
Next-Generation Sequencing of the Complete Huaibei Grey Donkey Mitogenome and Mitogenomic Phylogeny of the Equidae Family
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Specimen Collection and DNA Isolation
2.3. Amplification of Part of the mtDNA D-Loop and Sequencing
2.4. Sequence Assembly Annotation and Analysis
2.5. Phylogenetic Analysis
3. Results
3.1. Structure and Organization of the Complete Mitochondrial Genome
3.2. Codon Usage and Protein Coding Genes
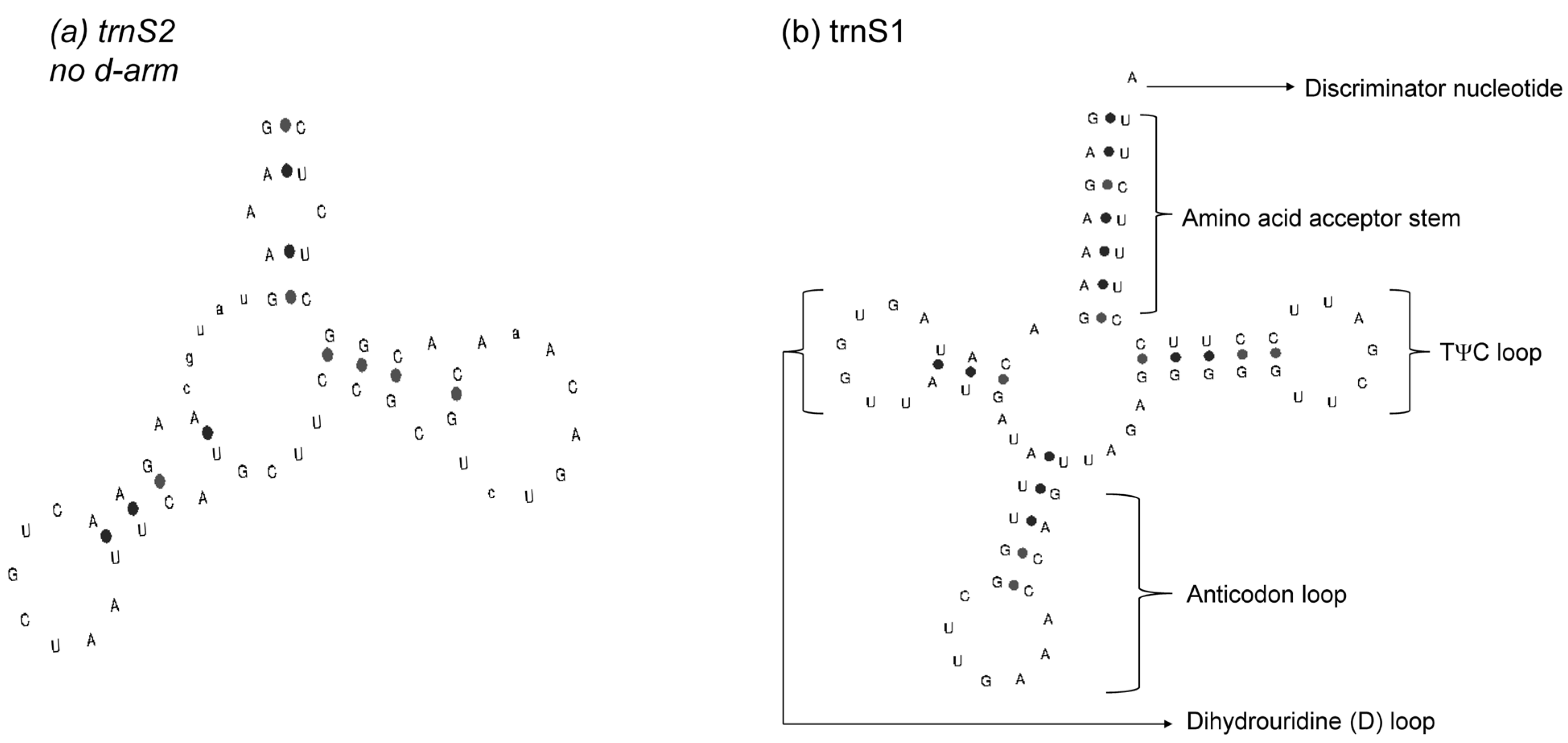
3.3. Transfer and Ribosomal RNA Genes
3.4. Genetic Variation and Genetic Diversity of Complete D-Loop Region
3.5. Maternal Origin of Huaibei Grey Donkey
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F.; et al. Sequence and organization of the human mitochondrial genome. Nature 1981, 290, 457–465. [Google Scholar] [CrossRef]
- Achilli, A.; Olivieri, A.; Soares, P.; Lancioni, H.; Hooshiar Kashani, B.; Perego, U.A.; Nergadze, S.G.; Carossa, V.; Santagostino, M.; Capomaccio, S.; et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 2449–2454. [Google Scholar] [CrossRef]
- Bruford, M.W.; Bradley, D.G.; Luikart, G. DNA markers reveal the complexity of livestock domestication. Nat. Rev. Genet. 2003, 4, 900–910. [Google Scholar] [CrossRef] [PubMed]
- Saccone, C.; Gissi, C.; Lanave, C.; Larizza, A.; Pesole, G.; Reyes, A. Evolution of the mitochondrial genetic system: An overview. Gene 2000, 261, 153–159. [Google Scholar] [CrossRef]
- Wolf, C.; Rentsch, J.; Hubner, P. PCR-RFLP analysis of mitochondrial DNA: A reliable method for species identification. J. Agric. Food Chem. 1999, 47, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Nobushige, I.; Tsendsuren, O.; Suguru, M.; Harutaka, M.; Naruya, S. Mitochondrial DNA sequences of various species of the genus Equus with special reference to the phylogenetic relationship between Przewalskii’s wild horse and domestic horse. J. Mol. Evol. 1995, 41, 180–188. [Google Scholar]
- Gissi, C.; Reyes, A.; Pesole, G.; Saccone, C. Lineage-specific evolutionary rate in mammalian mtDNA. Mol. Biol. Evol. 2000, 17, 1022–1031. [Google Scholar] [CrossRef]
- Beja-Pereira, A.; England, P.R.; Ferrand, N.; Jordan, S.; Bakhiet, A.O.; Abdalla, M.A.; Mashkour, M.; Jordana, J.; Taberlet, P.; Luikart, G. African origins of the domestic donkey. Science 2004, 304, 1781. [Google Scholar] [CrossRef]
- Kimura, B.; Marshall, F.B.; Chen, S.; Rosenbom, S.; Moehlman, P.D.; Tuross, N.; Sabin, R.C.; Peters, J.; Barich, B.; Yohannes, H.; et al. Ancient DNA from Nubian and Somali wild ass provides insights into donkey ancestry and domestication. Proc. R. Soc. B Biol. Sci. 2011, 278, 50–57. [Google Scholar] [CrossRef]
- Chen, S.Y.; Zhou, F.; Xiao, H.; Sha, T.; Wu, S.F.; Zhang, Y.P. Mitochondrial DNA diversity and population structure of four Chinese donkey breeds. Anim. Genet. 2006, 37, 427–429. [Google Scholar] [CrossRef]
- Han, L.; Zhu, S.; Ning, C.; Cai, D.; Wang, K.; Chen, Q.; Hu, S.; Yang, J.; Shao, J.; Zhu, H.; et al. Ancient DNA provides new insight into the maternal lineages and domestication of Chinese donkeys. BMC Evol. Biol. 2014, 14, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, C.Z.; Ge, Q.L.; Zhang, H.C.; Liu, R.Y.; Zhang, W.; JIang, Y.Q.; Dang, R.H.; Zheng, H.L.; Hou, W.T.; Chen, H. African maternal origin and genetic diversity of chinese domestic donkeys. Asian-Australas. J. Anim. Sci. 2007, 20, 645–652. [Google Scholar] [CrossRef]
- Gan, Q.F.; Lin, S.X.; Liang, X.W.; Meng, H.L.; Qiao, H.; Du, Y.F.; Ke, F.R.; Zheng, S.K. African maternal origin and genetic diversity of five chinese domestic donkeys breeds. J. Anim. Vet. Adv. 2011, 10, 3090–3094. [Google Scholar]
- Perez-Pardal, L.; Grizelj, J.; Traore, A.; Cubric-Curik, V.; Arsenos, G.; Dovenski, T.; Markovic, B.; Fernandez, I.; Cuervo, M.; Alvarez, I.; et al. Lack of mitochondrial DNA structure in Balkan donkey is consistent with a quick spread of the species after domestication. Anim. Genet. 2014, 45, 144–147. [Google Scholar] [CrossRef]
- Ozkan Unal, E.; Ozdil, F.; Kaplan, S.; Gurcan, E.K.; Genc, S.; Arat, S.; Soysal, M.I. Phylogenetic relationships of Turkish indigenous donkey populations determined by mitochondrial DNA D-loop region. Animals 2020, 10, 1970. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Ning, T.; Adeola, A.C.; Li, J.; Esmailizadeh, A.; Lichoti, J.K.; Agwanda, B.R.; Isakova, J.; Aldashev, A.A.; Wu, S.F.; et al. Potential dual expansion of domesticated donkeys revealed by worldwide analysis on mitochondrial sequences. Zool. Res. 2020, 41, 51–60. [Google Scholar] [PubMed]
- Xu, X.; Gullberg, A.; Arnason, U. The complete mitochondrial DNA (mtDNA) of the donkey and mtDNA comparisons among four closely related mammalian species-pairs. J. Mol. Evol. 1996, 43, 438–446. [Google Scholar] [CrossRef]
- Ishida, N.; Hasegawa, T.; Takeda, K.; Sakagami, M.; Onishi, A.; Inumaru, S.; Komatsu, M.; Mukoyama, H. Polymorphic sequence in the D-loop region of equine mitochondrial DNA. Anim. Genet. 1994, 25, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA genes in genomic sequences. In Gene Prediction; Methods in Molecular Biology; Kollmar, M., Ed.; Humana: New York, NY, USA, 2019; Volume 1962, pp. 1–14. [Google Scholar]
- Greiner, S.; Lehwark, P.; Bock, R. Organellar Genome DRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Xia, X.; Yu, J.; Zhao, X.; Yao, Y.; Zeng, L.; Ahmed, Z.; Shen, S.; Dang, R.; Lei, C. Genetic diversity and maternal origin of Northeast African and South American donkey populations. Anim. Genet. 2019, 50, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Ivankovic, A.; Kavar, T.; Caput, P.; Mioc, B.; Pavic, V.; Dovc, P. Genetic diversity of three donkey populations in the Croatian coastal region. Anim. Genet. 2002, 33, 169–177. [Google Scholar] [CrossRef]
- Oakenfull, E.A.; Lim, H.N.; Ryder, O.A. A survey of equid mitochondrial DNA: Implications for the evolution, genetic diversity and conservation of Equus. Conserv. Genet. 2000, 1, 341–355. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.P.; Danforth, B.N. How do insect nuclear and mitochondrial gene substitution patterns differ? Insights from Bayesian analyses of combined datasets. Mol. Phylogenet. Evol. 2004, 30, 686–702. [Google Scholar] [CrossRef]
- Gissi, C.; Iannelli, F.; Pesole, G. Evolution of the mitochondrial genome of Metazoa as exemplified by comparison of congeneric species. Heredity 2008, 101, 301–320. [Google Scholar] [CrossRef] [PubMed]
- Sbisa, E.; Tanzariello, F.; Reyes, A.; Pesole, G.; Saccone, C. Mammalian mitochondrial D-loop region structural analysis: Identification of new conserved sequences and their functional and evolutionary implications. Gene 1997, 205, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jiang, Q.; Yang, C.; Wang, X.; Tian, F.; Wang, Y.; Ma, Y.; Ju, Z.; Huang, J.; Zhou, X.; et al. Characterization of complete mitochondrial genome of Dezhou donkey (Equus asinus) and evolutionary analysis. Curr. Genet. 2016, 62, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.L.; Alonso, R.; Aluja, A.S.D. Study of the genetic origin of the Mexican creole donkey (Equus asinus) by means of the analysis of the D-loop region of mitochondrial DNA. Trop. Anim. Health Prod. 2005, 37, 173–188. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Position | Length/bp | Strand | Anticodon | Codon | ||
|---|---|---|---|---|---|---|---|
| From | To | Start | Stop | ||||
| tRNA-Phe | 1 | 71 | 71 | H | GAA | ||
| 12S rRNA | 72 | 1046 | 975 | H | |||
| tRNA-Val | 1046 | 1112 | 67 | H | UAC | ||
| 16S rRNA | 1113 | 2692 | 1580 | H | |||
| tRNA-Leu | 2693 | 2767 | 75 | H | UAA | ||
| ND1 | 2770 | 3726 | 957 | H | ATG | TAA | |
| tRNA-Ile | 3726 | 3794 | 69 | H | GAU | ||
| tRNA-Gln | 3792 | 3864 | 73 | L | UUG | ||
| tRNA-Met | 3867 | 3935 | 69 | H | CAU | ||
| ND2 | 3936 | 4976 | 1041 | H | ATA | TAG | |
| tRNA-Trp | 4975 | 5043 | 69 | H | UCA | ||
| tRNA-Ala | 5049 | 5117 | 69 | L | UGC | ||
| tRNA-Asn | 5119 | 5191 | 73 | L | GUU | ||
| tRNA-Cys | 5224 | 5289 | 66 | L | GCA | ||
| tRNA-Tyr | 5290 | 5356 | 67 | L | GUA | ||
| COX1 | 5358 | 6902 | 1545 | H | ATG | TAA | |
| tRNA-Ser | 6900 | 6968 | 69 | L | UGA | ||
| tRNA-Asp | 6977 | 7043 | 67 | H | GUC | ||
| COX2 | 7045 | 7728 | 684 | H | ATG | TAA | |
| tRNA-Lys | 7732 | 7800 | 69 | H | UUU | ||
| ATP8 | 7802 | 8005 | 204 | H | ATG | TAA | |
| ATP6 | 7963 | 8643 | 681 | H | ATG | TAA | |
| COX3 | 8643 | 9427 | 785 | H | ATG | TAG | |
| tRNA-Gly | 9427 | 9496 | 70 | H | UCC | ||
| ND3 | 9497 | 9843 | 347 | H | ATA | TAG | |
| tRNA-Arg | 9844 | 9912 | 69 | H | UCG | ||
| ND4L | 9914 | 10,210 | 297 | H | ATG | TAA | |
| ND4 | 10,204 | 11,581 | 1378 | H | ATG | AGA | |
| tRNA-His | 11,582 | 11,650 | 69 | H | GUG | ||
| tRNA-Ser | 11,651 | 11,710 | 60 | H | UGA | ||
| tRNA-Leu | 11,712 | 11,781 | 70 | H | UAG | ||
| ND5 | 11,773 | 13,602 | 1830 | H | ATA | TAA | |
| ND6 | 13,586 | 14,110 | 525 | L | ATG | TAA | |
| tRNA-Glu | 14,114 | 14,182 | 69 | L | UUC | ||
| CYTB | 14,187 | 15,326 | 1140 | H | ATG | AGA | |
| tRNA-Thr | 15,327 | 15,398 | 72 | H | UGU | ||
| tRNA-Pro | 15,400 | 15,465 | 66 | L | UGG | ||
| D-loop | 15,466 | 16,681 | 1216 | H | |||
| Gene/Region | T(%) | C(%) | A(%) | G(%) | A+T(%) | AT-Skew | GC-Skew |
|---|---|---|---|---|---|---|---|
| tRNA | 27.18 | 22.69 | 34.50 | 15.63 | 61.68 | 0.1187 | −0.1842 |
| rRNA | 21.44 | 24.62 | 36.41 | 17.54 | 57.85 | 0.2588 | −0.1679 |
| ND1 | 27.38 | 30.20 | 31.14 | 11.29 | 58.52 | 0.0643 | −0.4558 |
| ND2 | 24.78 | 31.32 | 34.77 | 9.13 | 59.55 | 0.1678 | −0.5486 |
| ND3 | 26.86 | 28.86 | 31.14 | 13.14 | 58.00 | 0.0738 | −0.3743 |
| ND4 | 25.91 | 31.28 | 31.71 | 11.10 | 57.62 | 0.1007 | −0.4762 |
| ND5 | 25.63 | 32.08 | 31.69 | 10.60 | 57.32 | 0.1057 | −0.5033 |
| ND6 | 19.62 | 31.62 | 41.14 | 7.62 | 60.76 | 0.3542 | −0.6116 |
| ND4L | 28.96 | 28.62 | 29.29 | 13.13 | 58.25 | 0.0057 | −0.3710 |
| COX1 | 28.61 | 27.38 | 28.03 | 15.99 | 56.64 | −0.0102 | −0.2626 |
| COX2 | 25.58 | 28.07 | 32.46 | 13.89 | 58.04 | 0.1185 | −0.3379 |
| COX3 | 26.62 | 30.70 | 27.13 | 15.54 | 53.75 | 0.0095 | −0.3279 |
| ATP6 | 27.61 | 29.66 | 31.72 | 11.01 | 59.33 | 0.0693 | −0.4586 |
| ATP8 | 27.94 | 25.00 | 37.75 | 9.31 | 65.69 | 0.1493 | −0.4573 |
| CYTB | 26.32 | 32.46 | 28.60 | 12.63 | 54.92 | 0.0415 | −0.4398 |
| D-loop | 21.65 | 34.90 | 30.37 | 13.09 | 52.02 | 0.1676 | −0.4545 |
| Overall | 25.59 | 28.88 | 32.31 | 13.22 | 57.90 | 0.1161 | −0.3720 |
| Codon | Count | RSCU | Codon | Count | RSCU | Codon | Count | RSCU | Codon | Count | RSCU |
|---|---|---|---|---|---|---|---|---|---|---|---|
| UUU(F) | 47 | 0.65 | UCU(S) | 65 | 1.01 | UAU(Y) | 73 | 0.94 | UGU(C) | 9 | 0.82 |
| UUC(F) | 98 | 1.35 | UCC(S) | 108 | 1.67 | UAC(Y) | 82 | 1.06 | UGC(C) | 13 | 1.18 |
| UUA(L) | 73 | 1.13 | UCA(S) | 126 | 1.95 | UAA(*) | 63 | 1.06 | UGA(*) | 64 | 1.07 |
| UUG(L) | 17 | 0.26 | UCG(S) | 34 | 0.53 | UAG(*) | 52 | 0.87 | UGG(W) | 9 | 1 |
| CUU(L) | 53 | 0.82 | CCU(P) | 75 | 1.08 | CAU(H) | 72 | 0.98 | CGU(R) | 9 | 0.71 |
| CUC(L) | 75 | 1.16 | CCC(P) | 84 | 1.21 | CAC(H) | 75 | 1.02 | CGC(R) | 14 | 1.11 |
| CUA(L) | 137 | 2.12 | CCA(P) | 110 | 1.58 | CAA(Q) | 76 | 1.27 | CGA(R) | 21 | 1.66 |
| CUG(L) | 32 | 0.5 | CCG(P) | 9 | 0.13 | CAG(Q) | 44 | 0.73 | CGG(R) | 10 | 0.79 |
| AUU(I) | 62 | 0.65 | ACU(T) | 77 | 1.04 | AAU(N) | 62 | 0.89 | AGU(S) | 14 | 0.22 |
| AUC(I) | 125 | 1.3 | ACC(T) | 80 | 1.08 | AAC(N) | 77 | 1.11 | AGC(S) | 40 | 0.62 |
| AUA(I) | 101 | 1.05 | ACA(T) | 110 | 1.49 | AAA(K) | 90 | 1.75 | AGA(R) | 10 | 0.79 |
| AUG(M) | 36 | 1 | ACG(T) | 29 | 0.39 | AAG(K) | 13 | 0.25 | AGG(R) | 12 | 0.95 |
| GUU(V) | 22 | 0.86 | GCU(A) | 46 | 1.07 | GAU(D) | 31 | 0.93 | GGU(G) | 14 | 0.5 |
| GUC(V) | 28 | 1.1 | GCC(A) | 65 | 1.51 | GAC(D) | 36 | 1.07 | GGC(G) | 36 | 1.27 |
| GUA(V) | 38 | 1.49 | GCA(A) | 53 | 1.23 | GAA(E) | 64 | 1.56 | GGA(G) | 52 | 1.84 |
| GUG(V) | 14 | 0.55 | GCG(A) | 8 | 0.19 | GAG(E) | 18 | 0.44 | GGG(G) | 11 | 0.39 |
| Haplotypes | Haplogroup | Localization of Polymorphic Sites | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 66 | 72 | 85 | 151 | 162 | 172 | 174 | 180 | 181 | 202 | 203 | 226 | 227 | 234 | 244 | 280 | 297 | 352 | 383 | 388 | 402 | 403 | 404 | ||
| NC_001788.1 | G | C | T | A | A | A | A | C | A | T | A | G | A | C | A | C | A | T | C | C | C | G | G | |
| H1 | Clade II | A | T | C | G | G | * | * | T | G | C | G | A | * | T | G | T | * | C | T | T | T | A | A |
| H2 | Clade I | * | * | * | * | * | * | * | * | G | * | * | * | * | * | * | * | T | * | * | * | * | A | * |
| H3 | Clade II | A | T | C | G | * | * | * | T | G | * | * | A | G | T | G | T | * | C | T | * | T | A | A |
| H4 | Clade II | A | T | C | G | G | * | * | T | G | * | * | A | * | T | G | T | * | C | T | T | T | A | A |
| H5 | Clade I | * | * | * | * | * | * | * | * | G | * | * | * | * | * | * | * | * | * | * | * | * | A | * |
| H6 | Clade II | A | T | C | G | G | * | * | T | G | * | * | A | G | T | G | T | * | C | T | * | T | A | A |
| H7 | Clade II | A | T | C | G | * | * | * | T | G | * | * | A | * | T | G | T | * | C | T | T | T | A | A |
| H8 | Clade II | A | T | C | G | G | * | G | T | G | * | * | A | G | T | G | T | * | C | T | * | T | A | A |
| H9 | Clade I | * | * | * | * | * | * | * | * | G | * | * | * | * | * | * | * | * | * | * | * | * | * | * |
| H10 | Clade II | A | T | C | G | G | G | * | T | G | * | * | A | * | T | G | T | * | C | T | T | T | A | A |
| H11 | Clade I | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | * | A | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, J.; Chang, L.; Xu, D.; Jia, Y.; Ding, Y.; Cao, C.; Geng, Z.; Jin, S. Next-Generation Sequencing of the Complete Huaibei Grey Donkey Mitogenome and Mitogenomic Phylogeny of the Equidae Family. Animals 2023, 13, 531. https://doi.org/10.3390/ani13030531
Xia J, Chang L, Xu D, Jia Y, Ding Y, Cao C, Geng Z, Jin S. Next-Generation Sequencing of the Complete Huaibei Grey Donkey Mitogenome and Mitogenomic Phylogeny of the Equidae Family. Animals. 2023; 13(3):531. https://doi.org/10.3390/ani13030531
Chicago/Turabian StyleXia, Jingjing, Liang Chang, Dashuang Xu, Yuqing Jia, Yuanfei Ding, Chengcheng Cao, Zhaoyu Geng, and Sihua Jin. 2023. "Next-Generation Sequencing of the Complete Huaibei Grey Donkey Mitogenome and Mitogenomic Phylogeny of the Equidae Family" Animals 13, no. 3: 531. https://doi.org/10.3390/ani13030531