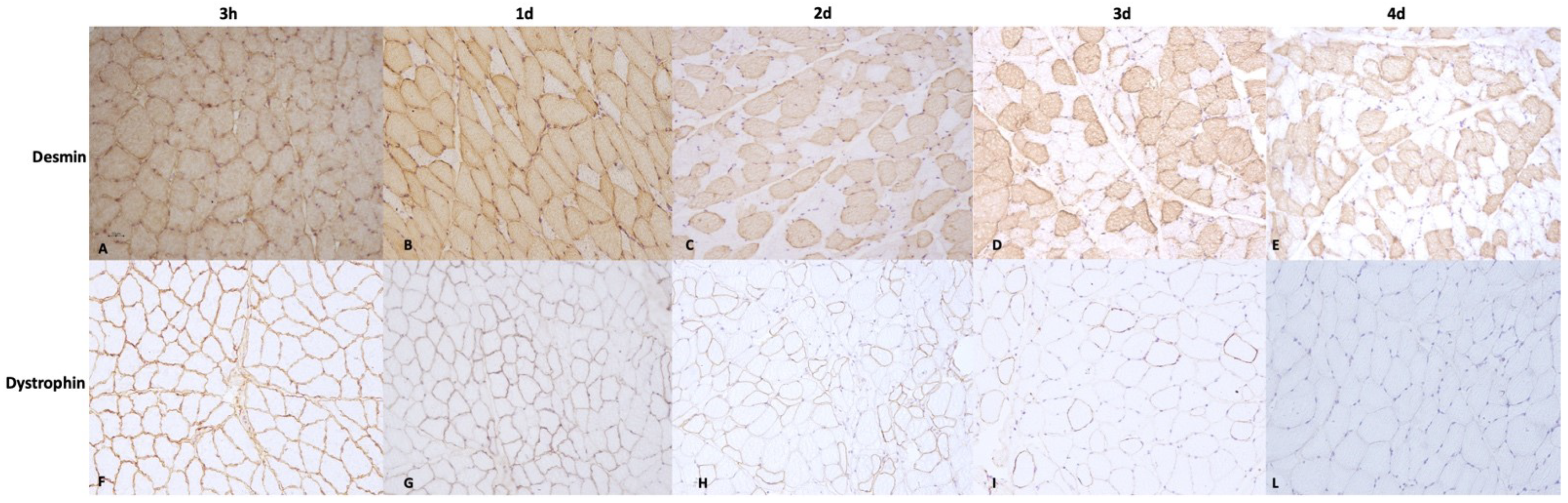
2.3. Histology and Immunohistochemical Examination
- Mouse anti-DYS-1 monoclonal antibody (NCL-DYS1) (NovoCastra Laboratories Ltd, Newcastle, UK) directed against the ROD domain of the muscular dystrophin protein;
- Mouse anti-DYS-2 monoclonal antibody (NCL-DYS2) (NovoCastra Laboratories Ltd, Newcastle, UK) directed against the C-terminal domain of the muscular dystrophin protein;
- Mouse anti-desmin clone D33 monoclonal antibody (M0760) (DakoCytomation, Denmark).
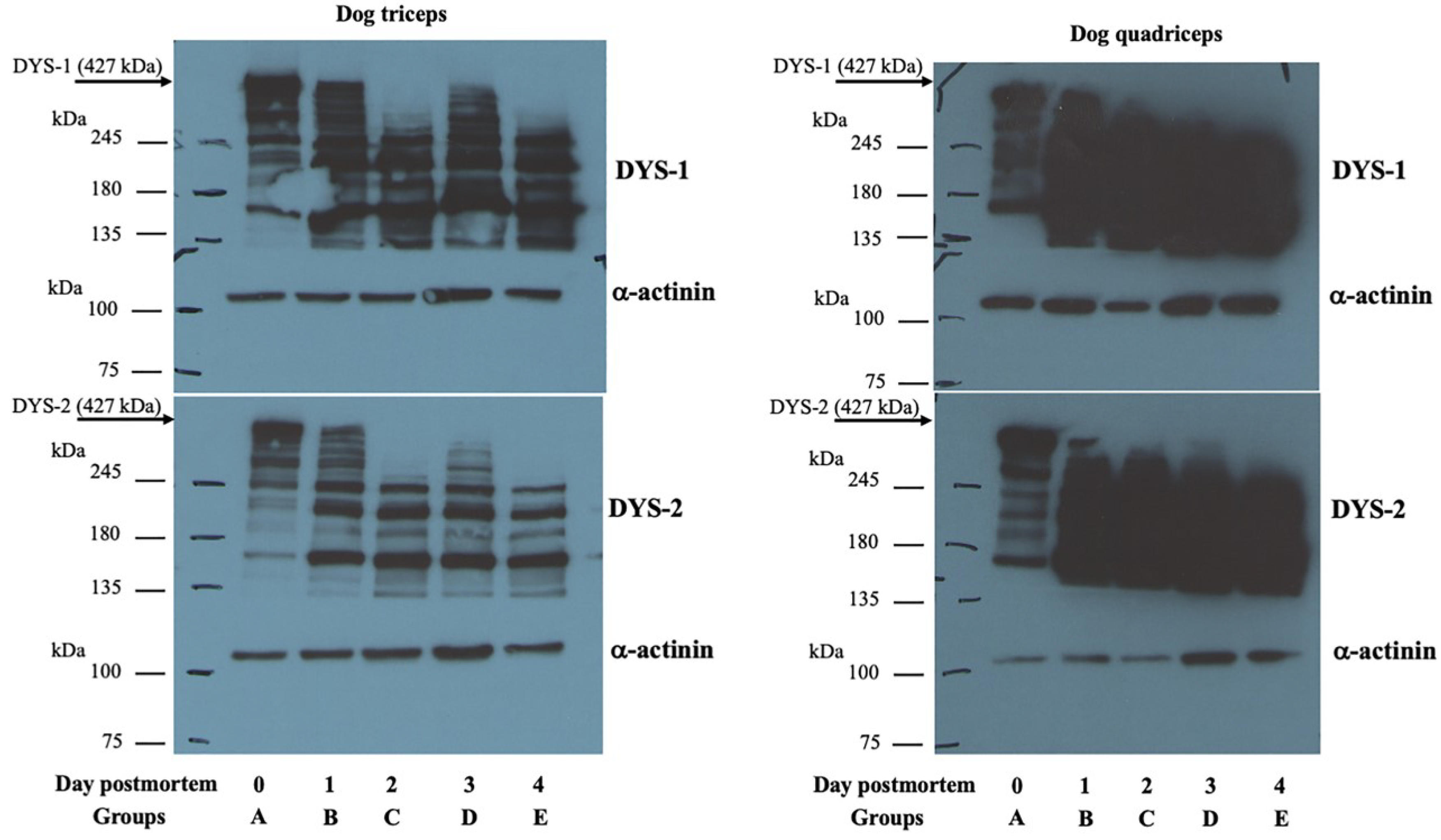
2.4. Western Blotting
- Mouse anti-GAPDH monoclonal antibody (clone 6C5, sc-32233, Santa Cruz Biotechnology Inc, Dallas, TX, USA);
- Mouse anti-alpha-actinin (clone H-2, sc-17829, Santa Cruz Biotechnology Inc, Dallas, TX, USA).
2.5. Statistical Analysis
3. Results
3.1. Histopathological Examination
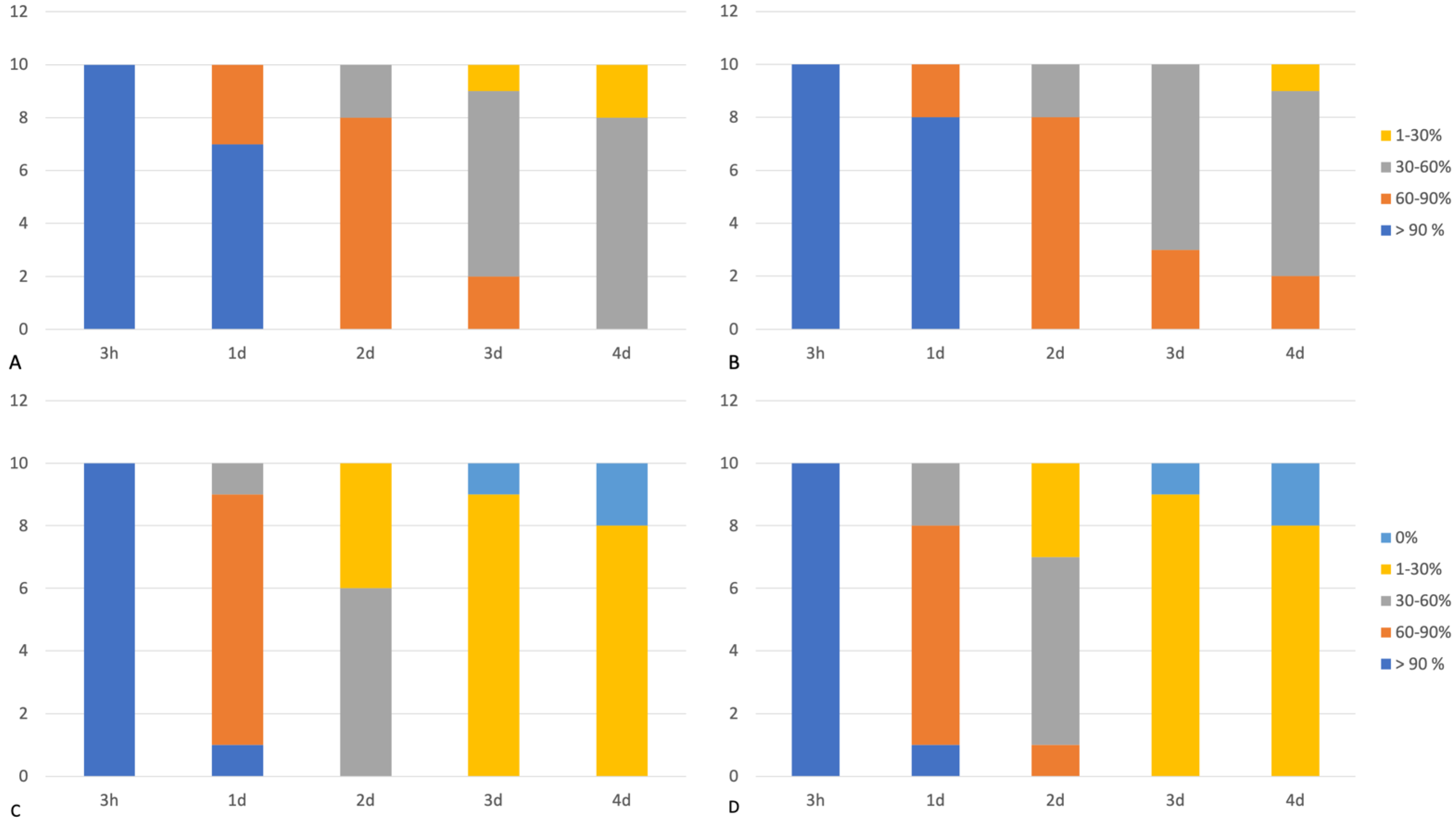
3.2. Immunohistochemical Examination
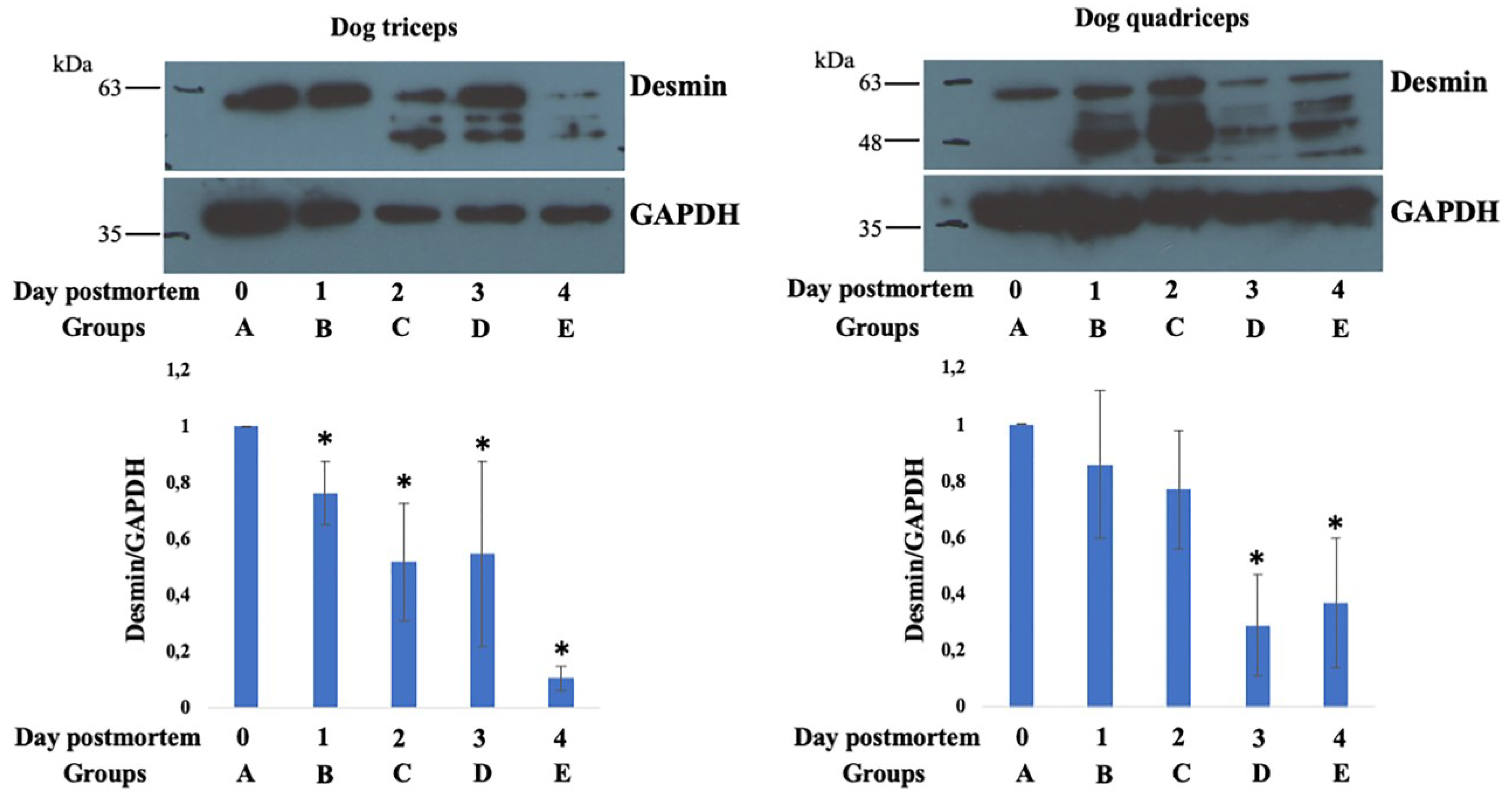
3.3. Western Blot Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biswas, G. Review of Forensic Medicine and Toxicology; Pvt Brooks; Jaypee Brothers Medical Publishers: New Delhi, India, 2012; p. 22. [Google Scholar]
- Merk, M.D. Veterinary forensics: Animal Cruelty Investigations; Wiley: New Jersey, NJ, USA, 2008; p. 241. [Google Scholar]
- Shedge, R.; Krishan, K.; Warrier, V.; Kanchan, T. Postmortem Changes; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Brooks, J.W. Postmortem Changes in Animal Carcasses and Estimation of the Postmortem Interval. Veter-Pathol. 2016, 53, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Guebelin, D.L.C.; Dobay, A.; Ebert, L.; Betschart, E.; Thali, M.J.; Franckenberg, S. Correlation of age, sex and season with the state of human decomposition as quantified by postmortem computed tomography. Forensic Sci. Med. Pathol. 2021, 17, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Matuszewski, S.; Konwerski, S.; Frątczak, K.; Szafałowicz, M. Effect of body mass and clothing on decomposition of pig carcasses. Int. J. Leg. Med. 2014, 128, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.; Adlam, R.E.; Moffatt, C. Debugging decomposition data—Comparative taphonomic studies and the influence of insects and carcass size on decomposition rate. J. Forensic Sci. 2010, 50, 8–13. [Google Scholar] [CrossRef]
- Sutherland, A.; Myburgh, J.; Steyn, M.; Becker, P.J. The effect of body size on the rate of decomposition in a temperate region of South Africa. Forensic Sci. Int. 2013, 231, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Byard, R.W. Factors and processes causing accelerated decomposition in human cadavers—An overview. J. Forensic Leg. Med. 2011, 18, 6–9. [Google Scholar] [CrossRef]
- McIntosh, C.S.; Dadour, I.R.; Voss, S.C. A comparison of carcass decomposition and associated insect succession onto burnt and unburnt pig carcasses. Int. J. Leg. Med. 2016, 131, 835–845. [Google Scholar] [CrossRef]
- Archer, M. Rainfall and temperature effects on the decomposition rate of exposed neonatal remains. Sci. Justice 2004, 44, 35–41. [Google Scholar] [CrossRef]
- Pittner, S.; Bugelli, V.; Weitgasser, K.; Zissler, A.; Sanit, S.; Lutz, L.; Monticelli, F.; Campobasso, C.P.; Steinbacher, P.; Amend, J. A field study to evaluate PMI estimation methods for advanced decomposition stages. Int. J. Legal Med. 2020, 134, 1361–1373. [Google Scholar] [CrossRef]
- Dogan, K.H.; Gunaydin, G.; Demirci, S.; Koç, S. Postmortem Changes in Element Levels in Rat Skeletal Muscle Tissue. Turk. Klin. J. Med. Sci. 2010, 30, 1332–1338. [Google Scholar] [CrossRef] [Green Version]
- Sabucedo, A.J.; Furton, K.G. Estimation of postmortem interval using the protein marker cardiac Troponin I. Forensic Sci. Int. 2003, 134, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Pittner, S.; Ehrenfellner, B.; Zissler, A.; Racher, V.; Trutschnig, W.; Bathke, A.C.; Sänger, A.M.; Stoiber, W.; Steinbacher, P.; Monticelli, F.C. First application of a protein-based approach for time since death estimation. Int. J. Leg. Med. 2016, 131, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martínez, C.; Pérez-Cárceles, M.D.; Legaz, I.; Prieto-Bonete, G.; Luna, A. Quantification of nitrogenous bases, DNA and Collagen type I for the estimation of the postmortem interval in bone remains. Forensic Sci. Int. 2017, 281, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Ortmann, J.; Doberentz, E.; Madea, B. Immunohistochemical methods as an aid in estimating the time since death. Forensic Sci. Int. 2017, 273, 71–79. [Google Scholar] [CrossRef]
- Zissler, A.; Stoiber, W.; Steinbacher, P.; Geissenberger, J.; Monticelli, F.C.; Pittner, S. Postmortem Protein Degradation as a Tool to Estimate the PMI: A Systematic Review. Diagnostics 2020, 10, 1014. [Google Scholar] [CrossRef]
- Pittner, S.; Monticelli, F.C.; Pfisterer, A.; Zissler, A.; Sänger, A.M.; Stoiber, W.; Steinbacher, P. Postmortem degradation of skeletal muscle proteins: A novel approach to determine the time since death. Int. J. Leg. Med. 2015, 130, 421–431. [Google Scholar] [CrossRef]
- Li, C.; Ma, D.; Deng, K.; Chen, Y.; Huang, P.; Wang, Z. Application of MALDI-TOF MS for Estimating the Postmortem Interval in Rat Muscle Samples. J. Forensic Sci. 2017, 62, 1345–1350. [Google Scholar] [CrossRef]
- Pittner, S.; Ehrenfellner, B.; Monticelli, F.C.; Zissler, A.; Sänger, A.M.; Stoiber, W.; Steinbacher, P. Postmortem muscle protein degradation in humans as a tool for PMI delimitation. Int. J. Leg. Med. 2016, 130, 1547–1555. [Google Scholar] [CrossRef]
- Vass, A.A.; Barshick, S.A.; Sega, G.; Caton, J.; Skeen, J.T.; Love, J.C.; Synstelien, J.A. Decomposition Chemistry of Human Remains: A New Methodology for Determining the Postmortem Interval. J. Forensic Sci. 2002, 47, 542–553. [Google Scholar] [CrossRef]
- Poloz, Y.O.; O’Day, D.H. Determining time of death: Temperature-dependent postmortem changes in calcineurin A, MARCKS, CaMKII, and protein phosphatase 2A in mouse. Int. J. Leg. Med. 2009, 123, 305–314. [Google Scholar] [CrossRef]
- Kang, S.; Kassam, N.; Gauthier, M.L.; O’Day, D.H. Post-mortem changes in calmodulin binding proteins in muscle and lung. Forensic Sci. Int. 2003, 131, 140–147. [Google Scholar] [CrossRef]
- Li, C.; Wang, Q.; Zhang, Y.; Lin, H.; Zhang, J.; Huang, P.; Wang, Z. Research progress in the estimation of the postmortem interval by Chinese forensic scholars. Forensic Sci. Res. 2016, 1, 3–13. [Google Scholar] [CrossRef]
- Zissler, A.; Ehrenfellner, B.; Foditsch, E.E.; Monticelli, F.C.; Pittner, S. Does altered protein metabolism interfere with postmortem degradation analysis for PMI estimation? Int. J. Legal Med. 2018, 32, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Foditsch, E.E.; Saenger, A.M.; Monticelli, F.C. Skeletal muscle proteins: A new approach to delimitate the time since death. Int. J. Leg. Med. 2015, 130, 433–440. [Google Scholar] [CrossRef]
- Horn, L.; Range, F.; Huber, L. Dogs’ attention towards humans depends on their relationship, not only on social familiarity. Anim. Cogn. 2013, 16, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Williams, V.; Dale, A.; Clarke, N.; Garrett, N. Animal abuse and family violence: Survey on the recognition of animal abuse by veterinarians in New Zealand and their understanding of the correlation between animal abuse and human violence. N. Z. Veter.-J. 2008, 56, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Siegel, D.; van Uhm, D. Illegal dogfighting: Sport or crime? Trends Organ. Crime 2021, 24, 563–580. [Google Scholar] [CrossRef]
- Mennonna, G.; Murino, C.; Micieli, F.; Costagliola, A.; D’Angelo, D.; Paciello, O.; Fatone, G.; Lamagna, F.; Navas, L.; Pompameo, M.; et al. Geographical information system analysis on road accidents involving wandering dogs in the urban area of Naples. Geospat. Health 2018, 13. [Google Scholar] [CrossRef]
- De Biase, D.; Piegari, G.; Prisco, F.; Cimmino, I.; D’Aquino, I.; Baldassarre, V.; Oriente, F.; Papparella, S.; Paciello, O. Implication of the NLRP3 Inflammasome in Bovine Age-Related Sarcopenia. Int. J. Mol. Sci. 2021, 22, 3609. [Google Scholar] [CrossRef] [PubMed]
- Stankler, L.; Walker, F. Periodic acid-Schiff (PAS) staining for glycogen in clinically normal psoriatic and non-psoriatic skin. Br. J. Dermatol. 1976, 95, 599–601. [Google Scholar] [CrossRef]
- Piegari, G.; De Biase, D.; D’Aquino, I.; Prisco, F.; Fico, R.; Ilsami, R.; Pozzato, N.; Genovese, A.; Paciello, O. Diagnosis of Drowning and the Value of the Diatom Test in Veterinary Forensic Pathology. Front. Veter.-Sci. 2019, 6, 404. [Google Scholar] [CrossRef]
- Cerulo, G.; Tafuri, S.; De Pasquale, V.; Rea, S.; Romano, S.; Costagliola, A.; Della Morte, R.; Avallone, L.; Pavone, L.M. Serotonin activates cell survival and apoptotic death responses in cultured epithelial thyroid cells. Biochimie 2014, 105, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Persico, M.; Ramunno, A.; Maglio, V.; Franceschelli, S.; Esposito, C.; Carotenuto, A.; Brancaccio, D.; De Pasquale, V.; Pavone, L.M.; Varra, M.; et al. New anticancer agents mimicking protein recognition motifs. J. Med. Chem. 2013, 12, 6666–6680. [Google Scholar] [CrossRef] [PubMed]
- Pavone, L.M.; Rea, S.; Trapani, F.; De Pasquale, V.; Tafuri, S.; Papparella, S.; Paciello, O. Role of serotonergic system in the pathogenesis of fibrosis in canine idiopathic inflammatory myopathies. Neuromuscul. Disord. 2012, 22, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Spina, A.; Rea, S.; De Pasquale, V.; Mastellone, V.; Avallone, L.; Pavone, L.M. Fate Map of Serotonin Transporter-Expressing Cells in Developing Mouse Thyroid. Anat. Rec. 2011, 294, 384–390. [Google Scholar] [CrossRef]
- Takeichi, S.; Tokunaga, I.; Yoshima, K.; Maeiwa, M.; Bando, Y.; Kominami, E.; Katunuma, N. Mechanism of postmortem autolysis of skeletal muscle. Biochem. Med. 1984, 32, 341–348. [Google Scholar] [CrossRef]
- Chauhan, S.S.; England, E.M. Postmortem glycolysis and glycogenolysis: Insights from species comparisons. Meat Sci. 2018, 144, 118–126. [Google Scholar] [CrossRef]
- Erlandsson, M.; Munro, R. Estimation of the post-mortem interval in beagle dogs. Sci. Justice 2007, 47, 150–154. [Google Scholar] [CrossRef]
- Tavichakorntrakool, R.; Prasongwattana, V.; Sriboonlue, P.; Puapairoj, A.; Pongskul, J.; Khuntikeo, N.; Hanpanich, W.; Yenchitsomanus, P.-T.; Wongkham, C.; Thongboonkerd, V. Serial analyses of postmortem changes in human skeletal muscle: A case study of alterations in proteome profile, histology, electrolyte contents, water composition, and enzyme activity. Proteom.–Clin. Appl. 2008, 2, 1255–1264. [Google Scholar] [CrossRef]
- Piegari, G. New Insights in Veterinary Forensic Medicine and Pathology. Ph.D. Thesis, University of Naples “Federico II”, Naples, India, 2018. [Google Scholar]
- Purintrapiban, J.; Wang, M.C.; Forsberg, N.E. Degradation of sarcomeric and cytoskeletal proteins in cultured skeletal muscle cells. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 136, 393–401. [Google Scholar] [CrossRef]
- Hoffman, E.P.; Brown, R.H., Jr.; Kunkel, L.M. Dystrophin: The protein product of the Duchenne muscular dystrophy locus. Cell 1987, 24, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Wojtysiak, D.; Górska, M. Effect of Aging Time on Meat Quality and Rate of Desmin and Dystrophin Degradation of Pale, Soft, Exudative (PSE) and Normal Turkey Breast Muscle. Folia Biol. 2018, 66, 63–72. [Google Scholar] [CrossRef]
- Choi, K.M.; Zissler, A.; Kim, E.; Ehrenfellner, B.; Cho, E.; Lee, S.I.; Steinbacher, P.; Yun, K.N.; Shin, J.H.; Kim, J.Y.; et al. Postmortem proteomics to discover biomarkers for forensic PMI estimation. Int. J. Leg. Med. 2019, 133, 899–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piegari, G.; De Pasquale, V.; d’Aquino, I.; De Biase, D.; Caccia, G.; Campobasso, C.P.; Tafuri, S.; Russo, V.; Paciello, O. Evaluation of Muscle Proteins for Estimating the Post-Mortem Interval in Veterinary Forensic Pathology. Animals 2023, 13, 563. https://doi.org/10.3390/ani13040563
Piegari G, De Pasquale V, d’Aquino I, De Biase D, Caccia G, Campobasso CP, Tafuri S, Russo V, Paciello O. Evaluation of Muscle Proteins for Estimating the Post-Mortem Interval in Veterinary Forensic Pathology. Animals. 2023; 13(4):563. https://doi.org/10.3390/ani13040563
Chicago/Turabian StylePiegari, Giuseppe, Valeria De Pasquale, Ilaria d’Aquino, Davide De Biase, Giulia Caccia, Carlo Pietro Campobasso, Simona Tafuri, Valeria Russo, and Orlando Paciello. 2023. "Evaluation of Muscle Proteins for Estimating the Post-Mortem Interval in Veterinary Forensic Pathology" Animals 13, no. 4: 563. https://doi.org/10.3390/ani13040563