
Distinguishing the Effects of Water Volumes versus Stocking Densities on the Skeletal Quality during the Pre-Ongrowing Phase of Gilthead Seabream (Sparus aurata)
 , , , , ,
, , , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing Setup
2.2. Sampling and Analyses
2.3. Skeletal Anomalies Survey
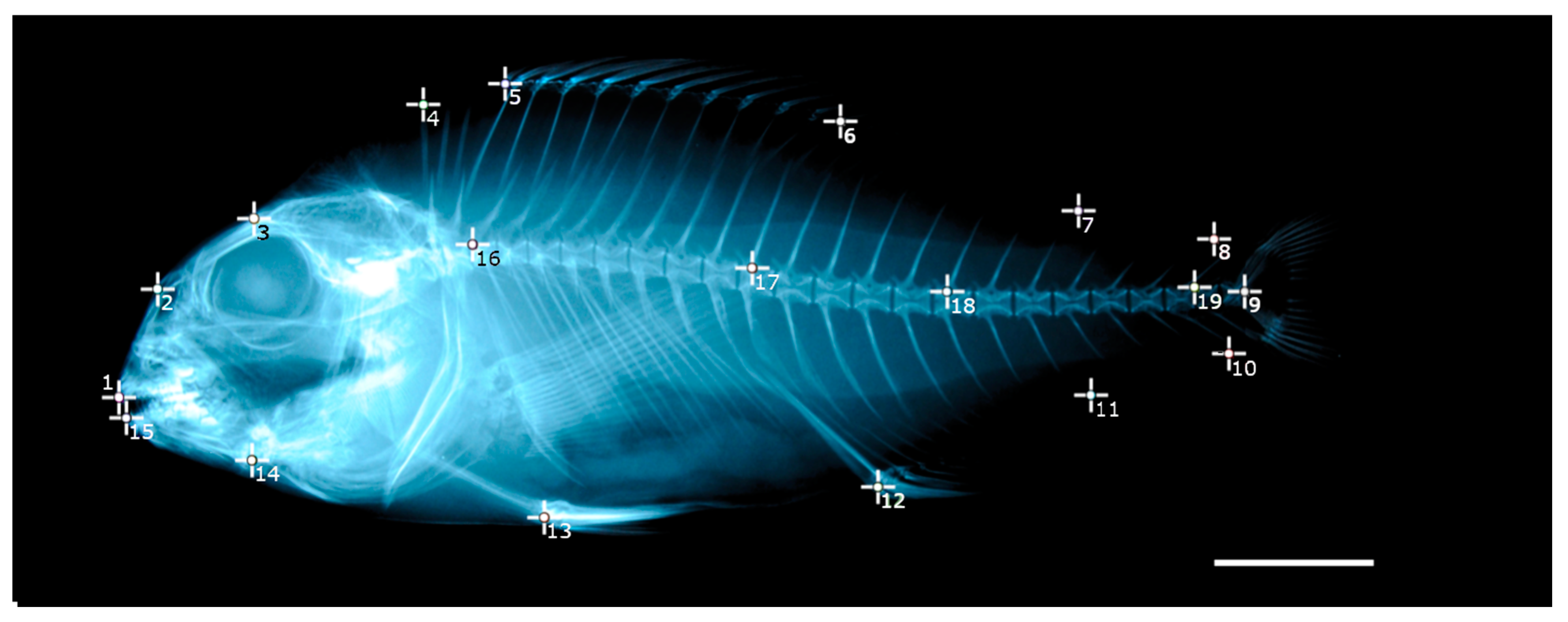
2.4. Geometric Morphometrics
3. Results
3.1. Performance Indicators
3.2. Anatomical Analyses
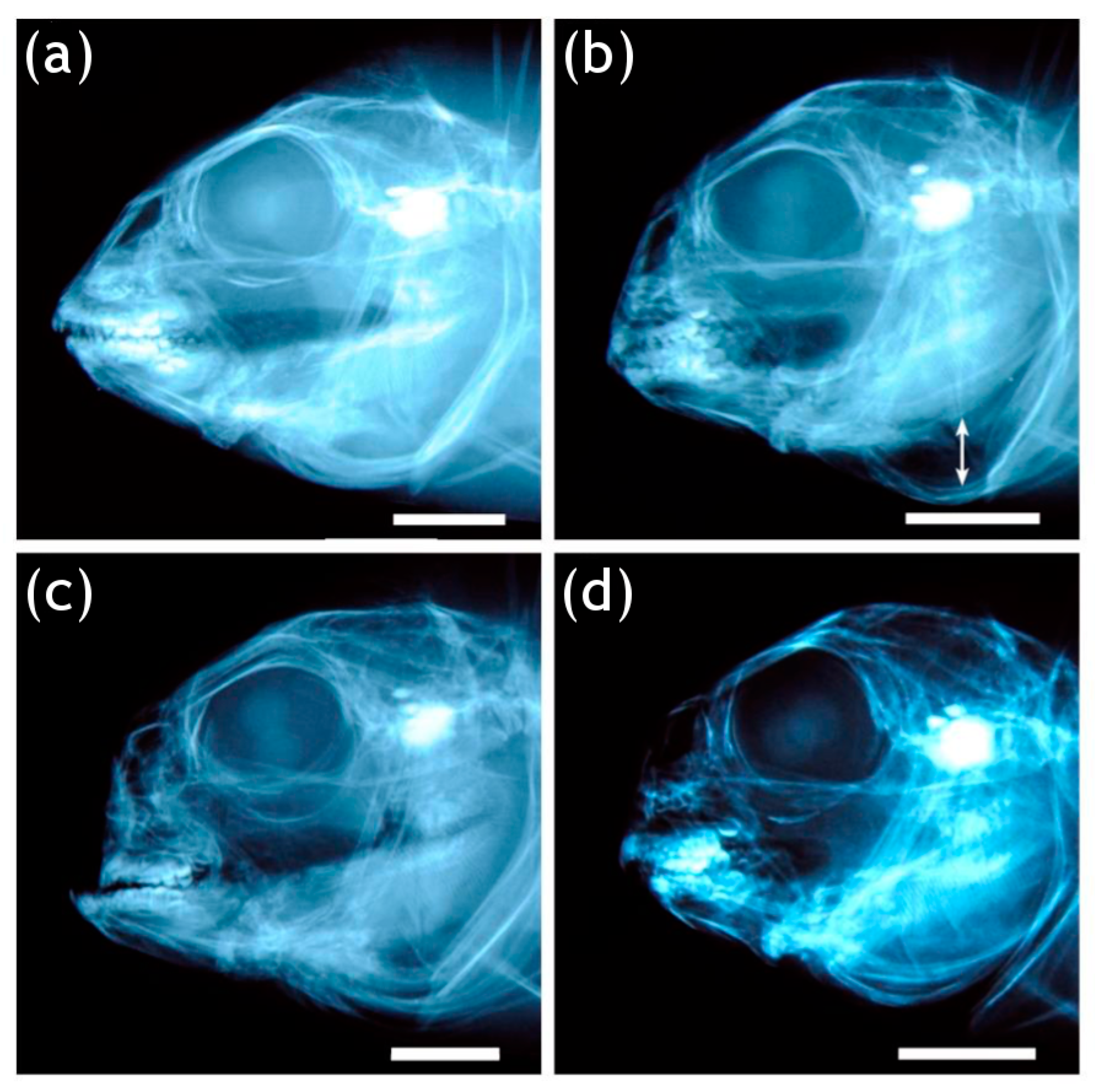
3.3. Cephalic Skeleton
3.4. Axial Skeleton
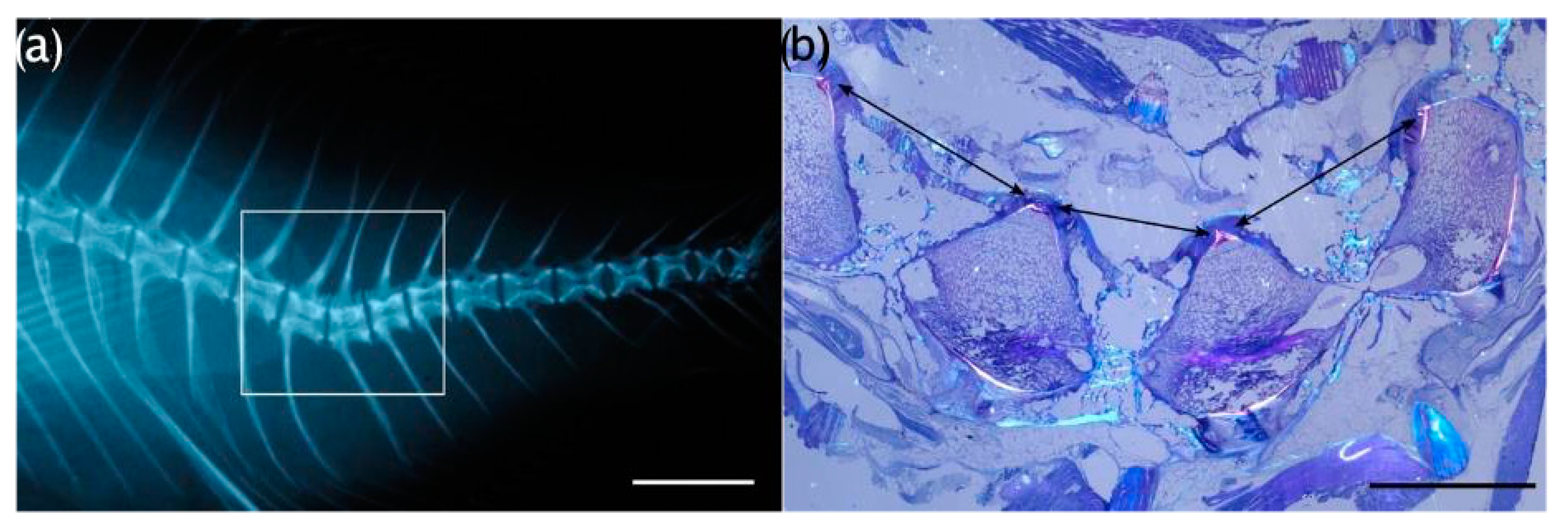
3.5. Histological Evaluations
3.6. Principal Components Analysis
3.7. Geometric Morphometrics
3.8. Calcium-Phosphorous Mineral Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| bgPCA | Between group principal components analysis |
| BM | Binary Matrix |
| FCR | Food conversion ratio |
| HD | High Density |
| K | Fulton’s condition factor |
| KS | Kolmogrov–Smirnov test for equal distribution |
| LDA | Linear discriminant analysis |
| LD | Low Density |
| MD | Medium Density |
| PCA | Principal components analysis |
| RM | Raw Matrix |
| SGR | Specific Growth Rate |
| T0 | Initial batch of gilthead seabream before being placed in experimental conditions |
| TF | Final samples of gilthead seabream after being reared in experimental conditions |
| TL | Total Length |
| TPS | Thin plate splines |
| WW | Wet Weight |
References
- FAO Fisheries & Aquaculture—FishStatJ. Available online: http://www.fao.org/fishery/statistics/software/fishstatj/en (accessed on 22 October 2022).
- Pavlidis, M.A.; Mylonas, C.C. Sparidae; John Wiley & Sons: Chichester, UK, 2011. [Google Scholar]
- Llorente, I.; Fernández-Polanco, J.; Baraibar-Diez, E.; Odriozola, M.D.; Bjørndal, T.; Asche, F.; Guillen, J.; Avdelas, L.; Nielsen, R.; Cozzolino, M.; et al. Assessment of the economic performance of the seabream and seabass aquaculture industry in the European Union. Mar. Policy 2020, 117, 103876. [Google Scholar] [CrossRef]
- Hough, C. Manual of Control of Malformations in Fish Aquaculture. Science and Practice; Federation of European Aquaculture Producers, RapidPRess: Luxembourg, 2009; p. 150. [Google Scholar]
- Bardon, A.; Vandeputte, M.; Dupont-Nivet, M.; Chavanne, H.; Haffray, P.; Vergnet, A.; Chatain, B. What is the heritable component of spinal deformities in the European sea bass (Dicentrarchus labrax)? Aquaculture 2009, 294, 194–201. [Google Scholar] [CrossRef]
- Daoulas, C.; Economou, A.; Bantavas, I. Osteological abnormalities in laboratory reared sea-bass (Dicentrarchus labrax) fingerlings. Aquaculture 1991, 97, 169–180. [Google Scholar] [CrossRef]
- Le Vay, L.; Carvalho, G.; Quinitio, E.; Lebata, J.; Ut, V.; Fushimi, H. Quality of hatchery-reared juveniles for marine fisheries stock enhancement. Aquaculture 2007, 268, 169–180. [Google Scholar] [CrossRef]
- Lijalad, M.; Powell, M.D. Effects of lower jaw deformity on swimming performance and recovery from exhaustive exercise in triploid and diploid Atlantic salmon Salmo salar L. Aquaculture 2009, 290, 145–154. [Google Scholar] [CrossRef]
- Boglione, C.; Gavaia, P.; Koumoundouros, G.; Gisbert, E.; Moren, M.; Fontagné, S.; Witten, P.E. Skeletal anomalies in reared European fish larvae and juveniles. Part 1: Normal and anomalous skeletogenic processes. Rev. Aquac. 2013, 5, S99–S120. [Google Scholar] [CrossRef]
- Nielsen, R.; Ankamah-Yeboah, I.; Llorente, I. Technical efficiency and environmental impact of seabream and seabass farms. Aquac. Econ. Manag. 2020, 25, 106–125. [Google Scholar] [CrossRef]
- Boglione, C.; Costa, C. Skeletal Deformities and Juvenile Quality. Sparidae 2011, 233–294. [Google Scholar] [CrossRef]
- Koumoundouros, G.; Gagliardi, F.; Divanach, P.; Boglione, C.; Cataudella, S.; Kentouri, M. Normal and abnormal osteological development of caudal fin in Sparus aurata L. fry. Aquaculture 1997, 149, 215–226. [Google Scholar] [CrossRef]
- Costa, C.; Antonucci, F.; Boglione, C.; Menesatti, P.; Vandeputte, M.; Chatain, B. Automated sorting for size, sex and skeletal anomalies of cultured seabass using external shape analysis. Aquac. Eng. 2013, 52, 58–64. [Google Scholar] [CrossRef]
- Puszkarski, J.; Śniadach, O. Instruments to implement sustainable aquaculture in the European Union. Mar. Policy 2022, 144, 105215. [Google Scholar] [CrossRef]
- Boglione, C.; Gisbert, E.; Gavaia, P.; Witten, P.E.; Moren, M.; Fontagné, S.; Koumoundouros, G. Skeletal anomalies in reared European fish larvae and juveniles. Part 2: Main typologies, occurrences and causative factors. Rev. Aquac. 2013, 5, S121–S167. [Google Scholar] [CrossRef]
- Eissa, A.E.; Abu-Seida, A.M.; Ismail, M.M.; Abu-Elala, N.M.; Abdelsalam, M. A comprehensive overview of the most common skeletal deformities in fish. Aquac. Res. 2021, 52, 2391–2402. [Google Scholar] [CrossRef]
- Kourkouta, C.; Tsipourlianos, A.; Power, D.M.; Moutou, K.A.; Koumoundouros, G. Variability of key-performance-indicators in commercial gilthead seabream hatcheries. Sci. Rep. 2022, 12, 17896. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Ghrab, W.; Roo, J.; Hamre, K.; Hernández-Cruz, C.M.; Bernardini, G.; Terova, G.; Saleh, R. Organic, inorganic and nanoparticles of Se, Zn and Mn in early weaning diets for gilthead seabream (Sparus aurata; Linnaeus, 1758). Aquac. Res. 2016, 48, 2852–2867. [Google Scholar] [CrossRef]
- Izquierdo, M.; Domínguez, D.; Jiménez, J.I.; Saleh, R.; Hernández-Cruz, C.M.; Zamorano, M.J.; Hamre, K. Interaction between taurine, vitamin E and vitamin C in microdiets for gilthead seabream (Sparus aurata) larvae. Aquaculture 2018, 498, 246–253. [Google Scholar] [CrossRef]
- Dominguez, D.; Montero, D.; Zamorano, M.J.; Castro, P.; Fontanillas, R.; Prabhu, P.A.J.; Izquierdo, M. Effects of vitamin D3 supplementation in gilthead seabream (Sparus aurata) juveniles fed diets high in plant based feedstuffs. Aquaculture 2021, 543, 736991. [Google Scholar] [CrossRef]
- Tseng, Y.; Dominguez, D.; Bravo, J.; Acosta, F.; Robaina, L.; Geraert, P.-A.; Kaushik, S.; Izquierdo, M. Organic Selenium (OH-MetSe) Effect on Whole Body Fatty Acids and Mx Gene Expression against Viral Infection in Gilthead Seabream (Sparus aurata) Juveniles. Animals 2021, 11, 2877. [Google Scholar] [CrossRef]
- Sivagurunathan, U.; Dominguez, D.; Tseng, Y.; Eryalçın, K.M.; Roo, J.; Boglione, C.; Prabhu, P.A.J.; Izquierdo, M. Effects of dietary vitamin D3 levels on survival, mineralization, and skeletal development of gilthead seabream (Sparus aurata) larvae. Aquaculture 2022, 560, 738505. [Google Scholar] [CrossRef]
- Dominguez, D.; Castro, P.; Lall, S.; Montero, D.; Zamorano, M.J.; Fontanillas, R.; Izquierdo, M. Effects of Menadione Sodium Bisulphite (Vitamin K3) Supplementation of the Diets Based on Plant Feed Ingredients on Growth and Bone Health of Gilthead Seabream (Sparus aurata) Fingerlings. Aquac. Nutr. 2022, 2022, 1613030. [Google Scholar] [CrossRef]
- Ferosekhan, S.; Sarih, S.; Afonso, J.M.; Zamorano, M.J.; Fontanillas, R.; Izquierdo, M.; Kaushik, S.; Montero, D. Selection for high growth improves reproductive performance of gilthead seabream Sparus aurata under mass spawning conditions, regardless of the dietary lipid source. Anim. Reprod. Sci. 2022, 241, 106989. [Google Scholar] [CrossRef] [PubMed]
- Berillis, P. Skeletal Deformities in Seabreams. Understanding the Genetic Origin Can Improve Production? J. Fish. Sci. 2017, 11, 57. [Google Scholar] [CrossRef]
- Fragkoulis, S.; Batargias, C.; Kolios, P.; Koumoundouros, G. Genetic parameters of the upper-jaw abnormalities in Gilthead seabream Sparus aurata. Aquaculture 2018, 497, 226–233. [Google Scholar] [CrossRef]
- Fragkoulis, S.; Economou, I.; Moukas, G.; Koumoundouros, G.; Batargias, C. Caudal fin abnormalities in Gilthead seabream (Sparus aurata L.) have a strong genetic variance component. J. Fish Dis. 2020, 43, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Balbuena-Pecino, S.; Riera-Heredia, N.; Vélez, E.J.; Gutiérrez, J.; Navarro, I.; Riera-Codina, M.; Capilla, E. Temperature Affects Musculoskeletal Development and Muscle Lipid Metabolism of Gilthead Sea Bream (Sparus aurata). Front. Endocrinol. 2019, 10, 173. [Google Scholar] [CrossRef]
- Suniaga, S.; Rolvien, T.; Scheidt, A.V.; Fiedler, I.A.K.; Bale, H.A.; Huysseune, A.; Witten, P.E.; Amling, M.; Busse, B. Increased mechanical loading through controlled swimming exercise induces bone formation and mineralization in adult zebrafish. Sci. Rep. 2018, 8, 3646. [Google Scholar] [CrossRef]
- Hall, B.K.; Witten, P.E. Plasticity and variation of skeletal cells and tissues and the evolutionary development of actinopterygian fishes. In Evolution and Development of Fishes; Johanson, Z., Underwood, C., Richter, M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2018; pp. 126–143. [Google Scholar]
- Boglione, C. Skeletal abnormalities. In Climate Change and Non-Infectious Fish Disorders; Woo, P.T.K., Iwama, G.K., Eds.; Cabi: Wallingford, UK; Boston, MA, USA, 2020; pp. 54–79. [Google Scholar]
- Palstra, A.P.; Roque, A.; Kruijt, L.; Jéhannet, P.; Pérez-Sánchez, J.; Dirks, R.P. Physiological Effects of Water Flow Induced Swimming Exercise in Seabream Sparus aurata. Front. Physiol. 2020, 11, 610049. [Google Scholar] [CrossRef]
- Araújo-Luna, R.; Ribeiro, L.; Bergheim, A.; Pousão-Ferreira, P. The impact of different rearing condition on gilthead seabream welfare: Dissolved oxygen levels and stocking densities. Aquac. Res. 2018, 49, 3845–3855. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Nazzaro-Alvarez, J.; Jardí-Pons, A.; Reig, L.; Carella, F.; Carrassón, M.; Roque, A. Linking stocking densities and feeding strategies with social and individual stress responses on gilthead seabream (Sparus aurata). Physiol. Behav. 2020, 213, 112723. [Google Scholar] [CrossRef]
- Canario, A.V.M.; Condeça, J.; Power, D.M.; Ingleton, P.M. The effect of stocking density on growth in the gilthead sea-bream, Sparus aurata (L.). Aquac. Res. 1998, 29, 177–181. [Google Scholar] [CrossRef]
- Montero, D.; Blazer, V.; Socorro, J.; Izquierdo, M.; Tort, L. Dietary and culture influences on macrophage aggregate parameters in gilthead seabream (Sparus aurata) juveniles. Aquaculture 1999, 179, 523–534. [Google Scholar] [CrossRef]
- Montero, D.; Robaina, L.; Socorro, J.; Vergara, J.; Tort, L.; Izquierdo, M. Alteration of liver and muscle fatty acid composition in gilthead seabream (Sparus aurata) juveniles held at high stocking density and fed an essential fatty acid deficient diet. Fish Physiol. Biochem. 2001, 24, 63–72. [Google Scholar] [CrossRef]
- Tort, L.; Sunyer, J.; Gómez, E.; Molinero, A. Crowding stress induces changes in serum haemolytic and agglutinating activity in the gilthead sea bream Sparus aurata. Vet. Immunol. Immunopathol. 1996, 51, 179–188. [Google Scholar] [CrossRef]
- Prestinicola, L.; Boglione, C.; Makridis, P.; Spanò, A.; Rimatori, V.; Palamara, E.; Scardi, M.; Cataudella, S. Environmental Conditioning of Skeletal Anomalies Typology and Frequency in Gilthead Seabream (Sparus aurata L., 1758) Juveniles. PLoS ONE 2013, 8, e55736. [Google Scholar] [CrossRef]
- Boglione, C.; Gagliardi, F.; Scardi, M.; Cataudella, S. Skeletal descriptors and quality assessment in larvae and post-larvae of wild-caught and hatchery-reared gilthead sea bream (Sparus aurata L. 1758). Aquaculture 2001, 192, 1–22. [Google Scholar] [CrossRef]
- Divanach, P.; Kentouri, M. Hatchery techniques for specific diversification in Mediterranean finfish larviculture. Cah. Opt. Medit. 2000, 47, 75–87. [Google Scholar]
- Martini, A.; Huysseune, A.; Witten, P.E.; Boglione, C. Plasticity of the skeleton and skeletal deformities in zebrafish (Danio rerio) linked to rearing density. J. Fish Biol. 2020, 98, 971–986. [Google Scholar] [CrossRef]
- Di Biagio, C.; Dellacqua, Z.; Martini, A.; Huysseune, A.; Scardi, M.; Witten, P.E.; Boglione, C. A Baseline for Skeletal Investigations in Medaka (Oryzias latipes): The Effects of Rearing Density on the Postcranial Phenotype. Front. Endocrinol. 2022, 13. [Google Scholar] [CrossRef]
- Dellacqua, Z.; Di Biagio, C.; Martini, A.; Mattei, F.; Rakaj, A.; Williams, J.; Fabris, A.; Izquierdo, M.; Boglione, C. Distinguishing the Impacts of Rearing Density versus Tank Volume on the Skeletal Quality and Development of Gilthead Seabream (Sparus aurata)during the Hatchery Phase. Animals 2023. submitted. [Google Scholar]
- Meena, D.; Sahoo, A.; Jayant, M.; Sahu, N.; Srivastava, P.; Swain, H.; Behera, B.; Satvik, K.; Das, B. Bioconversion of Terminalia arjuna bark powder into a herbal feed for Labeo rohita: Can it be a sustainability paradigm for Green Fish production? Anim. Feed. Sci. Technol. 2021, 284, 115132. [Google Scholar] [CrossRef]
- Witten, P.E.; Hansen, A.; Hall, B.K. Features of mono- and multinucleated bone resorbing cells of the zebrafish Danio rerio and their contribution to skeletal development, remodeling, and growth. J. Morphol. 2001, 250, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, K.X. On the criterion that a given system of deviations from the probable in the case of a correlated system of variables is such that it can be reasonably supposed to have arisen from random sampling. Lond. Edinb. Dublin Philos. Mag. 1900, 50, 157–175. [Google Scholar] [CrossRef]
- Sfakianakis, D.; Georgakopoulou, E.; Papadakis, I.; Divanach, P.; Kentouri, M.; Koumoundouros, G. Environmental determinants of haemal lordosis in European sea bass, Dicentrarchus labrax (Linnaeus, 1758). Aquaculture 2006, 254, 54–64. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Loy, B.A.; Boglione, C.; Cataudella, S. Geometric morphometrics and morpho-anatomy: A combined tool in the study of sea bream (Sparus aurata, sparidae) shape. J. Appl. Ichthyol. 1999, 15, 104–110. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Slice, D. Extensions of the Procrustes Method for the Optimal Superimposition of Landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Costa, C.; Tibaldi, E.; Pasqualetto, L.; Loy, A. Morphometric comparison of the cephalic region of cultured Acipenser baerii (Brandt, 1869), Acipenser naccarii (Bonaparte, 1836) and their hybrid*. J. Appl. Ichthyol. 2006, 22, 8–14. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier Sci.: Amsterdam, The Netherlands, 1998; 853p. [Google Scholar]
- Bookstein, F. Principal warps: Thin-plate splines and the decomposition of deformations. IEEE Trans. Pattern Anal. Mach. Intell. 1989, 11, 567–585. [Google Scholar] [CrossRef]
- Loy, A.; Mariani, L.; Bertelletti, M.; Tunesi, L. Visualizing Allometry: Geometric Morphometrics in the Study of Shape Changes in the Early Stages of the Two-Banded Sea Bream, Diplodus vulgaris (Perciformes, Sparidae). J. Morphol. 1998, 237, 137–146. [Google Scholar] [CrossRef]
- Colangelo, P.; Ventura, D.; Piras, P.; Bonaiuti, J.P.G.; Ardizzone, G. Are developmental shifts the main driver of phenotypic evolution in Diplodus spp. (Perciformes: Sparidae)? BMC Evol. Biol. 2019, 19, 106. [Google Scholar] [CrossRef]
- Kerschbaumer, M.; Sturmbauer, C. The Utility of Geometric Morphometrics to Elucidate Pathways of Cichlid Fish Evolution. Int. J. Evol. Biol. 2011, 2011, 290245. [Google Scholar] [CrossRef]
- Costa, C.; Pasqualetto, L.; Tibaldi, E.; Loy, A. Shape changes and differences in the cranial region in Acipenser baerii and A. naccarii of different size. Ital. J. Zool. 2004, 71, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Loy, A.; Boglione, C.; Gagliardi, F.; Ferrucci, L.; Cataudella, S. Geometric morphometrics and internal anatomy in sea bass shape analysis (Dicentrarchus labrax L., Moronidae). Aquaculture 2000, 186, 33–44. [Google Scholar] [CrossRef]
- Yiannis, G.R.; Katerina, K.K.; Alexios, R.; Evagelos, D.; George, N.K. Comparative morphology of wild, farmed and hatchery-released gilthead sea bream (Sparus aurata) in western Greece. Int. J. Fish. Aquac. 2011, 3, 1–9. [Google Scholar]
- Russo, T.; Costa, C.; Cataudella, S. Correspondence between shape and feeding habit changes throughout ontogeny of gilthead sea bream Sparus aurata L., 1758. J. Fish Biol. 2007, 71, 629–656. [Google Scholar] [CrossRef]
- Koumoundouros, G. Morpho-anatomical abnormalities in Mediterranean marine aquaculture. In Recent Advances in Aquaculture Research; Transworld Research Network: Trivandrum, India, 2010; Volume 661, pp. 125–148. [Google Scholar]
- Fragkoulis, S.; Koumoundouros, G. Simple morphometrics for predicting lordosis-induced deviations of body shape in reared Gilthead seabream (Sparus aurata L.). J. Fish Dis. 2021, 44, 1265–1267. [Google Scholar] [CrossRef]
- Printzi, A.; Fragkoulis, S.; Dimitriadi, A.; Keklikoglou, K.; Arvanitidis, C.; Witten, P.E.; Koumoundouros, G. Exercise-induced lordosis in zebrafish Danio rerio (Hamilton, 1822). J. Fish Biol. 2021, 98, 987–994. [Google Scholar] [CrossRef]
- Fragkoulis, S.; Printzi, A.; Geladakis, G.; Katribouzas, N.; Koumoundouros, G. Recovery of haemal lordosis in Gilthead seabream (Sparus aurata L.). Sci. Rep. 2019, 9, 9832. [Google Scholar] [CrossRef]
- Fragkoulis, S.; Kourkouta, C.; Geladakis, G.; Printzi, A.; Glaropoulos, A.; Koumoundouros, G. Recovery of Haemal Lordosis in European Seabass Dicentrarchus labrax (Linnaeus 1758). Aquac. J. 2022, 2, 1–12. [Google Scholar] [CrossRef]
- Chatain, B.; Dewavrin, G. Influence des anomalies de développement de la vessie natatoire sur la mortalité de Dicentrarchus labrax au cours du sevrage. Aquaculture 1989, 78, 55–61. [Google Scholar] [CrossRef]
- Chatain, B. Abnormal swimbladder development and lordosis in sea bass (Dicentrarchus labrax) and sea bream (Sparus auratus). Aquaculture 1994, 119, 371–379. [Google Scholar] [CrossRef]
- Kitajima, C.; Watanabe, T.; Tsukashima, Y.; Fujita, S. Lordotic Deformation and Abnormal Development of Swim Bladders in Some Hatchery-Bred Marine Physoclistous Fish in Japan. J. World Aquac. Soc. 1994, 25, 64–77. [Google Scholar] [CrossRef]
- Boglione, C.; Marino, G.; Fusari, A.; Ferreri, A.; Finoia, M.G.; Cataudella, S. Skeletal anomalies in Dicentrarchus labrax juveniles selected for functional swimbladder. ICES Mar. Sci. Symp. 1995, 201, 163–169. [Google Scholar]
- Andrades, J.; Becerra, J.; Fernández-Llebrez, P. Skeletal deformities in larval, juvenile and adult stages of cultured gilthead sea bream (Sparus aurata L.). Aquaculture 1996, 141, 1–11. [Google Scholar] [CrossRef]
- Divanach, P.; Papandroulakis, N.; Anastasiadis, P.; Koumoundouros, G.; Kentouri, M. Effect of water currents on the development of skeletal deformities in sea bass (Dicentrarchus labrax L.) with functional swimbladder during postlarval and nursery phase. Aquaculture 1997, 156, 145–155. [Google Scholar] [CrossRef]
- Kihara, M.; Ogata, S.; Kawano, N.; Kubota, I.; Yamaguchi, R. Lordosis induction in juvenile red sea bream, Pagrus major, by high swimming activity. Aquaculture 2002, 212, 149–158. [Google Scholar] [CrossRef]
- Kranenbarg, S.; Waarsing, J.H.; Muller, M.; Weinans, H.; van Leeuwen, J.L. Lordotic vertebrae in sea bass (Dicentrarchus labrax L.) are adapted to increased loads. J. Biomech. 2005, 38, 1239–1246. [Google Scholar] [CrossRef]
- Boursiaki, V.; Theochari, C.; Zaoutsos, S.P.; Mente, E.; Vafidis, D.; Apostologamvrou, C.; Berillis, P. Skeletal Deformity of Scoliosis in Gilthead Seabreams (Sparus aurata): Association with Changes to Calcium-Phosphor Hydroxyapatite Salts and Collagen Fibers. Water 2019, 11, 257. [Google Scholar] [CrossRef]
- Blasco, J.; Moya, A.; Millán-Cubillo, A.; Vélez, E.J.; Capilla, E.; Pérez-Sánchez, J.; Gutiérrez, J.; Borrás, J.F. Growth-promoting effects of sustained swimming in fingerlings of gilthead sea bream (Sparus aurata L.). J. Comp. Physiol. B 2015, 185, 859–868. [Google Scholar] [CrossRef]
- Moya, A.; Torrella, J.; Fernández-Borràs, J.; Rizo-Roca, D.; Millán-Cubillo, A.; Vélez, E.; Arcas, A.; Gutiérrez, J.; Blasco, J. Sustained swimming enhances white muscle capillarisation and growth by hyperplasia in gilthead sea bream (Sparus aurata) fingerlings. Aquaculture 2019, 501, 397–403. [Google Scholar] [CrossRef]
- Ibarz, A.; Felip, O.; Fernández-Borràs, J.; Martín-Pérez, M.; Blasco, J.; Torrella, J.R. Sustained swimming improves muscle growth and cellularity in gilthead sea bream. J. Comp. Physiol. B 2011, 181, 209–217. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.J.; Sánchez, B.; Barroso, F.G.; Toniolo, M.; Trenzado, C.E.; Rus, A.S. Effects of rearing conditions on behavioural responses, social kinetics and physiological parameters in gilthead sea bream Sparus aurata. Appl. Anim. Behav. Sci. 2017, 197, 120–128. [Google Scholar] [CrossRef]
- Beamish, F.W.H. Swimming Capacity. Fish Physiol. 1978, 7, 101–187. [Google Scholar]
- Gui, F.; Wang, P.; Wu, C. Evaluation approaches of fish swimming performance. Agric. Sci. 2014, 05, 106–113. [Google Scholar] [CrossRef]
- Kozhara, A.V. Phenotypic Variance of Bilateral Characters as an Indicator of Genetic and Environmental Conditions in Bream Abramis brama (L.) (Pisces, Cyprinidae) Populations. J. Appl. Ichthyol. 1994, 10, 167–181. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Most severe | <137.9° |
| Quite severe | 138.0°–144.9° |
| Severe | 145.0°–151.9° |
| Moderate | 152.0°–158.9° |
| Slight | 159.0°–165.9° |
| Relatively mild | 166.0°–172.9° |
| Normal | 173.0°–180° |
| Lots | T0 | LD1000 | MD1000 | HD1000 | LD500 | MD500 | HD500 |
|---|---|---|---|---|---|---|---|
| Number of observed specimens | 161 | 140 | 115 | 197 | 124 | 96 | 181 |
| % of individuals with at least one anomaly | 79 | 63 | 69 | 80 | 69 | 75 | 88 |
| Anomalies charge (n. of anomalies/affected fish) | 3 | 2 | 2 | 2 | 2 | 2 | 2 |
| Observed types of anomalies | 21 | 18 | 23 | 23 | 17 | 21 | 23 |
| Pairwise Post Hoc | p-Values |
|---|---|
| LD1000 vs. HD1000 | 0.002 |
| MD1000 vs. HD1000 | 0.002 |
| HD1000 vs. LD500 | 0.002 |
| HD1000 vs. MD500 | 0.02 |
| LD500 vs. HD500 | 0.02 |
| MD1000 | HD1000 | LD500 | MD500 | HD500 | |
|---|---|---|---|---|---|
| LD1000 | 6.7184 | 6.5169 | 4.0499 | 11.3400 | 8.7823 |
| MD1000 | 2.5247 | 9.1422 | 3.6365 | 3.9081 | |
| HD1000 | 11.321 | 6.9757 | 2.9881 | ||
| LD500 | 11.575 | 14.898 | |||
| MD500 | 5.905 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dellacqua, Z.; Di Biagio, C.; Costa, C.; Pousão-Ferreira, P.; Ribeiro, L.; Barata, M.; Gavaia, P.J.; Mattei, F.; Fabris, A.; Izquierdo, M.; et al. Distinguishing the Effects of Water Volumes versus Stocking Densities on the Skeletal Quality during the Pre-Ongrowing Phase of Gilthead Seabream (Sparus aurata). Animals 2023, 13, 557. https://doi.org/10.3390/ani13040557
Dellacqua Z, Di Biagio C, Costa C, Pousão-Ferreira P, Ribeiro L, Barata M, Gavaia PJ, Mattei F, Fabris A, Izquierdo M, et al. Distinguishing the Effects of Water Volumes versus Stocking Densities on the Skeletal Quality during the Pre-Ongrowing Phase of Gilthead Seabream (Sparus aurata). Animals. 2023; 13(4):557. https://doi.org/10.3390/ani13040557
Chicago/Turabian StyleDellacqua, Zachary, Claudia Di Biagio, Corrado Costa, Pedro Pousão-Ferreira, Laura Ribeiro, Marisa Barata, Paulo J. Gavaia, Francesco Mattei, Andrea Fabris, Marisol Izquierdo, and et al. 2023. "Distinguishing the Effects of Water Volumes versus Stocking Densities on the Skeletal Quality during the Pre-Ongrowing Phase of Gilthead Seabream (Sparus aurata)" Animals 13, no. 4: 557. https://doi.org/10.3390/ani13040557