
Effect of High Efficiency Digestion and Utilization of Organic Iron Made by Saccharomyces cerevisiae on Antioxidation and Caecum Microflora in Weaned Piglets

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Animals, Design, Diets and Feeding Management
2.3. Sample Collection
2.4. Determination of Dry Matter and Crude Protein in Feeds and Feces
2.5. Determination of Iron Content in Feed, Feces and Tissues
2.6. Determination of Antioxidant Indexes in Serum, Intestinal Tissue and Liver
2.7. Analysis of 16S rDNA Gene Sequencing
2.8. Statistical Analysis
3. Results
3.1. Digestibility of Iron and Protein
3.2. Serum Iron Index and the Iron Concentration in Organ Tissue
3.3. pH of Intestinal Contents
3.4. Antioxidant Indexes of Serum, Liver and Jejunum
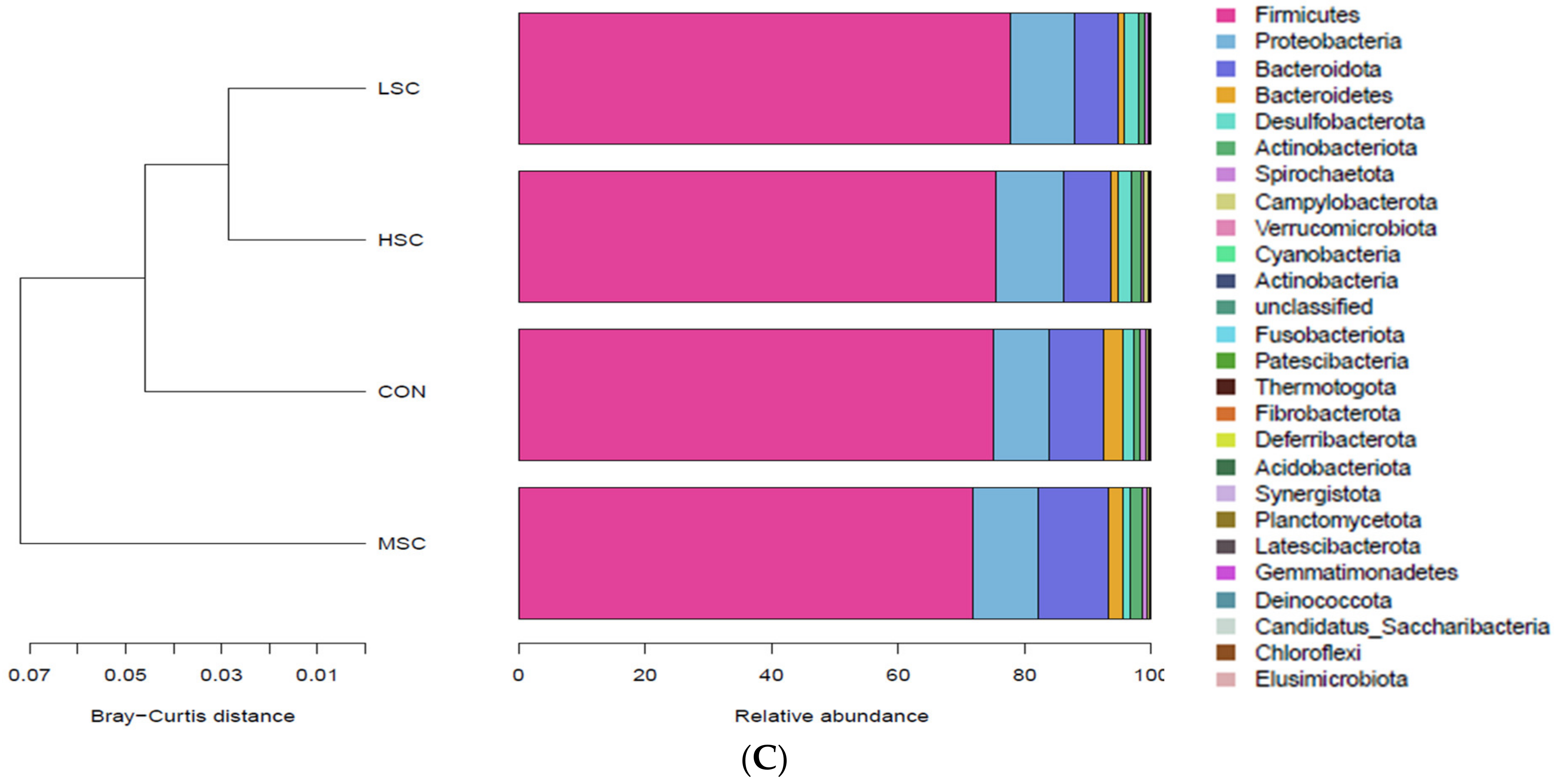
3.5. Cecal Microflora of Weaned Piglets
4. Discussion
4.1. High Efficiency Digestion and Utilization of Organic Iron Made by Saccharomyces cerevisiae
4.2. Effect of Yeast Iron on Antioxidant Function in Weaned Piglets
4.3. Effect of Yeast Iron on Cecal Microbial in Piglets
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hurrell, R. How to ensure adequate iron absorption from iron fortified-food. Nutr. Rev. 2002, 60, S7–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieu, T.P.; Heiskala, M.; Peterson, A.P.; Yang, Y. The roles of iron in health and disease. Mol. Asp. Med. 2001, 22, 1–87. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.L. Iron biology in immune function, muscle metabolism and neuronal functioning. J Nutr. 2001, 131, 568S–580S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luiggi, F.G.; Berto, D.A.; Mello, G.D.; Girão, L.V.C.; Villela, C.C.E.J.; Lo Tierzo, V.; de Trindade Neto, M.A. Relative bioavailability of iron from organic sources for weanling piglets. Semina Ciênc. Agrar. 2014, 35, 2807–2816. [Google Scholar] [CrossRef] [Green Version]
- Thomaz, M.C.; Watanabe, P.H.; Pascoal, L.A.; Assis, M.M.; Ruiz, U.S.; Amorim, A.B.; Robles-Huaynate, R.A. Inorganic and organic trace mineral supplementation in weanling pig diets. An. Acad. Bras. Ciênc. 2015, 87, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Maja, P.; Barbara, P.; Matevq, K.; Peter, R. Iron enriched yeast biomass—A promising mineral feed supplement. Bioresour. Technol. 2007, 98, 1622–1628. [Google Scholar] [CrossRef]
- Manner, K.; Lester, H.; Henriquez-Rodriguez, E. Ferric citrate is a safe and digestible source of iron in broilers and piglets. PeerJ. 2021, 9, e12636. [Google Scholar] [CrossRef]
- Gakh, O.; Adamec, J.; Gacy, A.M.; Twesten, R.D.; Owen, W.G.; Isaya, G. Physical evidence that yeast frataxin is an iron storage protein. Biochemistry 2002, 41, 6798–6804. [Google Scholar] [CrossRef]
- Wang, S.; Wu, S.; Zhang, Y.; Chen, J.; Zhou, X. Effects of different levels of organic trace minerals on oxidative status and intestinal function in weanling piglets. Biol. Trace Elem. Res. 2022, 201, 720–727. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Wei, S.Y.; Wang, Z.L.; Zhu, C.; Hu, S.L.; Zheng, C.T.; Yang, X.F. Effects of different forms of yeast Saccharomyces cerevisiae on growth performance, intestinal development, and systemic immunity in early-weaned piglets. J. Anim. Sci. Biotechnol. 2015, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Czech, A.; Smolczyk, A.; Grela, E.R.; Kiesz, M. Effect of dietary supplementation with Yarrowia lipolytica or Saccharomyces cerevisiae yeast and probiotic additives on growth performance, basic nutrients digestibility and biochemical blood profile in piglets. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1720–1730. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.G.; Wang, N.; Ma, G.D.; Liu, Z.Y.; Wei, G.X.; Liu, W.J. Preparation of S-iron-enriched yeast using siderophores and its effect on iron deficiency anemia in rats. Food Chem. 2021, 365, 130508. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Ghosh, T.K.; Haldar, S. Effects of methionine chelate-or yeast proteinate-based supplement of copper, iron, manganese and zinc on broiler growth performance, their distribution in the tibia and excretion into the environment. Biol. Trace Elem. Res. 2015, 164, 253–260. [Google Scholar] [CrossRef]
- Metges, C.C.; Petzke, K.J.; El-Khoury, A.E.; Henneman, L.; Grant, I.; Bedri, S.; Regan, M.M.; Fuller, M.F.; Young, V.R. Incorporation of urea and ammonia nitrogen into ileal and fecal microbial proteins and plasma free amino acids in normal men and ileostomates. Am. J. Clin. Nutr. 1999, 70, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Warren, M.W.; Paul, J.M.; Malcolm, F.F. Endogenous components of digesta Protein from the terminal ileum of pigs fed a casein-based diet. J. Agric. Food. Chem. 2009, 57, 2072–2078. [Google Scholar] [CrossRef]
- Männer, K.; Simon, O.; Schlegel, P.J.T. Effects of different iron, manganese, zinc and copper sources (sulfates, chelates, glycinates) on their bioavailability in early weaned piglets. In Tagung Schweine-und Geflügelernährung, 9th ed.; Universität Halle-Wittenberg: Halle, Germany, 2006. [Google Scholar]
- Finazzi, D.; Arosio, P. Biology of ferritin in mammals: An update on iron storage, oxidative damage and neurodegeneration. Arch. Toxicol. 2014, 88, 1787–1802. [Google Scholar] [CrossRef] [Green Version]
- Anderson, T.A.; Filer, L.J., Jr.; Fomon, S.J.; Andersen, D.W.; Nixt, T.L.; Rogers, R.R.; Nelson, S.E. Bioavailability of different sources of dietary iron fed to Pitman-Moore miniature pigs. J. Nutr. 1974, 104, 619–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.M.; Sun, X.M.; Wan, D.; Shu, X.G.; Yin, Y.L.; Wu, X. Effects of dipeptide chelated iron on growth performance, serum iron and antioxidant indexes of piglets. Chin. J. Anim. Nutr. 2016, 8, 2551–2555. [Google Scholar] [CrossRef]
- Erikson, K.M.; Pinero, D.J.; Connor, J.R.; Beard, J.L. Regional brain iron, ferritin and transferrin concentrations during iron deficiency and iron repletion in developing rats. J. Nutr. 1997, 127, 2030–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cazzola, M.J.B.P.; Haematology, R.C. Role of ferritin and ferroportin genes in unexplained hyperferritinaemia. Best Pract. Res. Clin. Haematol. 2005, 18, 251–263. [Google Scholar] [CrossRef]
- Yu, S.; West, C.E.; Beynen, A.C. Increasing intakes of iron reduce status, absorption and biliary excretion of copper in rats. Br. J. Nutr. 1994, 71, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Winiarska-Mieczan, A.; Kwiecien, M.; Grela, E.R.; Tomaszewska, E.; Klebaniuk, R. The chemical composition and sensory properties of raw, cooked and grilled thigh meat of broiler chickens fed with Fe-Gly chelate. J. Food Sci. Technol.-Mysore. 2016, 53, 3825–3833. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Qiao, S.; Li, D.; Piao, X.; Ren, J. Effects of Lactobacilli on the performance, diarrhea incidence, VFA concentration and gastrointestinal microbial flora of weaning pigs. Asian Australas. J. Anim. Sci. 2004, 17, 401–409. [Google Scholar] [CrossRef]
- Zha, A.; Yuan, D.; Cui, Z.; Qi, M.; Liao, S.; Liao, P.; Tan, B. The evaluation of the antioxidant and intestinal protective effects of baicalin-copper in deoxynivalenol-challenged piglets. Oxid Med. Cell. Longev. 2020, 2020, 5363546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickens, A.P. Ageing and the free radical theory. Respir. Physiol. 2001, 128, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Gao, W.; Li, Q.; Li, H.; Hao, R. Effect of grapeseed procyanidins on small intestinal mucosa morphology and small intestinal development in weaned piglets. Anim. Prod. Sci. 2020, 60, 1894–1901. [Google Scholar] [CrossRef]
- Nunokawa, Y.; Ishida, N.; Tanaka, S. Promoter analysis of human inducible nitric oxide synthase gene associated with cardiovascular homeostasis. Biochem. Biophys. Res. Commun. 1994, 200, 802–807. [Google Scholar] [CrossRef] [PubMed]
- Man, M.Q.; Wakefield, J.S.; Mauro, T.M.; Elias, P.M. Regulatory Role of Nitric Oxide in Cutaneous Inflammation. Inflammation 2022, 45, 949–964. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Stobberingh, E.E.; Brandt, P.V.D.; Thijs, C. The role of the intestinal microbiota in the development of atopic disorders. Allergy 2007, 62, 1223–1236. [Google Scholar] [CrossRef] [Green Version]
- Lallès, J.P.; Bosi, P.; Smidt, H.; Stokes, C.R. Weaning—A challenge to gut physiologists. Livest. Sci. 2007, 108, 82–93. [Google Scholar] [CrossRef]
- Li, Y.; Guo, Y.; Wen, Z.; Jiang, X.; Ma, X.; Han, X. Weaning stress perturbs gut microbiome and its metabolic profile in piglets. Sci. Rep. 2018, 8, 18068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čoklo, M.; Maslov, D.R.; Kraljević Pavelić, S. Modulation of gut microbiota in healthy rats after exposure to nutritional supplements. Gut Pathog. 2020, 12, 1779002. [Google Scholar] [CrossRef] [PubMed]
- Bervoets, L.; Van Hoorenbeeck, K.; Kortleven, I.; Van Noten, C.; Hens, N.; Vael, C.; Vankerckhoven, V. Differences in gut microbiota composition between obese and lean children: A cross-sectional study. Gut Pathog. 2013, 5, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [Green Version]
- Rasback, T.; Jansson, D.S.; Johansson, K.E.; Fellstrom, C. A novel enteropathogenic, strongly haemolytic spirochaete isolated from pig and mallard, provisionally designated ‘Brachyspira suanatina’ sp nov. Environ. Microbiol. 2007, 9, 983–991. [Google Scholar] [CrossRef]
- Murray, R.D.; Downham, D.Y.; Demirkan, I.; Carter, S.D. Some relationships between spirochaete infections and digital dermatitis in four UK dairy herds. Res. Vet. Sci. 2002, 73, 223–230. [Google Scholar] [CrossRef]
- Kassinen, A.; Krogius-Kurikka, L.; Mäkivuokko, H.; Rinttilä, T.; Paulin, L.; Corander, J.; Palva, A. The fecal microbiota of irritable bowel syndrome patients differs significantly from that of healthy subjects. Gastroenterology 2007, 133, 24–33. [Google Scholar] [CrossRef]
- Benítez-Páez, A.; Gómez del Pugar, E.M.; López-Almela, I.; Moya-Pérez, Á.; Codoñer-Franch, P.; Sanz, Y. Depletion of Blautia species in the microbiota of obese children relates to intestinal inflammation and metabolic phenotype worsening. mSystems 2020, 5, e00857-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Ingredients | Content | Nutritional Level | Content |
|---|---|---|---|
| Corn | 61.49 | Digestive energy (MJ/kg) | 14.34 |
| Soybean meal | 12.00 | Crude protein | 18.42 |
| Puffed soybean | 6.08 | Calcium | 0.78 |
| Fish meal | 5.00 | Total Phosphorus | 0.67 |
| Whey powder | 10.00 | Available Phosphorus | 0.40 |
| Fat powder | 1.59 | Lysine | 1.42 |
| Calcium hydrogen phosphate | 0.93 | Methionine | 0.51 |
| Limestone | 0.64 | Methionine + Cystine | 0.78 |
| Salt | 0.30 | Threonine | 0.90 |
| L-Lysine hydrochloride | 0.57 | Tryptophan | 0.21 |
| Methionine | 0.21 | Crude fiber | 2.50 |
| Threonine | 0.20 | Dry matter | 94.75 |
| Premix1 | 1.00 | ||
| Total | 100.00 |
| Items | Treatments 1 | SEM | p Value | |||
|---|---|---|---|---|---|---|
| CON | LSC | MSC | HSC | |||
| Fecal iron content (mg/kg) | 2816.16 a | 2107.45 b | 2157.80 b | 2153.79 b | 78.33 | <0.01 |
| The apparent digestibility of iron (%) | 73.46 c | 80.25 b | 86.93 a | 87.74 a | 1.54 | <0.01 |
| The apparent digestibility of crude protein (%) | 90.09 | 89.66 | 90.15 | 90.32 | 0.226 | 0.797 |
| The apparent digestibility of dry matter | 90.15 | 91.13 | 91.09 | 91.52 | 0.0092 | 0.188 |
| Items | Treatments 1 | SEM | p Value | |||
|---|---|---|---|---|---|---|
| CON | LSC | MSC | HSC | |||
| Serum iron content (μmol/L) | 84.38 b | 109.94 a | 110.47 a | 111.35 a | 3.36 | <0.01 |
| Total iron binding capacity (μmol/L) | 535.96 | 546.91 | 552.83 | 553.58 | 3.40 | 0.239 |
| Ferritin (ng/mL) | 66.91 | 68.71 | 70.95 | 71.30 | 1.08 | 0.482 |
| Transferrin (g/L) | 6.39 | 6.43 | 6.33 | 6.48 | 0.091 | 0.953 |
| Ceruloplasmin (U/mL) | 16.98 | 17.31 | 17.73 | 17.97 | 0.248 | 0.547 |
| The iron content | ||||||
| Heart (mg/kg) | 184.78 b | 184.77 b | 197.82 a | 202.39 a | 3.82 | <0.01 |
| Liver (mg/kg) | 623.24 b | 620.48 b | 695.89 a | 709.83 a | 23.23 | 0.018 |
| Spleen (mg/kg) | 749.46 | 746.47 | 820.06 | 832.89 | 15.28 | 0.061 |
| Lung (mg/kg) | 341.70 b | 435.67 a | 442.76 a | 449.37 a | 11.21 | <0.01 |
| Kidney (mg/kg) | 211.43 b | 217.57 b | 290.10 a | 307.60 a | 14.78 | 0.017 |
| Left gluteus muscle (mg/kg) | 48.59 b | 46.38 b | 55.04 a | 57.48 a | 1.68 | 0.039 |
| Left longissimus (mg/kg) | 45.79 | 48.33 | 54.06 | 58.31 | 2.000 | 0.097 |
| Items | Treatments 1 | SEM | p Value | |||
|---|---|---|---|---|---|---|
| CON | LSC | MSC | HSC | |||
| The pH of gastric juice | 4.98 a | 2.25 b | 2.80 b | 1.95 b | 0.354 | <0.01 |
| The pH of duodenal contents | 5.80 | 5.50 | 4.80 | 5.45 | 0.289 | 0.705 |
| The pH of jejunal contents | 6.20 | 6.15 | 6.88 | 6.20 | 0.144 | 0.234 |
| The pH of ileal contents | 7.45 | 7.38 | 7.28 | 6.93 | 0.043 | 0.107 |
| The pH of cecal contents | 6.43 | 6.35 | 6.18 | 6.30 | 0.049 | 0.355 |
| The pH of colonic contents | 6.85 | 6.88 | 6.90 | 7.05 | 0.087 | 0.878 |
| Items | Treatments 1 | SEM | p Value | |||
|---|---|---|---|---|---|---|
| CON | LSC | MSC | HSC | |||
| Catalase (U/mL) | 24.14 b | 25.70 b | 31.96 a | 27.84 a | 0.997 | 0.013 |
| Glutathione peroxidase(U/mL) | 845.08 b | 879.53 b | 940.29 a | 947.97 a | 12.90 | <0.01 |
| Superoxide dismutase (U/mL) | 338.57 b | 343.31 b | 453.49 a | 465.94 a | 17.38 | <0.01 |
| Xanthine oxidase (U/mL) | 5.89 b | 6.58 ab | 7.10 a | 7.27 a | 0.217 | <0.01 |
| Peroxidase (U/mL) | 12.99 b | 12.60 b | 19.62 a | 18.19 a | 0.936 | 0.021 |
| Superoxide anion removal rate (%) | 18.29 b | 18.64 b | 19.30 a | 19.83 a | 0.206 | 0.019 |
| Hydroxyl free radical removal rate (%) | 3.84 b | 3.97 b | 4.43 a | 4.54 a | 0.092 | <0.01 |
| Nitric oxide (μmoL/mL) | 0.239 a | 0.265 a | 0.226 b | 0.173 b | 0.013 | 0.044 |
| Glutathione (μmol/mL) | 0.216 b | 0.225 b | 0.256 a | 0.272 a | 0.016 | <0.01 |
| Oxidized glutathione (nmol/mL) | 2.71 | 2.72 | 1.90 | 2.10 | 0.161 | 0.155 |
| Total antioxidant capacity (μmol Trolox/mL) | 0.118 b | 0.127 b | 0.206 a | 0.201 a | 0.005 | 0.015 |
| Items | Treatments 1 | SEM | p Value | |||
|---|---|---|---|---|---|---|
| CON | LSC | MSC | HSC | |||
| Liver | ||||||
| Peroxidase (U/mg protein) | 6.62 b | 6.69 b | 9.86 a | 10.56 a | 0.537 | <0.01 |
| Glutathione synthase (U/g) | 0.560 | 0.565 | 0.608 | 0.637 | 0.017 | 0.352 |
| Heme oxygenase (U/g) | 0.245 | 0.248 | 0.251 | 0.291 | 0.007 | 0.055 |
| Sulfur redox protein (ng/g) | 1452.08 | 1486.11 | 1485.66 | 1553.43 | 38.52 | 0.853 |
| Nitric oxide (μ moL/mg protein) | 0.114 | 0.115 | 0.104 | 0.106 | 0.002 | 0.150 |
| Superoxide anion removal rate (%) | 17.78 b | 17.82 b | 19.32 a | 19.07 a | 0.139 | 0.0135 |
| Hydroxyl free radical scavenging rate (%) | 3.89 | 4.31 | 4.41 | 4.02 | 0.075 | 0.086 |
| Jejunal mucosa | ||||||
| Peroxidase (U/mg protein) | 4.08 b | 8.16 a | 8.21 a | 8.26 a | 0.550 | <0.01 |
| Glutathione synthase (U/g) | 0.623 | 0.641 | 0.634 | 0.668 | 0.013 | 0.696 |
| Heme oxygenase (U/g) | 0.325 | 0.332 | 0.345 | 0.371 | 0.010 | 0.426 |
| Superoxide anion removal rate (%) | 17.85 b | 18.53 a | 18.85 a | 18.90 a | 0.141 | 0.020 |
| Hydroxyl free radical scavenging rate (%) | 4.20 | 4.25 | 4.64 | 4.62 | 0.080 | 0.074 |
| Nitric oxide (μmoL/mg protein) | 0.136 | 0.130 | 0.127 | 0.128 | 0.007 | 0.973 |
| Sulfur redox protein (μg/g) | 1.07 b | 1.16 b | 1.42 a | 1.49 a | 0.049 | <0.01 |
| Items | Treatments 1 | SEM | p Value | ||||
|---|---|---|---|---|---|---|---|
| CON | LSC | MSC | HSC | ||||
| Alpha diversity index | |||||||
| Observed_otus | 537.00 c | 603.50 ab | 562.75 bc | 634.50 a | 13.20 | 0.023 | |
| Shannon | 7.19 | 6.54 | 7.08 | 7.23 | 0.183 | 0.561 | |
| Simpson | 0.967 | 0.928 | 0.978 | 0.972 | 0.010 | 0.318 | |
| Chao1 | 563.37 | 606.13 | 563.81 | 636.83 | 12.19 | 0.072 | |
| Pielou_e | 0.782 | 0.709 | 0.782 | 0.778 | 0.017 | 0.354 | |
| Phylum | |||||||
| Firmicutes | 73.91 b | 78.93 a | 79.33 a | 79.22 a | 0.592 | <0.01 | |
| Bacteroidetes | 15.02 a | 11.25 b | 8.94 c | 8.74 c | 0.664 | <0.01 | |
| Proteobacteria | 8.54 a | 7.80 a | 9.89 a | 9.97 a | 0.260 | <0.01 | |
| Actinobacteria | 1.28 | 1.33 | 1.43 | 1.39 | 0.036 | 0.495 | |
| Spirochaetota | 0.890 a | 0.343 b | 0.151 c | 0.316 b | 0.074 | <0.01 | |
| Tenericutes | 0.202 a | 0.128 b | 0.062 c | 0.148 b | 0.015 | <0.01 | |
| Genus | |||||||
| Escherichia-Shigella | 1.97 | 1.45 | 1.90 | 1.80 | 0.112 | 0.403 | |
| Acinetobacter | 0.124 a | 0.040 b | 0.034 b | 0.025 b | 0.011 | <0.01 | |
| Streptococcus | 7.78 a | 4.72 b | 4.47 b | 4.31 b | 0.390 | <0.01 | |
| Prevotella | 11.78 a | 7.93 c | 8.21 bc | 9.43 b | 0.433 | <0.01 | |
| Staphylococcus | 0.392 | 0.366 | 0.404 | 0.299 | 0.028 | 0.623 | |
| Clostridium | 10.24 | 10.61 | 11.15 | 9.73 | 0.203 | 0.070 | |
| Veillonellaceae | 4.56 | 4.25 | 4.30 | 4.15 | 0.095 | 0.510 | |
| Bacteroides | 0.408 b | 0.316 b | 0.361 b | 0.650 a | 0.039 | <0.01 | |
| Lactobacillus | 3.39 b | 3.32 b | 4.39 a | 4.76 a | 0.175 | <0.01 | |
| Blautia | 0.246 b | 0.399 a | 0.459 a | 0.425 a | 0.025 | <0.01 | |
| Peptococcus | 0.036 c | 0.103 b | 0.104 b | 0.155 a | 0.012 | <0.01 | |
| Lactococcus | 0.566 | 0.518 | 0.550 | 0.547 | 0.035 | 0.978 | |
| Bifidobacterium | 0.141 | 0.138 | 0.173 | 0.210 | 0.015 | 0.299 | |
| Lachnospiraceae | 10.88 | 12.34 | 12.04 | 11.46 | 0.259 | 0.198 | |
| Roseburia | 0.785 | 0.960 | 1.048 | 1.31 | 0.082 | 0.140 | |
| Ruminococcus | 3.71 | 3.50 | 3.69 | 4.23 | 0.152 | 0.392 | |
| Faecalibacterium | 0.497 | 0.581 | 0.756 | 1.17 | 0.093 | 0.026 | |
| Eubacterium | 3.00 | 3.34 | 3.53 | 3.71 | 0.139 | 0.318 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.; Jiang, L.; Zhou, B.; Liu, Y.; Wang, L.; Hu, Z.; Wang, C.; Tang, Z. Effect of High Efficiency Digestion and Utilization of Organic Iron Made by Saccharomyces cerevisiae on Antioxidation and Caecum Microflora in Weaned Piglets. Animals 2023, 13, 498. https://doi.org/10.3390/ani13030498
Zeng Y, Jiang L, Zhou B, Liu Y, Wang L, Hu Z, Wang C, Tang Z. Effect of High Efficiency Digestion and Utilization of Organic Iron Made by Saccharomyces cerevisiae on Antioxidation and Caecum Microflora in Weaned Piglets. Animals. 2023; 13(3):498. https://doi.org/10.3390/ani13030498
Chicago/Turabian StyleZeng, Yan, Liwen Jiang, Bingyu Zhou, Yubo Liu, Lingang Wang, Zhijin Hu, Chunping Wang, and Zhiru Tang. 2023. "Effect of High Efficiency Digestion and Utilization of Organic Iron Made by Saccharomyces cerevisiae on Antioxidation and Caecum Microflora in Weaned Piglets" Animals 13, no. 3: 498. https://doi.org/10.3390/ani13030498