

Two Are Better Than One: Integrating Spatial Geometry with a Conspicuous Landmark in Zebrafish Reorientation Behavior

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
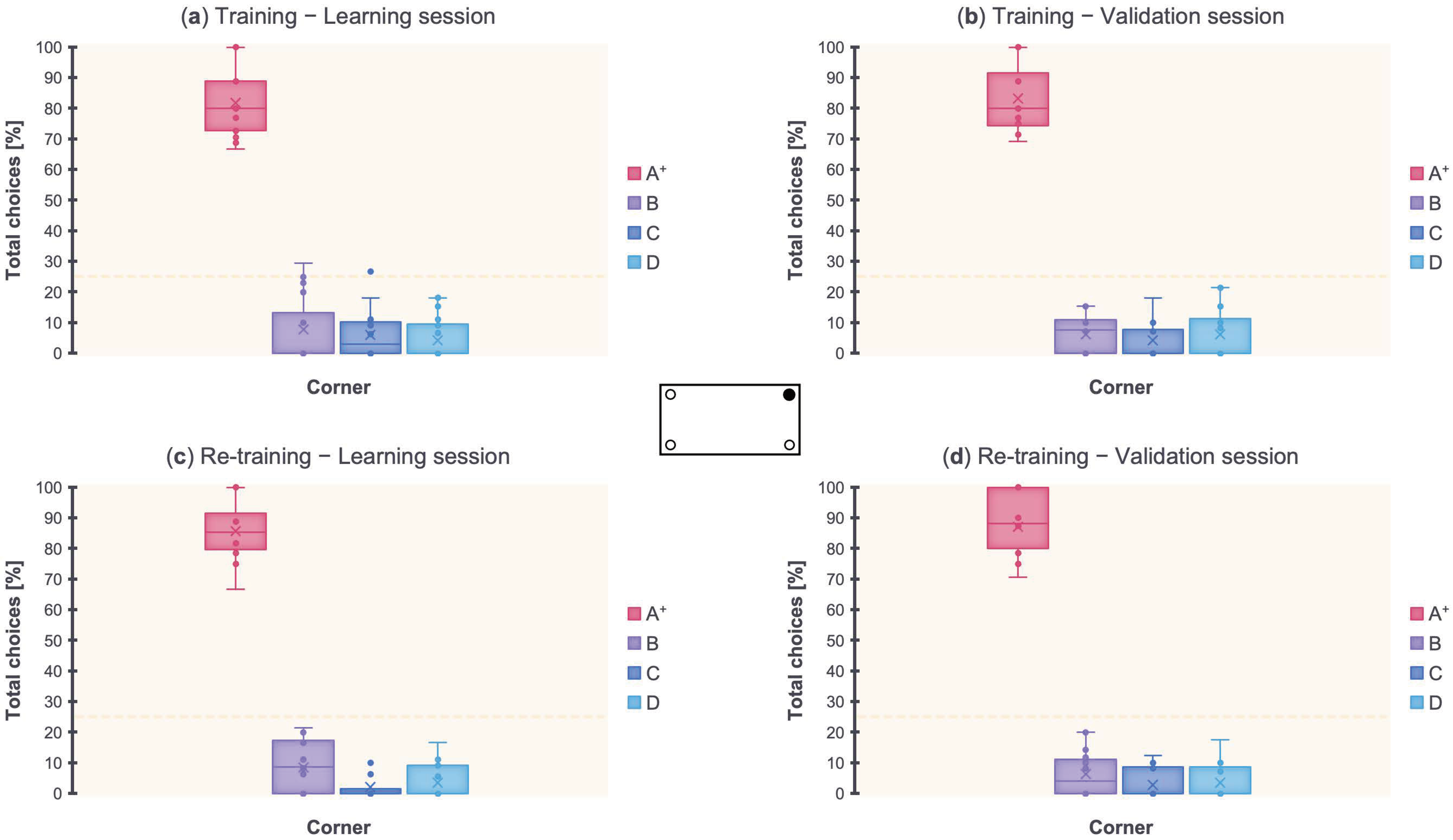
3.1. Training and Re-Training
3.2. Test 1—Geometric Test, Test 2—Affine Transformation
3.3. Movement Patterns: Strategy and Direction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, K.; Gallistel, C.R. Testing the Geometric Power of an Animal’s Spatial Representation. In Animal Cognition; Roitblat, H.L., Terrace, H.S., Bever, T.G., Eds.; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 1984; pp. 409–423. [Google Scholar]
- Cheng, K. A Purely Geometric Module in the Rat’s Spatial Representation. Cognition 1986, 23, 149–178. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Zanforlin, M.; Pasti, G. Geometric modules in animals’ spatial representations: A test with chicks (Gallus gallus domesticus). J. Comp. Psychol. 1990, 104, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.M.; Spetch, M.L.; Heth, C.D. Pigeons’ (Columba livia) encoding of geometric and featural properties of a spatial environment. J. Comp. Psychol. 1998, 112, 259–269. [Google Scholar] [CrossRef]
- Gouteux, S.; Thinus-Blanc, C.; Vauclair, J. Rhesus monkeys use geometric and nongeometric information during a reorientation task. J. Exp. Psychol. General 2001, 130, 505–519. [Google Scholar] [CrossRef]
- Hermer, L.; Spelke, E.S. A geometric process for spatial reorientation in young children. Nature 1994, 370, 57–59. [Google Scholar] [CrossRef]
- Hermer, L.; Spelke, E.S. Modularity and development: The case of spatial reorientation. Cognition 1996, 61, 195–232. [Google Scholar] [CrossRef]
- Learmonth, A.E.; Nadel, L.; Newcombe, N.S. Children’s use of landmarks: Implications for modularity theory. Psychol. Sci. 2002, 13, 337–341. [Google Scholar] [CrossRef]
- Learmonth, A.E.; Newcombe, N.S.; Sheridan, N.; Jones, M. Why size counts: Children’s spatial reorientation in large and small enclosures. Dev. Sci. 2008, 11, 414–426. [Google Scholar] [CrossRef]
- Lee, S.A.; Shusterman, A.; Spelke, E.S. Reorientation and landmark-guided search by young children: Evidence for two systems. Psychol. Sci. 2006, 17, 577–582. [Google Scholar] [CrossRef]
- Twyman, A.; Friedman, A.; Spetch, M.L. Penetrating the geometric module: Catalyzing children’s use of landmarks. Dev. Psychol. 2007, 43, 1523–1530. [Google Scholar] [CrossRef] [Green Version]
- Shusterman, A.; Lee, S.A.; Spelke, E.S. Cognitive effects of language on human navigation. Cognition 2011, 120, 186–201. [Google Scholar] [CrossRef]
- Wang, R.F.; Hermer, L.; Spelke, E.S. Mechanisms of reorientation and object localization by children: A comparison with rats. Behav. Neurosci. 1999, 113, 475–485. [Google Scholar] [CrossRef]
- Baratti, G.; Potrich, D.; Lee, S.A.; Morandi-Raikova, A.; Sovrano, V.A. The Geometric World of Fishes: A Synthesis on Spatial Reorientation in Teleosts. Animals 2022, 12, 881. [Google Scholar] [CrossRef] [PubMed]
- Baratti, G.; Potrich, D.; Sovrano, V.A. The Environmental Geometry in Spatial Learning by Zebrafish (Danio rerio). Zebrafish 2020, 17, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Baratti, G.; Rizzo, A.; Miletto Petrazzini, M.E.; Sovrano, V.A. Learning by Doing: The Use of Distance, Corners and Length in Rewarded Geometric Tasks by Zebrafish (Danio rerio). Animals 2021, 11, 2001. [Google Scholar] [CrossRef]
- Haight, J.L.; Schroeder, J.A. Spatial cognition in Zebrafish. In Zebrafish Models in Neurobehavioral Research; Kalueff, A.V., Cachat, J., Eds.; Humana Press: Totowa, NJ, USA, 2011; pp. 235–248. [Google Scholar] [CrossRef]
- Yashina, K.; Tejero-Cantero, Á.; Herz, A.; Baier, H. Zebrafish exploit visual cues and geometric relationships to form a spatial memory. IScience 2019, 19, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Vallortigara, G.; Flore, M.; Spelke, E.S.; Sovrano, V.A. Navigation by environmental geometry: The use of zebrafish as a model. J. Exp. Biol. 2013, 216, 3693–3699. [Google Scholar] [CrossRef]
- Lee, S.A.; Vallortigara, G.; Ruga, V.; Sovrano, V.A. Independent effects of geometry and landmark in a spontaneous reorientation task: A study of two species of fish. Anim. Cogn. 2012, 15, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Spence, R.; Magurran, A.E.; Smith, C. Spatial cognition in zebrafish: The role of strain and rearing environment. Anim. Cogn. 2011, 14, 607–612. [Google Scholar] [CrossRef]
- Sovrano, V.A.; Baratti, G.; Potrich, D.; Bertolucci, C. The geometry as an eyed fish feels it in spontaneous and rewarded spatial reorientation tasks. Sci. Rep. 2020, 10, 8020. [Google Scholar] [CrossRef]
- Lee, S.A.; Ferrari, A.; Vallortigara, G.; Sovrano, V.A. Boundary primacy in spatial mapping: Evidence from zebrafish (Danio rerio). Behav. Process. 2015, 119, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Gatto, E.; Santacà, M.; Verza, I.; Dadda, M.; Bisazza, A. Automated Operant Conditioning Devices for Fish. Do They Work? Animals 2021, 11, 1397. [Google Scholar] [CrossRef] [PubMed]
- Miletto Petrazzini, M.E.; Pecunioso, A.; Dadda, M.; Agrillo, C. The Impact of brain lateralization and anxiety-like behaviour in an extensive operant conditioning task in zebrafish (Danio rerio). Symmetry 2019, 11, 1395. [Google Scholar] [CrossRef]
- Santacà, M.; Dadda, M.; Miletto Petrazzini, M.E.; Bisazza, A. Stimulus characteristics, learning bias and visual discrimination in zebrafish (Danio rerio). Behav. Process. 2021, 192, 104499. [Google Scholar] [CrossRef] [PubMed]
- Sovrano, V.A.; Vicidomini, S.; Potrich, D.; Miletto Petrazzini, M.E.; Baratti, G.; Rosa-Salva, O. Visual discrimination and amodal completion in zebrafish. PloS ONE 2022, 17, e0264127. [Google Scholar] [CrossRef] [PubMed]
- Sovrano, V.A.; Bisazza, A.; Vallortigara, G. Modularity and Spatial Reorientation in a Simple Mind: Encoding of Geometric and Nongeometric Properties of a Spatial Environment by Fish. Cognition 2002, 85, B51–B59. [Google Scholar] [CrossRef]
- Sovrano, V.A.; Bisazza, A.; Vallortigara, G. Modularity as a Fish (Xenotoca eiseni) Views It: Conjoining Geometric and Nongeometric Information for Spatial Reorientation. J. Exp. Psychol. Anim. Behav. Process. 2003, 29, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Sovrano, V.A.; Bisazza, A.; Vallortigara, G. Animals’ Use of Landmarks and Metric Information to Reorient: Effects of the Size of the Experimental Space. Cognition 2005, 97, 121–133. [Google Scholar] [CrossRef]
- Sovrano, V.A.; Dadda, M.; Bisazza, A. Lateralized Fish Perform Better than Nonlateralized Fish in Spatial Reorientation Tasks. Behav. Brain Res. 2005, 163, 122–127. [Google Scholar] [CrossRef]
- Sovrano, V.A.; Bisazza, A.; Vallortigara, G. How Fish Do Geometry in Large and in Small Spaces. Anim. Cogn. 2007, 10, 47–54. [Google Scholar] [CrossRef]
- Dijkgraaf, S. The Functioning and Significance of the Lateral-Line Organs. Biol. Rev. Camb. Philos. Soc. 1963, 38, 51–105. [Google Scholar] [CrossRef] [PubMed]
- Dijkgraaf, S. The Supposed Use of the Lateral Line as an Organ of Hearing in Fish. Experientia 1964, 20, 586–587. [Google Scholar] [CrossRef]
- Dijkgraaf, S. A Short Personal Review of the History of Lateral Line Research. In The Mechanosensory Lateral Line; Coombs, S., Görner, P., Münz, H., Eds.; Springer: New York, NY, USA, 1989; pp. 7–14. [Google Scholar] [CrossRef]
- Sovrano, V.A.; Potrich, D.; Foà, A.; Bertolucci, C. Extra-Visual Systems in the Spatial Reorientation of Cavefish. Sci. Rep. 2018, 8, 17698. [Google Scholar] [CrossRef]
- Kasumyan, A.O. The lateral line in fish: Structure, function, and role in behavior. J. Ichthyol. 2003, 43, S175–S231. [Google Scholar]
- Sovrano, V.A.; Baratti, G.; Lee, S.A. The role of learning and environmental geometry in landmark-based spatial reorientation of fish (Xenotoca eiseni). PloS ONE 2020, 15, e0229608. [Google Scholar] [CrossRef] [PubMed]
- Caro, T.M.; Roper, R.; Young, M.; Dank, G.R. Inter-observer reliability. Behaviour 1979, 69, 303–315. [Google Scholar] [CrossRef]
- Cheng, K.; Huttenlocher, J.; Newcombe, N.S. 25 years of research on the use of geometry in spatial reorientation: A current theoretical perspective. Psychon. Bull. Rev. 2013, 20, 1033–1054. [Google Scholar] [CrossRef]
- Vallortigara, G. Comparative cognition of number and space: The case of geometry and of the mental number line. Philos. Trans. Royal Soc. B Biol. Sci. 2018, 373, 20170120. [Google Scholar] [CrossRef]
- Tommasi, L.; Chiandetti, C.; Pecchia, T.; Sovrano, V.A.; Vallortigara, G. From natural geometry to spatial cognition. Neurosci. Biobehav. Rev. 2012, 36, 799–824. [Google Scholar] [CrossRef]
- Vallortigara, G. Animals as natural geometers. In Cognitive Biology; Tommasi, L., Peterson, M.A., Nadel, L., Eds.; MIT Press: Cambridge, MA, USA, 2009; pp. 83–104. [Google Scholar] [CrossRef]
- Vargas, J.P.; López, J.C.; Salas, C.; Thinus-Blanc, C. Encoding of geometric and featural spatial information by goldfish (Carassius auratus). J. Comp. Psychol. 2004, 118, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Dawson, M.R.W.; Kelly, D.M.; Spetch, M.L.; Dupuis, B. Using perceptrons to explore the reorientation task. Cognition 2010, 114, 207–226. [Google Scholar] [CrossRef]
- Miglino, O.; Lund, H.H. Do rats need Euclidean cognitive maps of the environmental shape? Cogn. Process. 2001, 3, 22–33. [Google Scholar]
- Miglino, O.; Ponticorvo, M.; Bartolomeo, P. Place cognition and active perception: A study with evolved robots. Connect. Sci. 2009, 21, 3–14. [Google Scholar] [CrossRef]
- Miglino, O.; Ponticorvo, M. Place Cognition as an Example of Situated Cognition: A Study with Evolved Agents. Cogn. Process. 2009, 10, S250–S252. [Google Scholar] [CrossRef]
- Ponticorvo, M.; Miglino, O. Encoding geometric and non-geometric information: A study with evolved agents. Anim. Cogn. 2010, 13, 157–174. [Google Scholar] [CrossRef]
- Baratti, G.; Boffelli, S.; Potrich, D.; Sovrano, V.A. Spatial Learning by Using Non-Visual Geometry and a Visual 3D Landmark in Zebrafish (Danio rerio). Animals 2023, 13, 440. [Google Scholar] [CrossRef]
- Barnett, S.A. The Rat: A Study in Behavior; Metheun: London, UK, 1963. [Google Scholar]
- Treit, D.; Fundytus, M. Thigmotaxis as a test for anxiolytic activity in rats. Pharmacol. Biochem. Behav. 1988, 31, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Simon, P.; Dupuis, R.; Costentin, J. Thigmotaxis as an index of anxiety in mice. Influence of dopaminergic transmissions. Behav. Brain Res. 1994, 61, 59–64. [Google Scholar] [CrossRef]
- Hill, E.W.; Rieser, J.J.; Hill, M.M.; Hill, M.; Halpin, J.; Halpin, R. How persons with visual impairments explore novel spaces: Strategies of good and poor performers. JVIB 1993, 87, 295–301. [Google Scholar] [CrossRef]
- Kallai, J.; Makany, T.; Csatho, A.; Karadi, K.; Horvath, D.; Kovacs-Labadi, B.; Jacobs, J.W. Cognitive and affective aspects of thigmotaxis strategy in humans. Behav. Neurosci. 2007, 121, 21–30. [Google Scholar] [CrossRef]
- Kallai, J.; Makany, T.; Karadi, K.; Jacobs, W.J. Spatial orientation strategies in Morris-type virtual water task for humans. Behav. Brain Res. 2005, 159, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, H.; Hassan, E.S.; Von Campenhausen, C. Sensory performance of blind Mexican cave fish after destruction of the canal neuromasts. Naturwissenschaften 1990, 77, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Breder, C.M.; Gresser, E.B. Correlation between structural eye defects and behavior in the Mexican blind characin. Zoologica 1941, 26, 123–131. [Google Scholar] [CrossRef]
- Gertychowa, R. Studies on the ethology and space orientation of the blind cave fish Anoptichthys jordani Hubbs et Innes 1936 (Characidae). Folia Biol. 1970, 18, 9–69. [Google Scholar]
- Sharma, S.; Coombs, S.; Patton, P.; Burt De Perera, T. The function of wall-following behaviors in the Mexican blind cavefish and a sighted relative, the Mexican tetra (Astyanax). J. Comp. Physiol. A 2009, 195, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Teyke, T. Collision with and avoidance of obstacles by blind cave fish Anoptichthys jordani (Characidae). J. Comp. Physiol. A 1985, 157, 837–843. [Google Scholar] [CrossRef]
- Teyke, T. Learning and remembering the environment in the blind cave fish Anoptichthys jordani. J. Comp. Physiol. A 1989, 164, 655–662. [Google Scholar] [CrossRef]
- Basil, J.; Sandeman, D. Crayfish (Cherax destructor) use tactile cues to detect and learn topographical changes in their environment. Ethology 2000, 106, 247–259. [Google Scholar] [CrossRef]
- Creed, R.P.; Miller, J.R. Interpreting animal wall-following behavior. Experientia 1990, 46, 758–761. [Google Scholar] [CrossRef]
- Jeanson, R.; Blanco, S.; Fournier, R.; Deneubourg, J.L.; Fourcassié, V.; Theraulaz, G. A model of animal movements in a bounded space. J. Theor. Biol. 2003, 225, 443–451. [Google Scholar] [CrossRef]
- Besson, M.; Martin, J.R. Centrophobism/thigmotaxis, a new role for the mushroom bodies in Drosophila. J. Neurobiol. 2004, 62, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.R. A portrait of locomotor behaviour in Drosophila determined by a video-tracking paradigm. Behav. Process. 2004, 67, 207–219. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, J.; Nadel, L. The Hippocampus As a Cognitive Map; Clarendon Press: Oxford, UK, 1978. [Google Scholar]
- Oman, C.M.; Shebilske, W.L.; Richards, J.T.; Tubré, T.C.; Beall, A.C.; Natapoff, A. Three dimensional spatial memory and learning in real and virtual environments. Spat. Cogn. Comput. 2000, 2, 355–372. [Google Scholar] [CrossRef]
- Choleris, E.; Thomas, A.W.; Kavaliers, M.; Prato, F.S. A detailed ethological analysis of the mouse open field test: Effects of diazepam, chlordiazepoxide and an extremely low frequency pulsed magnetic field. Neurosci. Biobehav. Rev. 2001, 25, 235–260. [Google Scholar] [CrossRef] [PubMed]
- Ohl, F.; Holsboer, F.; Landgraf, R. The modified hole board as a differential screen for behavior in rodents. Behav. Res. Methods Instrum. Comput. 2001, 33, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Pellow, S.; File, S.E. Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: A novel test of anxiety in the rat. Pharmacol. Biochem. Behav. 1986, 24, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Grunwald, D.J.; Eisen, J.S. Headwaters of the zebrafish—Emergence of a new model vertebrate. Nat. Rev. Gen. 2002, 3, 717–724. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Training | Re-Training | |||
|---|---|---|---|---|
| Fish | % Learning | % Validation | % Learning | % Validation |
| 1 | 70 | 87.5 | 66.67 | 87.5 |
| 2 | 87.5 | 66.67 1 | 87.50 | 66.67 |
| 3 | 87.5 | 75 | 66.67 | 75 |
| 4 | 75 | 70 | 75 | 77.78 |
| 5 | 75 | 77.78 | 87.5 | 75 |
| 6 | 75 | 70 | 80 | 87.5 |
| 7 | 70 | 100 | 87.5 | 75 |
| 8 | 100 | 100 | 100 | 100 |
| 9 | 87.5 | 100 | 100 | 100 |
| 10 | 87.5 | 75 | 100 | 100 |
| 11 | 100 | 70 | 87.5 | 83.33 |
| 12 | 70 | 75 | 87.5 | 87.5 |
| 13 | 75 | 70 | 70 | 100 |
| 14 | 75 | 75 | 75 | 75 |
| 15 | 87.5 | 87.5 | 100 | 100 |
| 16 | 87.5 | 100 | 87.5 | 87.5 |
| 17 | 87.5 | 87.5 | 75 | 75 |
| 18 | 75 | 75 | 75 | 75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baratti, G.; Sovrano, V.A. Two Are Better Than One: Integrating Spatial Geometry with a Conspicuous Landmark in Zebrafish Reorientation Behavior. Animals 2023, 13, 537. https://doi.org/10.3390/ani13030537
Baratti G, Sovrano VA. Two Are Better Than One: Integrating Spatial Geometry with a Conspicuous Landmark in Zebrafish Reorientation Behavior. Animals. 2023; 13(3):537. https://doi.org/10.3390/ani13030537
Chicago/Turabian StyleBaratti, Greta, and Valeria Anna Sovrano. 2023. "Two Are Better Than One: Integrating Spatial Geometry with a Conspicuous Landmark in Zebrafish Reorientation Behavior" Animals 13, no. 3: 537. https://doi.org/10.3390/ani13030537