
Ovulation Induced by Intrauterine Seminal Plasma Increases Total Protein, PGE2, IL-8, and IL-1β in Uterine Fluid of Llamas (Lama glama)
 and
and 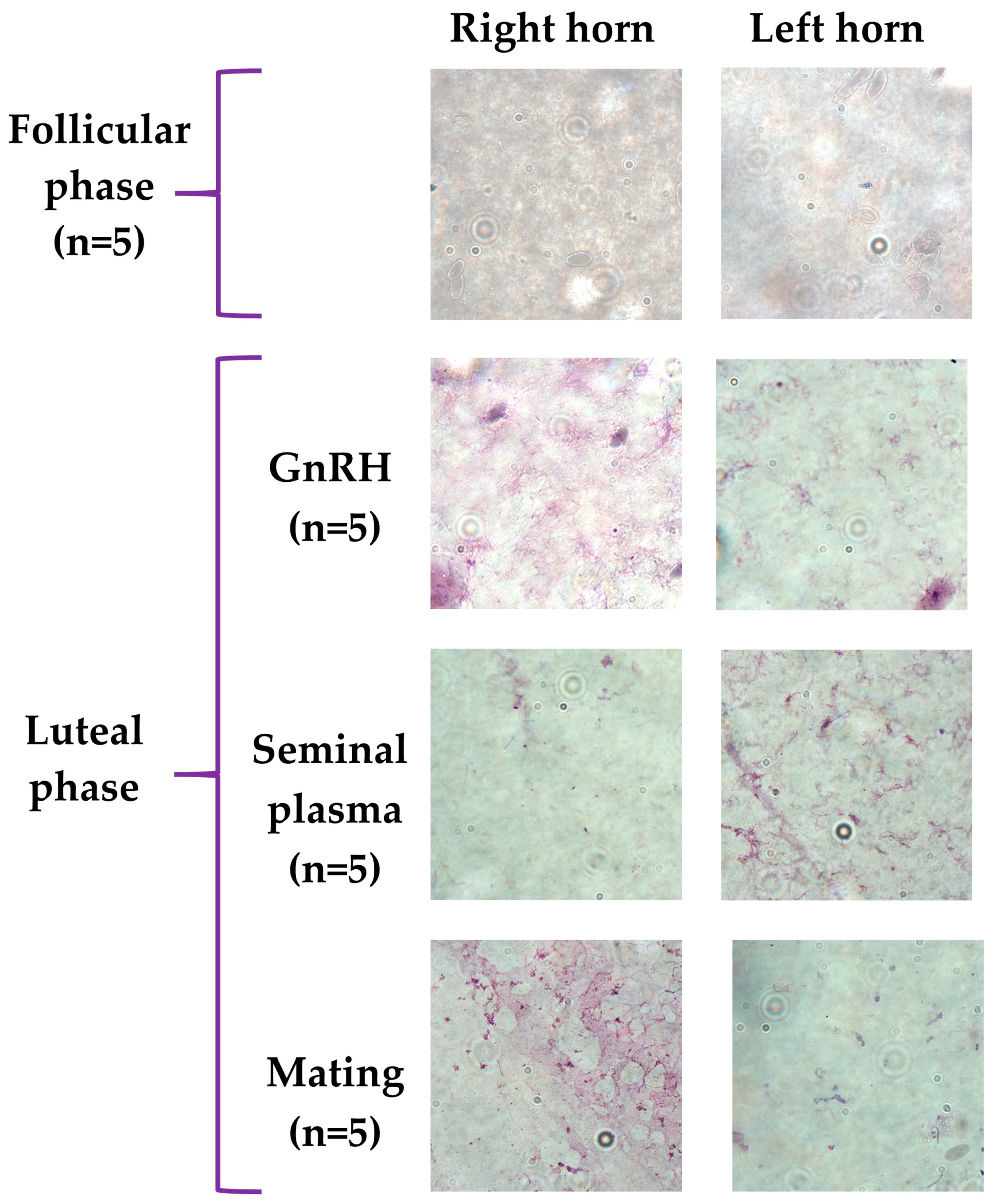{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Experimental Design
- i
- Follicular phase: (n = 5) based on previous studies [51,53] the presence of a follicle ≥ 8 mm in diameter for 3 consecutive days is an indicator of the follicular phase and it is associated with sexual receptivity. Ovulation will not occur unless females are given ovulatory stimuli (hormones or mating). The females of this group were submitted to uterine flushing at the third day of scanning corroborating the presence of the dominant follicle;
- ii
- Luteal phase induced by GnRH: (n = 5) females were given an i.m. dose of 50 ug analogue of GnRH gonadorelin acetate Gonasyl® (Syva, León, Spain);
- iii
- Luteal phase induced by intrauterine infusion of seminal plasma: (n = 5) females were intrauterine-infused with 5 mL of llama seminal plasma with an insemination catheter accoupled to a 20 mL syringe using the transrectal palpation technique;
- iv
- Luteal phase induced by mating: (n = 4) females were mated with fertile males, successful copulation was confirmed by the introduction of the penis into the female genital tract, with a minimum duration of the copulation of 25 min.
2.3. Llama Male Semen Collection for Preparation of Seminal Plasma
2.4. Uterine Fluid Collection by Non-Surgical Uterine Flushing
2.5. Cytology Analyses of Uterine Fluid
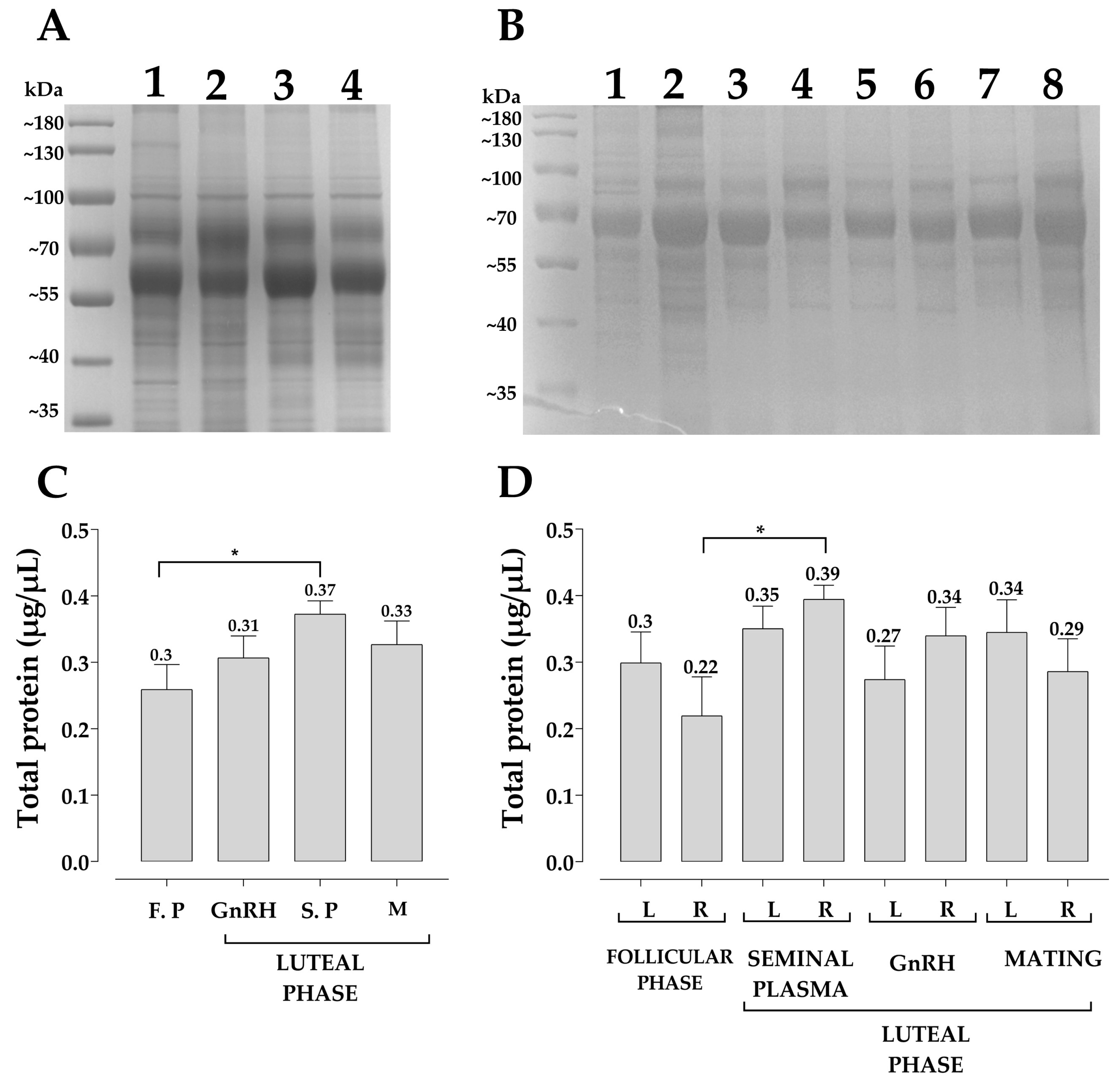
2.6. Protein Quantification and Characterization Pattern of Uterine Fluid
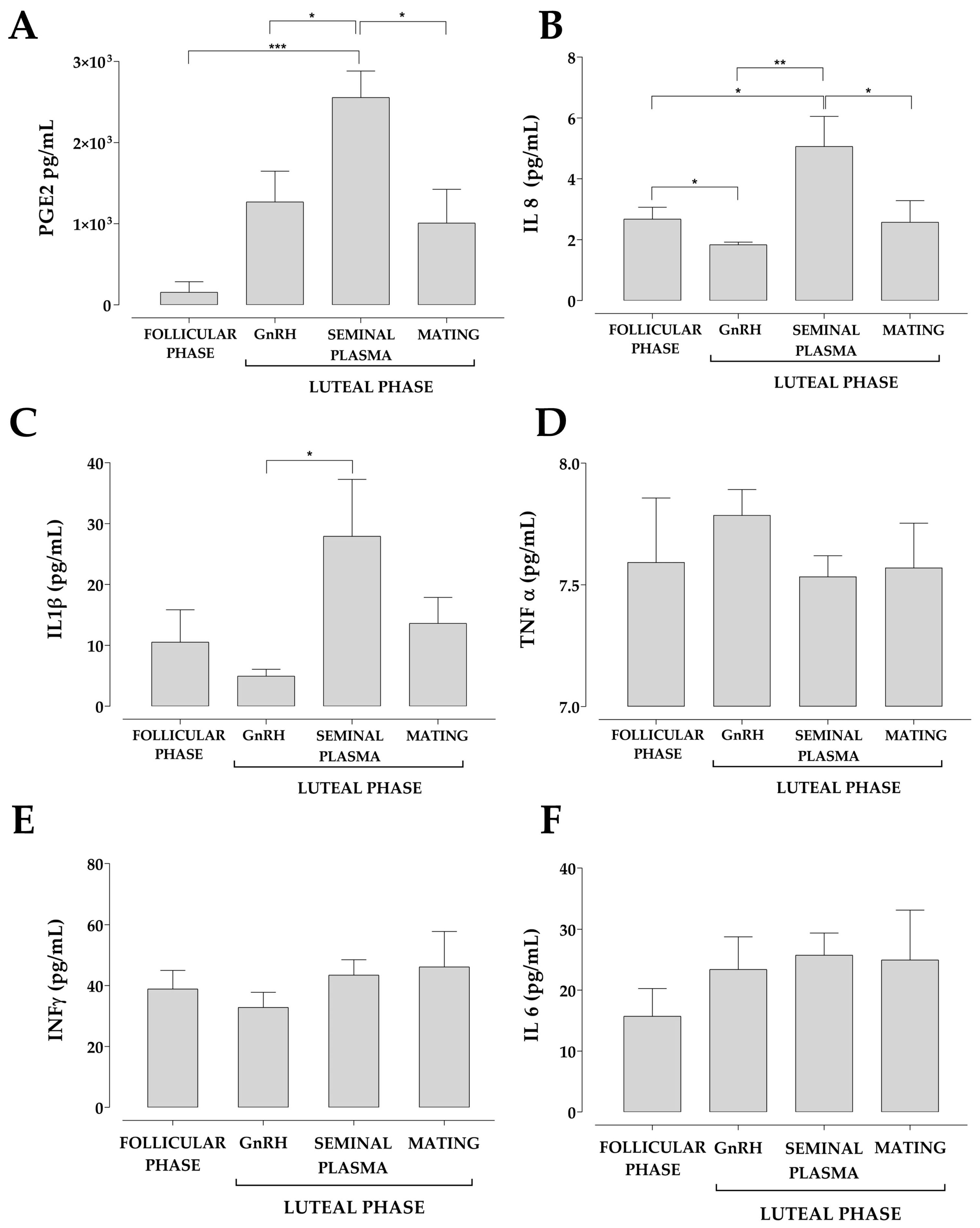
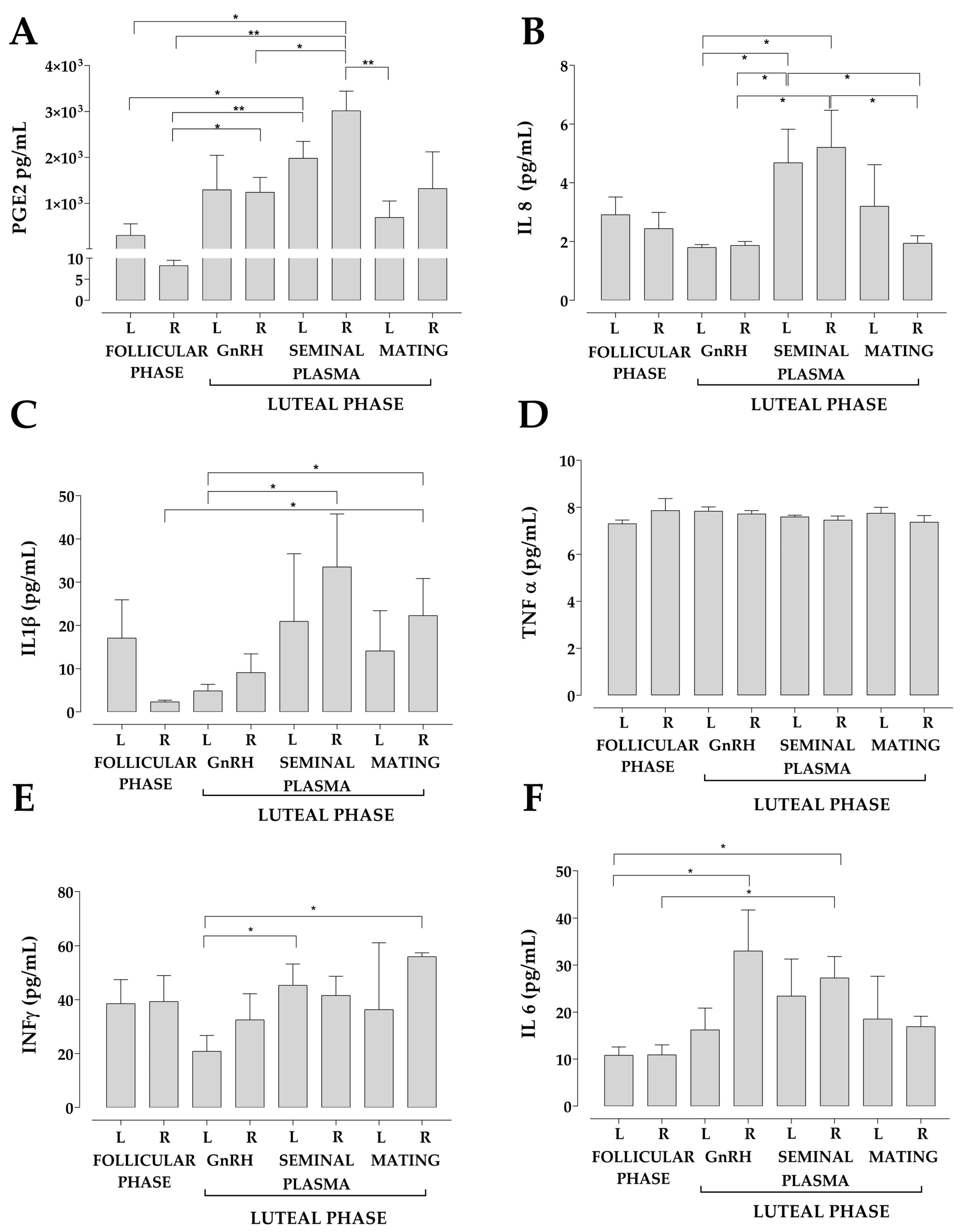
2.7. Measurements of IL-1β, IL-6, IL-8, IFN γ, TNF-α and PGE2 Concentration in the Uterine Fluid
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agostinis, C.; Mangogna, A.; Bossi, F.; Ricci, G.; Kishore, U.; Bulla, R. Uterine immunity and microbiota: A shifting paradigm. Front. Immunol. 2019, 10, 2387. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, S.; Di Nicuolo, F.; Pontecorvi, A.; Gratta, M.; Scambia, G.; Di Simone, N. Endometrial microbes and microbiome: Recent insights on the inflammatory and immune ‘players’ of the human endometrium. Am. J. Reprod. Immunol. 2018, 80, e13065. [Google Scholar] [CrossRef]
- García-Velasco, J.A.; Arici, A. Chemokines and human reproduction. Fertil. Steril. 1999, 71, 983–993. [Google Scholar] [CrossRef]
- Duffy, D.M.; Ko, C.; Jo, M.; Brannstrom, M.; Curry, T.E. Ovulation: Parallels with inflammatory processes. Endocr. Rev. 2019, 40, 369–416. [Google Scholar] [CrossRef]
- Dekel, N.; Gnainsky, Y.; Granot, I.; Mor, G. Inflammation and implantation. Am. J. Reprod. Immunol. 2009, 63, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and pregnancy: The role of the immune system at the implantation site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef]
- Robertson, S.A. Control of the immunological environment of the uterus. Rev. Reprod. 2000, 5, 164–174. [Google Scholar] [CrossRef]
- Wira, C.R.; Rodriguez-Garcia, M.; Patel, M.V. The role of sex hormones in immune protection of the female reproductive tract. Nat. Rev. Immunol. 2015, 15, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Katila, T. Post-mating Inflammatory Responses of the Uterus. Reprod. Domest. Anim. 2012, 47, 31–41. [Google Scholar] [CrossRef]
- Recuero, S.; Sánchez, J.M.; Mateo-Otero, Y.; Bagés-Arnal, S.; McDonald, M.; Behura, S.K.; Spencer, T.E.; Kenny, D.A.; Yeste, M.; Lonergan, P.; et al. Mating to Intact, but Not Vasectomized, Males Elicits Changes in the Endometrial Transcriptome: Insights From the Bovine Model. Front. Cell Dev. Biol. 2020, 8, 547. [Google Scholar] [CrossRef]
- Martins, T.; Pugliesi, G.; Sponchiado, M.; Gonella-Diaza, A.M.; Ojeda-Rojas, O.A.; Rodriguez, F.D.; Ramos, R.S.; Basso, A.C.; Binelli, M. Perturbations in the uterine luminal fluid composition are detrimental to pregnancy establishment in cattle. J. Anim. Sci. Biotechnol. 2018, 9, 70. [Google Scholar] [CrossRef]
- Argañaraz, M.E.; Apichela, S.A.; Zampini, R.; Vencato, J.; Stelletta, C. Biochemical and Protein Profile of Alpaca (Vicugna pacos) Uterine Horn Fluid During Early Pregnancy. Reprod. Domest. Anim. 2015, 50, 121–128. [Google Scholar] [CrossRef]
- Hugentobler, S.A.; Humpherson, P.G.; Leese, H.J.; Sreenan, J.M.; Morris, D.G. Energy substrates in bovine oviduct and uterine fluid and blood plasma during the oestrous cycle. Mol. Reprod. Dev. 2008, 75, 496–503. [Google Scholar] [CrossRef]
- Tríbulo, P.; Siqueira, L.G.B.; Oliveira, L.J.; Scheffler, T.; Hansen, P.J. Identification of potential embryokines in the bovine reproductive tract. J. Dairy Sci. 2018, 101, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Hempstock, J.; Cindrova-Davies, T.; Jauniaux, E.; Burton, G.J. Endometrial glands as a source of nutrients, growth factors and cytokines during the first trimester of human pregnancy: A morphological and immunohistochemical study. Reprod. Biol. Endocrinol. 2004, 2, 58. [Google Scholar] [CrossRef]
- Ametzazurra, A.; Matorras, R.; Garcia-Velasco, J.A.; Prieto, B.; Simon, L.; Martinez, A.; Nagore, D. Endometrial fluid is a specific and non-invasive biological sample for protein biomarker identification in endometriosis. Hum. Reprod. 2009, 24, 954–965. [Google Scholar] [CrossRef]
- Paulesu, L.; Bhattacharjee, J.; Bechi, N.; Romagnoli, R.; Jantra, S.; Ietta, F. Pro-inflammatory Cytokines in Animal and Human Gestation. Curr. Pharm. Des. 2010, 16, 3601–3615. [Google Scholar] [CrossRef]
- Zhu, J.; Mayr, D.; Kuhn, C.; Mahner, S.; Jeschke, U.; von Schönfeldt, V. Prostaglandin E2 receptor EP1 in healthy and diseased human endometrium. Histochem. Cell Biol. 2018, 149, 153–160. [Google Scholar] [CrossRef]
- Shimizu, Y.; Mita, S.; Takeuchi, T.; Notsu, T.; Mizuguchi, K.; Kyo, S. Dienogest, a synthetic progestin, inhibits prostaglandin E2 production and aromatase expression by human endometrial epithelial cells in a spheroid culture system. Steroids 2011, 76, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Sacco, K.; Portelli, M.; Pollacco, J.; Schembri-Wismayer, P.; Calleja-Agius, J. The role of prostaglandin E2 in endometriosis. Gynecol. Endocrinol. 2011, 28, 134–138. [Google Scholar] [CrossRef]
- Arici, A.; Head, J.R.; MacDonald, P.C.; Casey, M.L. Regulation of interleukin-8 gene expression in human endometrial cells in culture. Mol. Cell. Endocrinol. 1993, 94, 195–204. [Google Scholar] [CrossRef]
- Kelly, P.; Meade, K.G.; O’Farrelly, C. Non-canonical inflammasome-mediated IL-1β production by primary endometrial epithelial and stromal fibroblast cells is NLRP3 and caspase-4 dependent. Front. Immunol. 2019, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Argilés, J.M.; Carbó, N.; López-Soriano, F.J. TNF and pregnancy: The paradigm of a complex interaction. Cytokine Growth Factor Rev. 1997, 8, 181–188. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-γ: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef]
- Gill, T.J.; Coulam, C.B.; Stern, J.J. Immunology of ovarian failure. Am. J. Reprod. Immunol. 1991, 25, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Kim, J.Y.; Park, J.E.; Yoon, Y.-D.; Tsang, B.K.; Kim, J.-M. Induction of Fas-Mediated Apoptosis by Interferon-γ is Dependent on Granulosa Cell Differentiation and Follicular Maturation in the Rat Ovary. Dev. Reprod. 2016, 20, 315–329. [Google Scholar] [CrossRef]
- Terranova, P.F.; Rice, V.M. Review: Cytokine involvement in ovarian processes. Am. J. Reprod. Immunol. 1997, 37, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Terranova, P.F. Potential roles of tumor necrosis factor-α in follicular development, ovulation, and the life span of the corpus luteum. Domest. Anim. Endocrinol. 1997, 14, 1–15. [Google Scholar] [CrossRef]
- Niringiyumukiza, J.D.; Cai, H.; Xiang, W. Prostaglandin E2 involvement in mammalian female fertility: Ovulation, fertilization, embryo development and early implantation. Reprod. Biol. Endocrinol. 2018, 16, 43. [Google Scholar] [CrossRef]
- Seyam, E.; Hefzy, E. Tumor necrosis factor alpha versus LH and androstendione as a reliable predictor of spontaneous ovulation after laparoscopic ovarian drilling for women with clomiphene citrate resistance polycystic ovarian disease. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 222, 126–133. [Google Scholar] [CrossRef]
- Hales, H.A.; Peterson, C.M.; Mitchell, M.D.; Jones, K.P.; Hatasaka, H.H.; Poulson, A.M. Tumor Necrosis Factor-Alpha Inhibits Ovulation and Steroidogenesis, but Not Prostaglandin Production in the Perfused Rat Ovary. J. Soc. Gynecol. Investig. 1994, 1, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kuwahara, A.; Taniguchi, Y.; Yamasaki, M.; Tanaka, Y.; Mukai, Y.; Yamashita, M.; Matsuzaki, T.; Yasui, T.; Irahara, M. Tumor necrosis factor alpha inhibits ovulation and induces granulosa cell death in rat ovaries. Reprod. Med. Biol. 2015, 14, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Arici, A.; Oral, E.; Bukulmez, O.; Buradagunta, S.; Engin, O.; Olive, D.L. Interleukin-8 expression and modulation in human preovulatory follicles and ovarian cells. Endocrinology 1996, 137, 3762–3769. [Google Scholar] [CrossRef]
- Ujioka, T.; Matsukawa, A.; Tanaka, N.; Matsuura, K.; Yoshinaga, M.; Okamura, H. Interleukin-8 as an essential factor in the human chorionic gonadotropin-induced rabbit ovulatory process: Interleukin-8 induces neutrophil accumulation and activation in ovulation. Biol. Reprod. 1998, 58, 526–530. [Google Scholar] [CrossRef]
- Shimizu, T.; Kaji, A.; Murayama, C.H.; Magata, F.; Shirasuna, K.; Wakamiya, K.; Okuda, K.; Miyamoto, T.; Shimizu, A.; Kaji, A.; et al. Miyamoto. Effects of interleukin-8 on estradiol and progesterone production by bovine granulosa cells from large follicles and progesterone production by luteinizing granulosa cells in culture. Cytokine 2012, 57, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Zhou, C.; Wei, C.-J.; Zhao, S.; Sun, F.; Zhou, H.; Xu, W.; Liu, J.; Yang, C.; Wu, L.; et al. Evaluation of in vitro fertilization outcomes using interleukin-8 in culture medium of human preimplantation embryos. Fertil. Steril. 2017, 107, 649–656. [Google Scholar] [CrossRef]
- Stassi, A.F.; Baravalle, M.E.; Belotti, E.M.; Rey, F.; Gareis, N.C.; Díaz, P.U.; Rodríguez, F.M.; Leiva, C.J.; Ortega, H.H.; Salvetti, N.R. Altered expression of IL-1β IL-1RI, IL-1RII, IL-1RA and IL-4 could contribute to anovulation and follicular persistence in cattle. Theriogenology 2018, 110, 61–73. [Google Scholar] [CrossRef]
- Caballero-Campo, P. Hormonal and embryonic regulation of chemokines IL-8, MCP-1 and RANTES in the human endometrium during the window of implantation. Mol. Hum. Reprod. 2002, 8, 375–384. [Google Scholar] [CrossRef]
- Freis, A.; Germeyer, A.; Jauckus, J.; Capp, E.; Strowitzki, T.; Zorn, M.; Weber, A.M. Endometrial expression of receptivity markers subject to ovulation induction agents. Arch. Gynecol. Obstet. 2019, 300, 1741–1750. [Google Scholar] [CrossRef]
- Brännström, V. The Ovary. In Potential Role of Cytokines in Ovarian Physiology: The Case for Interleukin-1, 2nd ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2004; pp. 261–271. [Google Scholar]
- Simón, C.; Mercader, A.; Gimeno, M.J.; Pellicer, A. The interleukin-1 system and human implantation. Am. J. Reprod. Immunol. 1997, 37, 64–72. [Google Scholar] [CrossRef]
- Sequeira, K.; Espejel-Núñez, A.; Vega-Hernández, E.; Molina-Hernández, A.; Grether-González, P. An increase in IL-1β concentrations in embryo culture-conditioned media obtained by in vitro fertilization on day 3 is related to successful implantation. J. Assist. Reprod. Genet. 2015, 32, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Kreines, F.M.; Nasioudis, D.; Minis, E.; Irani, M.; Witkin, S.S.; Spandorfer, S. IL-1β predicts IVF outcome: A prospective study. J. Assist. Reprod. Genet. 2018, 35, 2031–2035. [Google Scholar] [CrossRef]
- Chatterjee, A.; Guchhait, R.; Maity, S.; Mukherjee, D.; Pramanick, K. Functions of interleukin-6 in ovulation of female climbing perch, Anabas testudineus. Anim. Reprod. Sci. 2019, 219, 106528. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, X.; Wang, L.; Liu, Y.; Li, N.; Li, M.; Chen, L.; Liu, Y.; Yao, Y. Interleukin-6 regulates expression of Fos and Jun genes to affect the development of mouse preimplantation embryos. J. Obstet. Gynaecol. Res. 2018, 44, 253–262. [Google Scholar] [CrossRef]
- Sakumoto, R.; Komatsu, T.; Kasuya, E.; Saito, T.; Okuda, K. Expression of mRNAs for interleukin-4, interleukin-6 and their receptors in porcine corpus luteum during the estrous cycle. Domest. Anim. Endocrinol. 2006, 31, 246–257. [Google Scholar] [CrossRef]
- Alijotas-Reig, J.; Esteve-Valverde, E.; Ferrer-Oliveras, R.; Llurba, E.; Gris, J.M. Tumor Necrosis Factor-Alpha and Pregnancy: Focus on Biologics. An Updated and Comprehensive Review. Clin. Rev. Allergy Immunol. 2017, 53, 40–53. [Google Scholar] [CrossRef]
- Brown, B.W. A review on reproduction in South American camelids. Anim. Reprod. Sci. 2000, 58, 169–195. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.L.; Tibary, A. Reproduction in female South American camelids: A review and clinical observations. Small Rumin. Res. 2006, 61, 259–281. [Google Scholar] [CrossRef]
- Bravo, P.W. Reproductive endocrinology of llamas and alpacas. Vet. Clin. N. Am. Food Anim. Pract. 1994, 10, 265–279. [Google Scholar] [CrossRef]
- Ratto, M.H.; Urra, F.; Silva, M. Laterality of Ovulation and Presence of the Embryo Do Not Affect Uterine Horn Blood Flow During the First Month of Gestation in Llamas. Front. Vet. Sci. 2020, 7, 598117. [Google Scholar] [CrossRef]
- Picha, Y.; Tibary, A.; Memon, M.; Kasimanickam, R.; Sumar, J. Chronology of early embryonic development and embryo uterine migration in alpacas. Theriogenology 2013, 79, 702–708. [Google Scholar] [CrossRef]
- Ratto, M.; Huanca, W.; Singh, J.; Adams, G.P. Comparison of the effect of natural mating, LH, and GnRH on interval to ovulation and luteal function in llamas. Anim. Reprod. Sci. 2006, 91, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Adams, G.P.; Ratto, M.H.; Huanca, W.; Singh, J. Ovulation-Inducing Factor in the Seminal Plasma of Alpacas and Llamas. Biol. Reprod. 2005, 73, 452–457. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, C.J.; Kim, D.-J.; Kang, J. Immune Cells in the Female Reproductive Tract. Immune Netw. 2015, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Akbalik, M.E.; Liman, N.; Sagsoz, H.; Saruhan, B.G. Tissue distribution of some immune cells in bovine reproductive tract during follicular and luteal phase. Microsc. Res. Tech. 2018, 81, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Kaushic, C.; Frauendorf, E.; Rossoll, R.M.; Richardson, J.M.; Wira, C.R. Influence of the estrous cycle on the presence and distribution of immune cells in the rat reproductive tract. Am. J. Reprod. Immunol. 1998, 39, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Cronin, J.G.; Healey, G.D.; Gabler, C.; Heuwieser, W.; Streyl, D.; Bromfield, J.J.; Miyamoto, A.; Fergani, C.; Dobson, H. Innate immunity and inflammation of the bovine female reproductive tract in health and disease. Reproduction 2014, 148, R41–R51. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Owens, S.-E.; Turner, M.L. Innate immunity and the sensing of infection, damage and danger in the female genital tract. J. Reprod. Immunol. 2017, 119, 67–73. [Google Scholar] [CrossRef]
- Barlund, C.S.; Carruthers, T.D.; Waldner, C.L.; Palmer, C.W. A comparison of diagnostic techniques for postpartum endometritis in dairy cattle. Theriogenology 2008, 69, 714–723. [Google Scholar] [CrossRef]
- Cazales, N.; Estradé, M.J.; Pereyra, F.; Fiala-Rechsteiner, S.M.; Mattos, R.C. Sperm transport and endometrial inflammatory response in mares after artificial insemination with cryopreserved spermatozoa. Theriogenology 2020, 158, 180–187. [Google Scholar] [CrossRef]
- Ibrahim, L.A.; Rizo, J.A.; Fontes, P.L.P.; Lamb, G.C.; Bromfield, J.J. Seminal plasma modulates expression of endometrial inflammatory meditators in the bovine. Biol. Reprod. 2019, 100, 660–671. [Google Scholar] [CrossRef]
- Nongbua, T.; Guo, Y.; Ntallaris, T.; Rubér, M.; Rodriguez-Martinez, H.; Humblot, P.; Morrell, J.M. Bull seminal plasma stimulates in vitro production of TGF-β, IL-6 and IL-8 from bovine endometrial epithelial cells, depending on dose and bull fertility. J. Reprod. Immunol. 2020, 142, 103179. [Google Scholar] [CrossRef] [PubMed]
- Shahnazi, M.; Nouri, M.; Mohaddes, G.; Latifi, Z.; Fattahi, A.; Mohammadi, M. Prostaglandin E Pathway in Uterine Tissue During Window of Preimplantation in Female Mice Mated With Intact and Seminal Vesicle-Excised Male. Reprod. Sci. 2018, 25, 550–558. [Google Scholar] [CrossRef]
- Nongbua, T.; Guo, Y.; Edman, A.; Humblot, P.; Morrell, J.M. Effect of bovine seminal plasma on bovine endometrial epithelial cells in culture. Reprod. Domest. Anim. 2018, 53, 85–92. [Google Scholar] [CrossRef]
- Scott, J.L.; Ketheesan, N.; Summers, P.M. Spermatozoa and seminal plasma induce a greater inflammatory response in the ovine uterus at oestrus than dioestrus. Reprod. Fertil. Dev. 2009, 21, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.H.; Li, Z.Y.; Li, D.D.; Fang, W.N.; Liu, H.Y.; Yang, D.D.; Meng, C.Y.; Yang, Y.; Peng, J.P. Seminal plasma induces inflammation in the uterus through the γδ T/IL-17 pathway. Sci. Rep. 2016, 6, 25118. [Google Scholar] [CrossRef]
- Gutsche, S.; von Wolff, M.; Strowitzki, T.; Thaler, C.J. Seminal plasma induces mRNA expression of IL-1β, IL-6 and LIF in endometrial epithelial cells in vitro. Mol. Hum. Reprod. 2003, 9, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Taylor, U.; Zerbe, H.; Seyfert, H.M.; Rath, D.; Baulain, U.; Langner, K.F.A.; Schuberth, H.J. Porcine spermatozoa inhibit post-breeding cytokine induction in uterine epithelial cells in vivo. Anim. Reprod. Sci. 2009, 115, 279–289. [Google Scholar] [CrossRef]
- Ishihara, O.; Sullivan, M.; Elder, M. Differences of metabolism of prostaglandin E2 and F2 alpha by decidual stromal cells and macrophages in culture. Eicosanoids 1991, 4, 203–207. [Google Scholar]
- Villarete, L.H.; Remick, D.G. Transcriptional and post-transcriptional regulation of interleukin-8. Am. J. Pathol. 1996, 149, 1685–1693. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1865258/ (accessed on 14 January 2023).
- Wu, M.H.; Lu, C.W.; Chuang, P.C.; Tsai, S.J. Prostaglandin E2: The master of endometriosis? Exp. Biol. Med. 2010, 235, 668–677. [Google Scholar] [CrossRef]
- Bocci, V. Interleukins: Clinical Pharmacokinetics and Practical Implications. Clin. Pharmacokinet. 1991, 21, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.L.; D’Alexandri, F.L.; Pugliesi, G.; Van Hoeck, V.; Mesquita, F.S.; Membrive, C.M.B.; Negrão, J.A.; Wheelock, C.E.; Binelli, M. Peri-ovulatory endocrine regulation of the prostanoid pathways in the bovine uterus at early dioestrus. Reprod. Fertil. Dev. 2017, 29, 544–556. [Google Scholar] [CrossRef]
- Chabot, V.; Lambert, R.D.; Laforest, J.-P.; St-Jacques, S.; Matte, J.J.; Guay, F.; Palin, M.-F.; Lessard, M. Effect of oestrous cycle and early pregnancy on uterine production and expression of immune regulatory factors in gilts. Anim. Reprod. Sci. 2004, 81, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Samokhodskaya, L.; Mladova, E.; Panina, O. Mucosal biomarkers for endometrial receptivity: A promising yet underexplored aspect of reproductive medicine. Syst. Biol. Reprod. Med. 2021, 68, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Ujioka, T.; Matsukawa, A.; Tanaka, N.; Matsuura, K.; Yoshinaga, M.; Okamura, H. Analysis of the cytokine interaction among interleukin-1β, interleukin- 8, and interleukin-1 receptor antagonist in the rabbit ovulatory process. Fertil. Steril. 1998, 70, 759–765. [Google Scholar] [CrossRef]
- Katila, T.; Ferreira-Dias, G. Evolution of the Concepts of Endometrosis, Post Breeding Endometritis, and Susceptibility of Mares. Animals 2022, 12, 779. [Google Scholar] [CrossRef]
- Liu, Y.; Yao, C.J.; Tao, F.B.; Luo, C.M.; Cao, Y.; Su-Juan, Z.; Chen, Y.Y.; Zhu, X.X.; Cao, J.Y. Association between maternal tumor necrosis factor-α G308A polymorphism and interferon-γ A874T polymorphism and risk of preterm birth: A meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 190, 11–19. [Google Scholar] [CrossRef]
- Kacerovsky, M.; Musilova, I.; Jacobsson, B.; Drahosova, M.; Hornychova, H.; Janku, P.; Prochazka, M.; Simetka, O.; Andrys, C. Cervical fluid IL-6 and IL-8 levels in pregnancies complicated by preterm prelabor rupture of membranes. J. Matern. Neonatal Med. 2014, 28, 134–140. [Google Scholar] [CrossRef]
- Llarena, N.C.; Richards, E.G.; Priyadarshini, A.; Fletcher, D.; Bonfield, T.; Flyckt, R.L. Characterizing the endometrial fluid cytokine profile in women with endometriosis. J. Assist. Reprod. Genet. 2020, 37, 2999. [Google Scholar] [CrossRef] [PubMed]
- Cope, D.I.; Monsivais, D. Progesterone Receptor Signaling in the Uterus Is Essential for Pregnancy Success. Cells 2022, 11, 1474. [Google Scholar] [CrossRef] [PubMed]
- Chi, R.P.A.; Wang, T.; Adams, N.; Wu, S.P.; Young, S.L.; Spencer, T.E.; DeMayo, F. Human Endometrial Transcriptome and Progesterone Receptor Cistrome Reveal Important Pathways and Epithelial Regulators. J. Clin. Endocrinol. Metab. 2020, 105, e1419. [Google Scholar] [CrossRef] [PubMed]
- King, A.E.; Critchley, H.O. Oestrogen and progesterone regulation of inflammatory processes in the human endometrium. J. Steroid Biochem. Mol. Biol. 2010, 120, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Gründker, C.; Emons, G. The Role of Gonadotropin-Releasing Hormone in Cancer Cell Proliferation and Metastasis. Front. Endocrinol. 2017, 8, 187. [Google Scholar] [CrossRef]
- Maggi, R.; Cariboni, A.M.; Marelli, M.M.; Moretti, R.M.; Andrè, V.; Marzagalli, M.; Limonta, P. GnRH and GnRH receptors in the pathophysiology of the human female reproductive system. Hum. Reprod. 2016, 22, 358–381. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Harada, T.; Horie, S.; Iba, Y.; Taniguchi, F.; Yoshida, S.; Iwabe, T.; Terakawa, N. Tumor Necrosis Factor-α-Induced Interleukin-8 (IL-8) Expression in Endometriotic Stromal Cells, Probably through Nuclear Factor-κB Activation: Gonadotropin-Releasing Hormone Agonist Treatment Reduced IL-8 Expression. J. Clin. Endocrinol. Metab. 2003, 88, 730–735. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hidalgo, A.I.; Ulloa-Leal, C.; Gajardo, G.; López, G.; Carretta, D.; Burgos, R.A.; Ratto, M. Ovulation Induced by Intrauterine Seminal Plasma Increases Total Protein, PGE2, IL-8, and IL-1β in Uterine Fluid of Llamas (Lama glama). Animals 2023, 13, 554. https://doi.org/10.3390/ani13040554
Hidalgo AI, Ulloa-Leal C, Gajardo G, López G, Carretta D, Burgos RA, Ratto M. Ovulation Induced by Intrauterine Seminal Plasma Increases Total Protein, PGE2, IL-8, and IL-1β in Uterine Fluid of Llamas (Lama glama). Animals. 2023; 13(4):554. https://doi.org/10.3390/ani13040554
Chicago/Turabian StyleHidalgo, Alejandra Isabel, Cesar Ulloa-Leal, Gonzalo Gajardo, Gerardo López, Daniella Carretta, Rafael Agustín Burgos, and Marcelo Ratto. 2023. "Ovulation Induced by Intrauterine Seminal Plasma Increases Total Protein, PGE2, IL-8, and IL-1β in Uterine Fluid of Llamas (Lama glama)" Animals 13, no. 4: 554. https://doi.org/10.3390/ani13040554