
Epigenetic Alterations in Canine Malignant Lymphoma: Future and Clinical Outcomes

Abstract
:Simple Summary
Abstract
1. Introduction
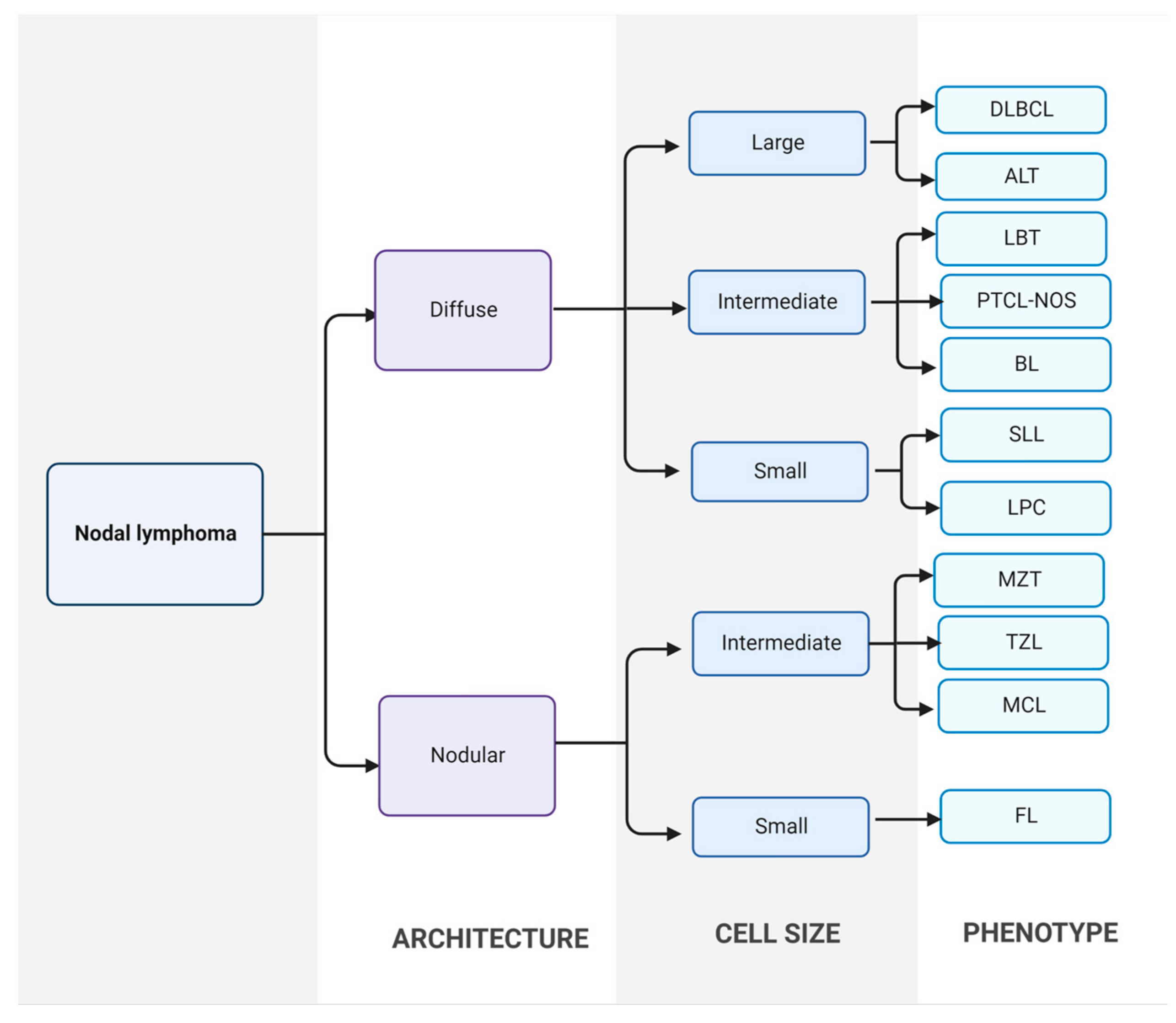
2. Canine Malignant Lymphomas (CML)
2.1. Aetiology of Lymphomas
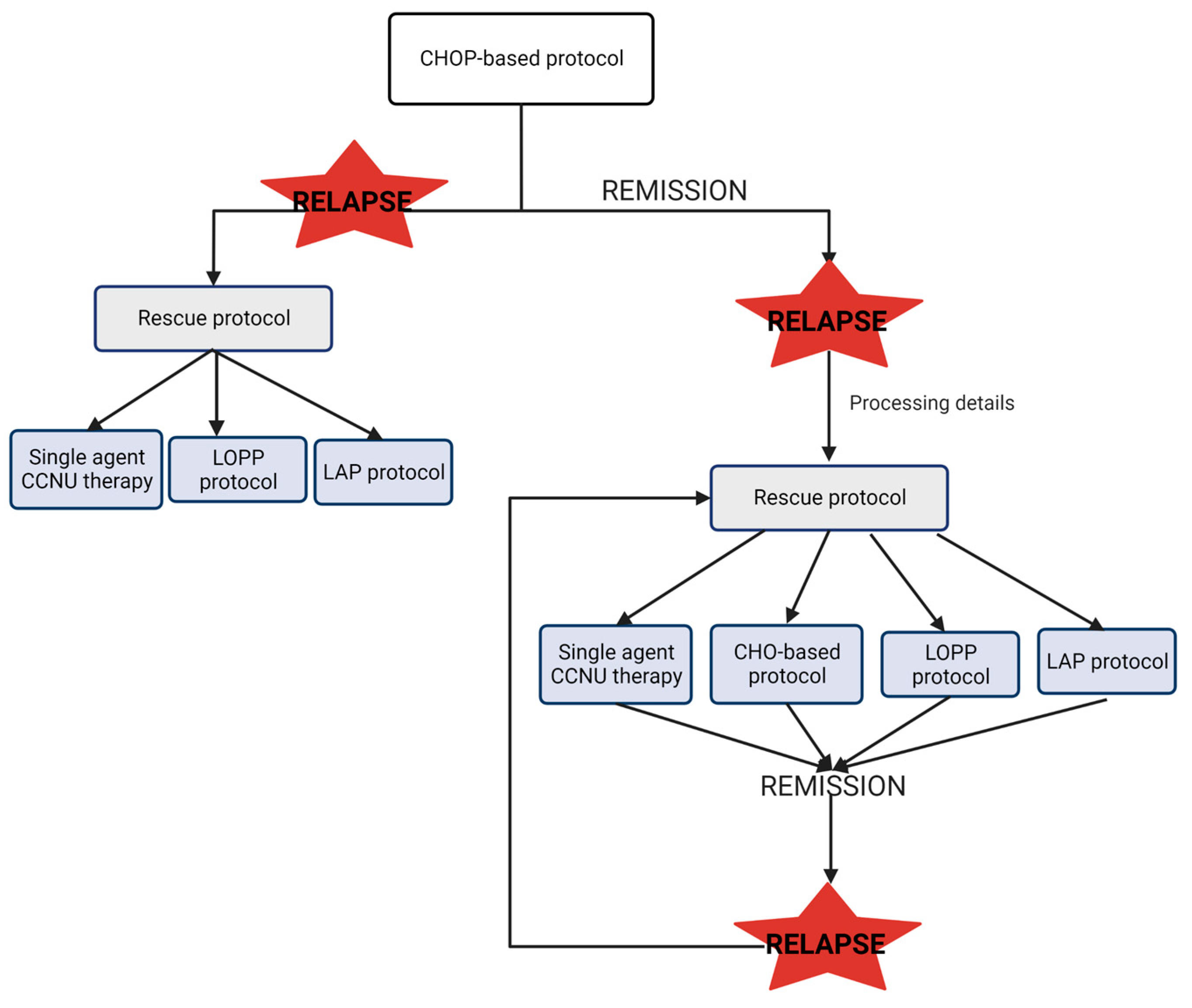
2.2. Diagnosis, Treatment and Prognosis of CML
3. Epigenetic Mechanisms and CML
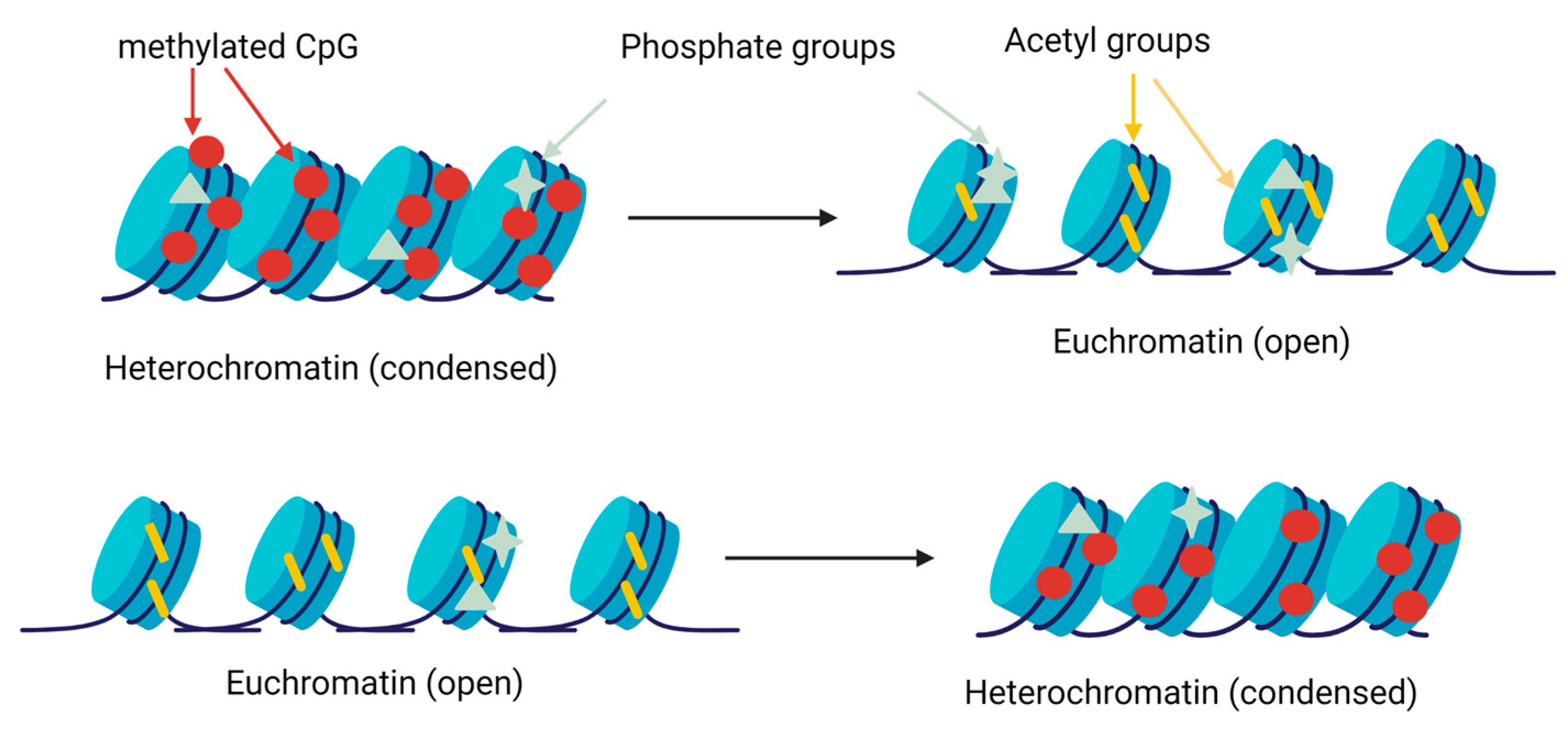
3.1. Histone Modifications
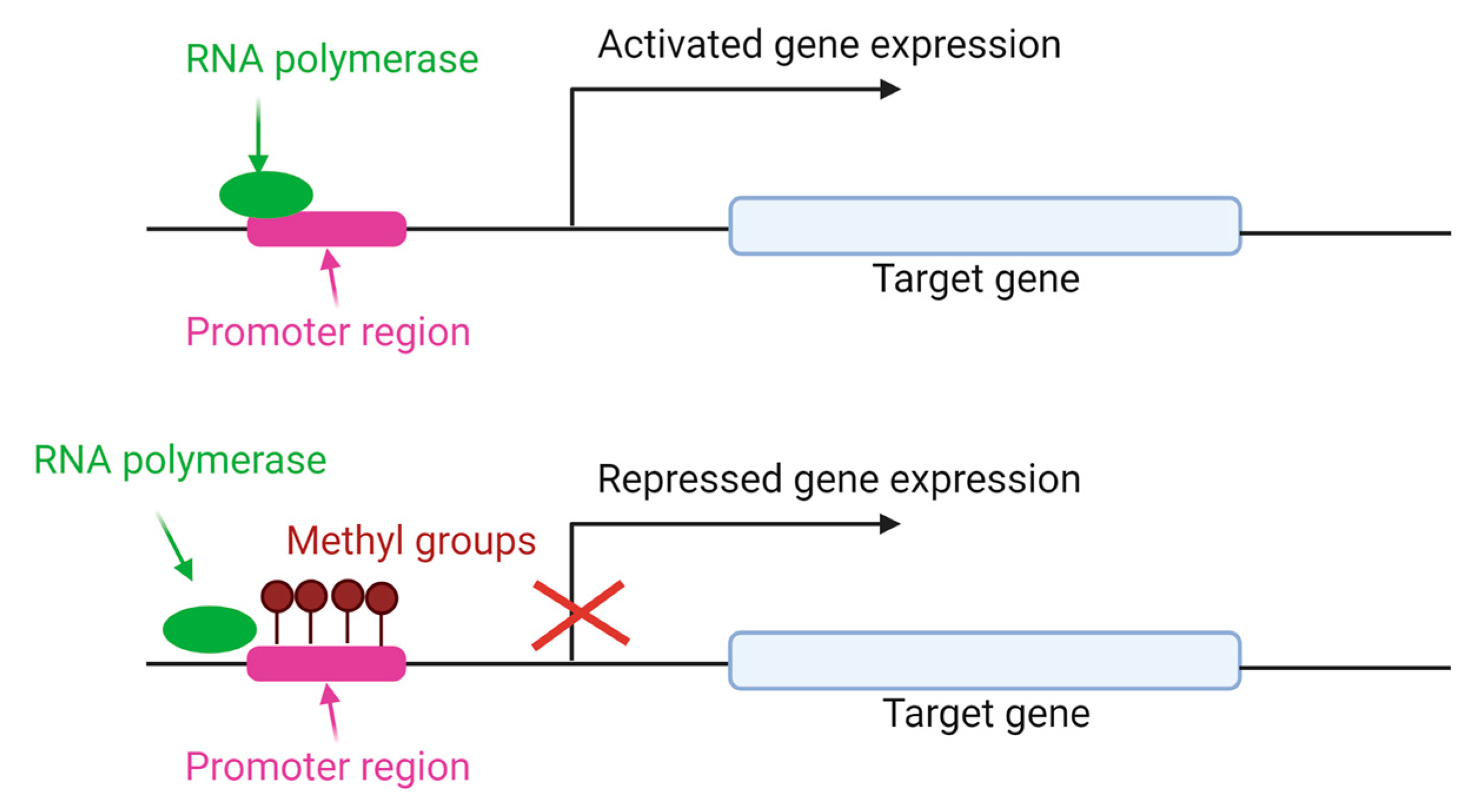
3.2. DNA Methylation
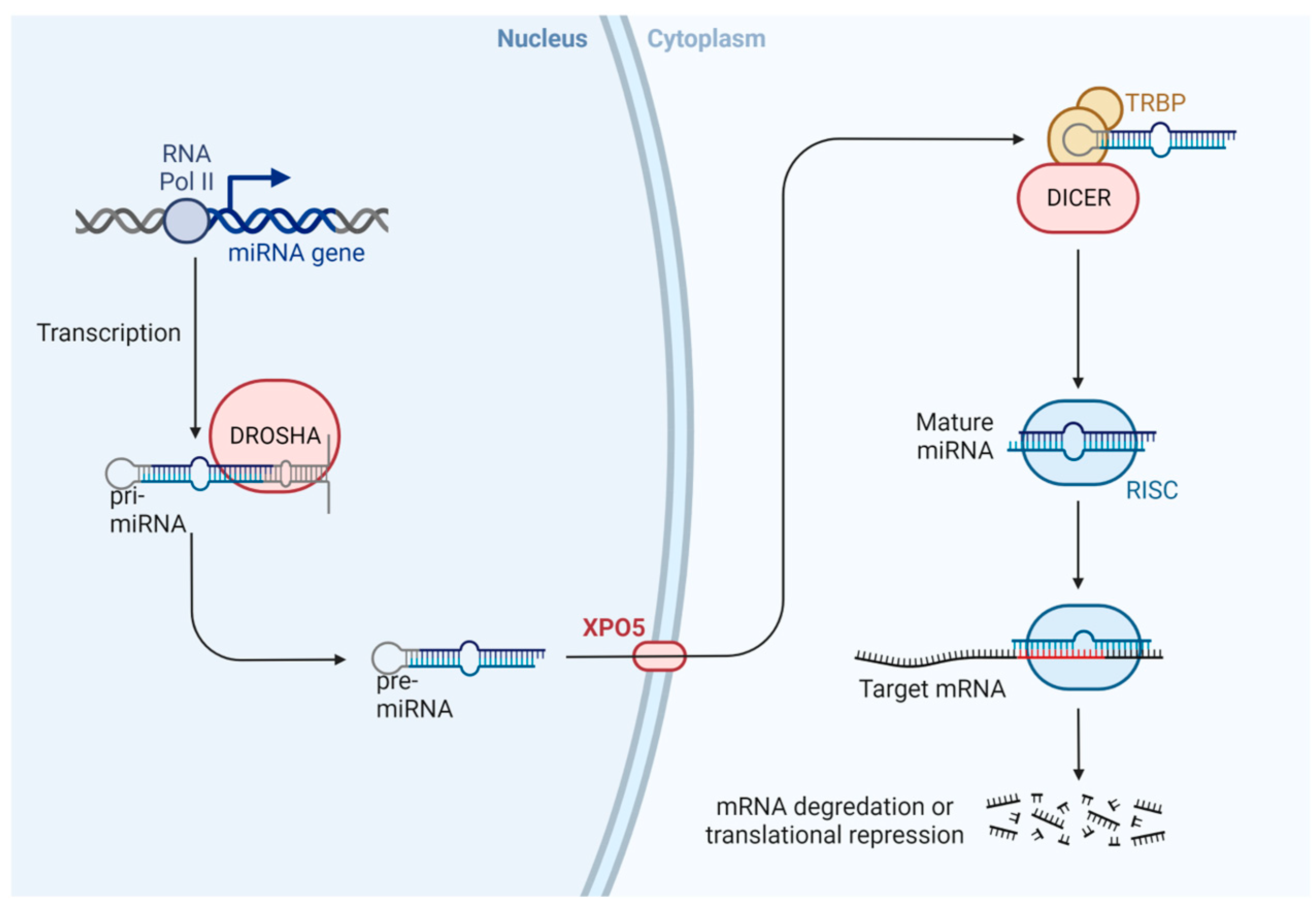
3.3. MicroRNAs (miRNAs)
4. New Treatments and Biomarkers Based on Epigenetics in CML
4.1. Histone Deacetylase (HDAC) Inhibitors as Treatment
4.2. Demethylation and Deacetylation Drugs as Treatment for Canine Lymphoma
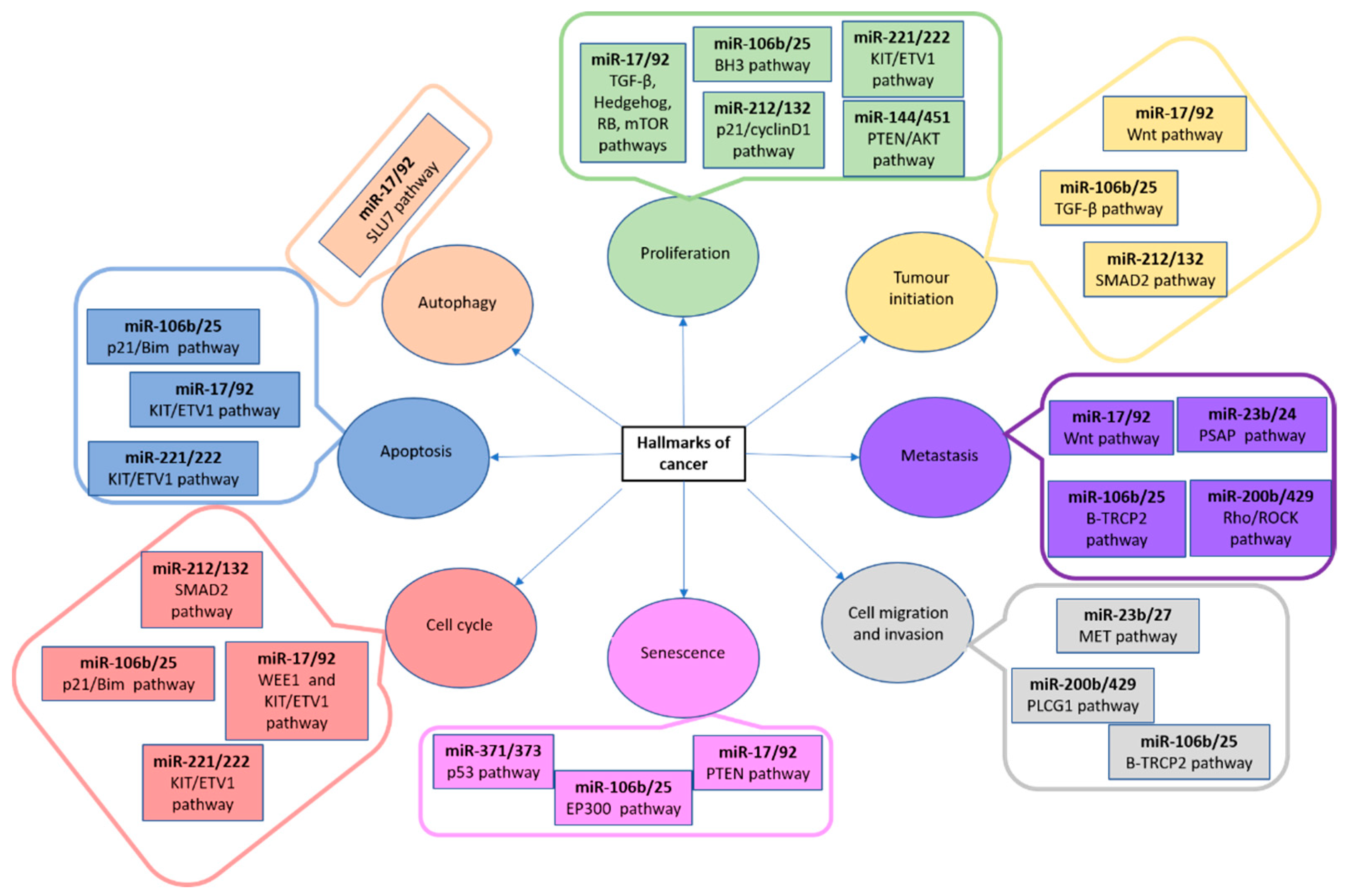
4.3. microRNAs as Biomarkers and Treatment in CML
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobson, J.M.; Samuel, S.; Milstein, H.; Rogers, K.; Wood, J.L.N. Canine Neoplasia in the UK: Estimates of Incidence Rates from a Population of Insured Dogs. J. Small Anim. Pract. 2002, 43, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Van Rooyen, L.J.; Hooijberg, E.; Reyers, F. Breed Prevalence of Canine Lymphoma in South Africa. J. S. Afr. Vet. Assoc. 2018, 89, e1–e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modiano, J.F.; Breen, M.; Burnett, R.C.; Parker, H.G.; Inusah, S.; Thomas, R.; Avery, P.R.; Lindblad-Toh, K.; Ostrander, E.A.; Cutter, G.C.; et al. Distinct B-Cell and T-Cell Lymphoproliferative Disease Prevalence among Dog Breeds Indicates Heritable Risk. Cancer Res. 2005, 65, 5654–5661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comazzi, S.; Marelli, S.; Cozzi, M.; Rizzi, R.; Finotello, R.; Henriques, J.; Pastor, J.; Ponce, F.; Rohrer-Bley, C.; Rütgen, B.C.; et al. Breed-Associated Risks for Developing Canine Lymphoma Differ among Countries: An European Canine Lymphoma Network Study. BMC Vet. Res. 2018, 14, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valli, V.E.; San Myint, M.; Barthel, A.; Bienzle, D.; Caswell, J.; Colbatzky, F.; Durham, A.; Ehrhart, E.J.; Johnson, Y.; Jones, C.; et al. Classification of Canine Malignant Lymphomas According to the World Health Organization Criteria. Vet. Pathol. 2011, 48, 198–211. [Google Scholar] [CrossRef] [Green Version]
- Zandvliet, M. Canine Lymphoma: A Review. Vet. Q. 2016, 36, 76–104. [Google Scholar] [CrossRef]
- Ponce, F.; Marchal, T.; Magnol, J.P.; Turinelli, V.; Ledieu, D.; Bonnefont, C.; Pastor, M.; Delignette, M.L.; Fournel-Fleury, C. A Morphological Study of 608 Cases of Canine Malignant Lymphoma in France with a Focus on Comparative Similarities between Canine and Human Lymphoma Morphology. Vet. Pathol. 2010, 47, 414–433. [Google Scholar] [CrossRef]
- Kluin, P.; Schuuring, E. Molecular Cytogenetics of Lymphoma: Where Do We Stand in 2010? Histopathology 2011, 58, 128–144. [Google Scholar] [CrossRef] [Green Version]
- Teske, E. Canine Malignant Lymphoma: A Review and Comparison with Human Non-Hodgkin’s Lymphoma. Vet. Q. 1994, 16, 209–219. [Google Scholar] [CrossRef]
- Vail, D.M.; MacEwen, E.G. Spontaneously Occurring Tumors of Companion Animals as Models for Human Cancer. Cancer Investig. 2000, 18, 781–792. [Google Scholar] [CrossRef]
- Yau, P.; Dhand, N.K.; Thomson, P.C.; Taylor, R.M. Retrospective Study on the Occurrence of Canine Lymphoma and Associated Breed Risks in a Population of Dogs in NSW (2001–2009). Aust. Vet. J. 2017, 95, 149–155. [Google Scholar] [CrossRef]
- Thandra, K.C.; Barsouk, A.; Saginala, K.; Padala, S.A.; Barsouk, A.; Rawla, P. Epidemiology of Non-Hodgkin’s Lymphoma. Med. Sci. 2021, 9, 5. [Google Scholar] [CrossRef]
- Alexander, D.D.; Mink, P.J.; Adami, H.-O.; Chang, E.T.; Cole, P.; Mandel, J.S.; Trichopoulos, D. The Non-Hodgkin Lymphomas: A Review of the Epidemiologic Literature. Int. J. Cancer 2007, 120 (Suppl. S12), 1–39. [Google Scholar] [CrossRef]
- Horesh, N.; Horowitz, N.A. Does Gender Matter in Non-Hodgkin Lymphoma? Differences in Epidemiology, Clinical Behavior, and Therapy. Rambam. Maimonides Med. J. 2014, 5, e0038. [Google Scholar] [CrossRef]
- Villamil, J.A.; Henry, C.J.; Hahn, A.W.; Bryan, J.N.; Tyler, J.W.; Caldwell, C.W. Hormonal and Sex Impact on the Epidemiology of Canine Lymphoma. J. Cancer Epidemiol. 2009, 2009, 591753. [Google Scholar] [CrossRef] [Green Version]
- Pierdominici, M.; Maselli, A.; Colasanti, T.; Giammarioli, A.M.; Delunardo, F.; Vacirca, D.; Sanchez, M.; Giovannetti, A.; Malorni, W.; Ortona, E. Estrogen Receptor Profiles in Human Peripheral Blood Lymphocytes. Immunol. Lett. 2010, 132, 79–85. [Google Scholar] [CrossRef]
- Rachoń, D.; Myśliwska, J.; Suchecka-Rachoń, K.; Wieckiewicz, J.; Myśliwski, A. Effects of Oestrogen Deprivation on Interleukin-6 Production by Peripheral Blood Mononuclear Cells of Postmenopausal Women. J. Endocrinol. 2002, 172, 387–395. [Google Scholar] [CrossRef] [Green Version]
- Preti, H.A.; Cabanillas, F.; Talpaz, M.; Tucker, S.L.; Seymour, J.F.; Kurzrock, R. Prognostic Value of Serum Interleukin-6 in Diffuse Large-Cell Lymphoma. Ann. Intern. Med. 1997, 127, 186–194. [Google Scholar] [CrossRef]
- Mizutani, N.; Goto-Koshino, Y.; Takahashi, M.; Uchida, K.; Tsujimoto, H. Clinical and Histopathological Evaluation of 16 Dogs with T-Zone Lymphoma. J. Vet. Med. Sci. 2016, 78, 1237–1244. [Google Scholar] [CrossRef] [Green Version]
- Seelig, D.M.; Avery, P.; Webb, T.; Yoshimoto, J.; Bromberek, J.; Ehrhart, E.J.; Avery, A.C. Canine T-Zone Lymphoma: Unique Immunophenotypic Features, Outcome, and Population Characteristics. J. Vet. Intern. Med. 2014, 28, 878–886. [Google Scholar] [CrossRef]
- Jankowska, U.; Jagielski, D.; Czopowicz, M.; Sapierzyński, R. The Animal-Dependent Risk Factors in Canine T-Cell Lymphomas. Vet. Comp. Oncol. 2017, 15, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Pastor, M.; Chalvet-Monfray, K.; Marchal, T.; Keck, G.; Magnol, J.P.; Fournel-Fleury, C.; Ponce, F. Genetic and Environmental Risk Indicators in Canine Non-Hodgkin’s Lymphomas: Breed Associations and Geographic Distribution of 608 Cases Diagnosed throughout France over 1 Year. J. Vet. Intern. Med. 2009, 23, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Vail, D.M.; Thamm, D.H.; Liptak, J.M. Hematopoietic Tumors. In Withrow and MacEwen’s Small Animal Clinical Oncology; Elsevier Health Sciences: Amsterdam, The Netherlands, 2019; pp. 688–772. [Google Scholar] [CrossRef]
- Lane, J.; Price, J.; Moore, A.; Dandrieux, J.R.S.; Clifford, C.; Curran, K.; Choy, K.; Cannon, C. Low-Grade Gastrointestinal Lymphoma in Dogs: 20 Cases (2010 to 2016). J. Small Anim. Pract. 2018, 59, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.L.; Vernau, W.; Rebhun, R.B.; Skorupski, K.A.; Burton, J.H. Patient Characteristics, Prognostic Factors and Outcome of Dogs with High-Grade Primary Mediastinal Lymphoma. Vet. Comp. Oncol. 2018, 16, E45–E51. [Google Scholar] [CrossRef]
- Fontaine, J.; Bovens, C.; Bettenay, S.; Mueller, R.S. Canine Cutaneous Epitheliotropic T-Cell Lymphoma: A Review. Vet. Comp. Oncol. 2009, 7, 1–14. [Google Scholar] [CrossRef]
- Ponce, F.; Magnol, J.-P.; Ledieu, D.; Marchal, T.; Turinelli, V.; Chalvet-Monfray, K.; Fournel-Fleury, C. Prognostic Significance of Morphological Subtypes in Canine Malignant Lymphomas during Chemotherapy. Vet. J. 2004, 167, 158–166. [Google Scholar] [CrossRef]
- Valli, V.E.; Kass, P.H.; San Myint, M.; Scott, F. Canine Lymphomas: Association of Classification Type, Disease Stage, Tumor Subtype, Mitotic Rate, and Treatment with Survival. Vet. Pathol. 2013, 50, 738–748. [Google Scholar] [CrossRef]
- Seelig, D.M.; Avery, A.C.; Ehrhart, E.J.; Linden, M.A. The Comparative Diagnostic Features of Canine and Human Lymphoma. Vet. Sci. 2016, 3, 11. [Google Scholar] [CrossRef]
- Riondato, F.; Comazzi, S. Flow Cytometry in the Diagnosis of Canine B-Cell Lymphoma. Front. Vet. Sci. 2021, 8, 600986. [Google Scholar] [CrossRef]
- Thalheim, L.; Williams, L.E.; Borst, L.B.; Fogle, J.E.; Suter, S.E. Lymphoma Immunophenotype of Dogs Determined by Immunohistochemistry, Flow Cytometry, and Polymerase Chain Reaction for Antigen Receptor Rearrangements. J. Vet. Intern. Med. 2013, 27, 1509–1516. [Google Scholar] [CrossRef]
- Waugh, E.M.; Gallagher, A.; Haining, H.; Johnston, P.E.J.; Marchesi, F.; Jarrett, R.F.; Morris, J.S. Optimisation and Validation of a PCR for Antigen Receptor Rearrangement (PARR) Assay to Detect Clonality in Canine Lymphoid Malignancies. Vet. Immunol. Immunopathol. 2016, 182, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Ehrhart, E.J.; Wong, S.; Richter, K.; Zismann, V.; Grimes, C.; Hendricks, W.; Khanna, C. Polymerase Chain Reaction for Antigen Receptor Rearrangement: Benchmarking Performance of a Lymphoid Clonality Assay in Diverse Canine Sample Types. J. Vet. Intern. Med. 2019, 33, 1392–1402. [Google Scholar] [CrossRef] [Green Version]
- Burnett, R.C.; Vernau, W.; Modiano, J.F.; Olver, C.S.; Moore, P.F.; Avery, A.C. Diagnosis of Canine Lymphoid Neoplasia Using Clonal Rearrangements of Antigen Receptor Genes. Vet. Pathol. 2003, 40, 32–41. [Google Scholar] [CrossRef]
- Moore, A.S. Treatment of T Cell Lymphoma in Dogs. Vet. Rec. 2016, 179, 277. [Google Scholar] [CrossRef]
- Saba, C.F.; Hafeman, S.D.; Vail, D.M.; Thamm, D.H. Combination Chemotherapy with Continuous L-Asparaginase, Lomustine, and Prednisone for Relapsed Canine Lymphoma. J. Vet. Intern. Med. 2009, 23, 1058–1063. [Google Scholar] [CrossRef]
- Brown, P.M.; Tzannes, S.; Nguyen, S.; White, J.; Langova, V. LOPP Chemotherapy as a First-Line Treatment for Dogs with T-Cell Lymphoma. Vet. Comp. Oncol. 2018, 16, 108–113. [Google Scholar] [CrossRef]
- Lautscham, E.M.; Kessler, M.; Ernst, T.; Willimzig, L.; Neiger, R. Comparison of a CHOP-LAsp-Based Protocol with and without Maintenance for Canine Multicentric Lymphoma. Vet. Rec. 2017, 180, 303. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of Chromatin by Histone Modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef]
- Copeland, R.A.; Solomon, M.E.; Richon, V.M. Protein Methyltransferases as a Target Class for Drug Discovery. Nat. Rev. Drug Discov. 2009, 8, 724–732. [Google Scholar] [CrossRef]
- Xhemalce, B.; Dawson, M.A.; Bannister, A.J. Histone Modifications. In Reviews in Cell Biology and Molecular Medicine; American Cancer Society: Atlanta, GA, USA, 2011; ISBN 978-3-527-60090-8. [Google Scholar]
- Oki, M.; Aihara, H.; Ito, T. Role of Histone Phosphorylation in Chromatin Dynamics and Its Implications in Diseases. Subcell Biochem. 2007, 41, 319–336. [Google Scholar]
- Smith, Z.D.; Meissner, A. DNA Methylation: Roles in Mammalian Development. Nat. Rev. Genet. 2013, 14, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Jambhekar, A.; Dhall, A.; Shi, Y. Roles and Regulation of Histone Methylation in Animal Development. Nat. Rev. Mol. Cell Biol. 2019, 20, 625–641. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Serrano, M.; Winkler, R.; Santos, J.C.; Le Pannérer, M.-M.; Buschbeck, M.; Roué, G. Histone Modifications and Their Targeting in Lymphoid Malignancies. Int. J. Mol. Sci. 2021, 23, 253. [Google Scholar] [CrossRef] [PubMed]
- Shaknovich, R.; Melnick, A. Epigenetics and B-Cell Lymphoma. Curr. Opin. Hematol. 2011, 18, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, P.; Wang, Y. Epigenetics of Cutaneous T-Cell Lymphoma: Biomarkers and Therapeutic Potentials. Cancer Biol. Med. 2021, 18, 34–51. [Google Scholar] [CrossRef]
- Ji, M.-M.; Huang, Y.-H.; Huang, J.-Y.; Wang, Z.-F.; Fu, D.; Liu, H.; Liu, F.; Leboeuf, C.; Wang, L.; Ye, J.; et al. Histone Modifier Gene Mutations in Peripheral T-Cell Lymphoma Not Otherwise Specified. Haematologica 2018, 103, 679–687. [Google Scholar] [CrossRef]
- Lemercier, C.; Brocard, M.-P.; Puvion-Dutilleul, F.; Kao, H.-Y.; Albagli, O.; Khochbin, S. Class II Histone Deacetylases Are Directly Recruited by BCL6 Transcriptional Repressor. J. Biol. Chem. 2002, 277, 22045–22052. [Google Scholar] [CrossRef] [Green Version]
- Yoshimitsu, M.; Ando, K.; Ishida, T.; Yoshida, S.; Choi, I.; Hidaka, M.; Takamatsu, Y.; Gillings, M.; Lee, G.T.; Onogi, H.; et al. Oral Histone Deacetylase Inhibitor HBI-8000 (Tucidinostat) in Japanese Patients with Relapsed or Refractory Non-Hodgkin’s Lymphoma: Phase I Safety and Efficacy. Jpn. J. Clin. Oncol. 2022, 52, 1014–1020. [Google Scholar] [CrossRef]
- Krämer, O.H.; Schneider, G. Single-Cell Profiling Guided Combination Therapy of c-Fos and Histone Deacetylase Inhibitors in Diffuse Large B-Cell Lymphoma. Clin. Transl. Med. 2022, 12, e858. [Google Scholar] [CrossRef]
- Zrimšek, M.; Kuchaříková, H.; Draganić, K.; Dobrovolná, P.; Heiss Spornberger, V.; Winkelmayer, L.; Hassler, M.R.; Lochmanová, G.; Zdráhal, Z.; Egger, G. Quantitative Acetylomics Uncover Acetylation-Mediated Pathway Changes Following Histone Deacetylase Inhibition in Anaplastic Large Cell Lymphoma. Cells 2022, 11, 2380. [Google Scholar] [CrossRef]
- Fujiwara-Igarashi, A.; Tomiyasu, H.; Igarashi, H.; Yu, Y.; Gogo-Koshino, Y.; Ohno, K.; Tsujimoto, H. Regulation of P16 Gene Expression by Histone H3 Acetylation in Canine Lymphoid Tumor Cell Lines. Jpn. J. Vet. Res. 2016, 64, 257–263. [Google Scholar]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA Methyltransferases Dnmt3a and Dnmt3b Are Essential for de Novo Methylation and Mammalian Development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Szyf, M. Multiple Isoforms of DNA Methyltransferase Are Encoded by the Vertebrate Cytosine DNA Methyltransferase Gene. J. Biol. Chem. 1998, 273, 22869–22872. [Google Scholar] [CrossRef] [Green Version]
- Robertson, K.D.; Uzvolgyi, E.; Liang, G.; Talmadge, C.; Sumegi, J.; Gonzales, F.A.; Jones, P.A. The Human DNA Methyltransferases (DNMTs) 1, 3a and 3b: Coordinate MRNA Expression in Normal Tissues and Overexpression in Tumors. Nucleic Acids Res. 1999, 27, 2291–2298. [Google Scholar] [CrossRef] [Green Version]
- Llobat, L.; Gourbault, O. Role of MicroRNAs in Human Osteosarcoma: Future Perspectives. Biomedicines 2021, 9, 463. [Google Scholar] [CrossRef]
- Papanicolau-Sengos, A.; Aldape, K. DNA Methylation Profiling: An Emerging Paradigm for Cancer Diagnosis. Annu. Rev. Pathol. 2022, 17, 295–321. [Google Scholar] [CrossRef]
- Pelham, J.T.; Irwin, P.J.; Kay, P.H. Genomic Hypomethylation in Neoplastic Cells from Dogs with Malignant Lymphoproliferative Disorders. Res. Vet. Sci. 2003, 74, 101–104. [Google Scholar] [CrossRef]
- Morimoto, C.Y.; Tedardi, M.V.; da Fonseca, I.I.M.; Kimura, K.C.; Sanches, D.S.; Epiphanio, T.F.; de Francisco Strefezzi, R.; Dagli, M.L.Z. Evaluation of the Global DNA Methylation in Canine Mast Cell Tumour Samples by Immunostaining of 5-Methyl Cytosine. Vet. Comp. Oncol. 2017, 15, 1014–1018. [Google Scholar] [CrossRef]
- Herrera, C.L.; Kim, D.Y.; Kumar, S.R.; Bryan, J.N. Peroxisome Proliferator Activated Receptor γ Protein Expression Is Asymmetrically Distributed in Primary Lung Tumor and Metastatic to Lung Osteosarcoma Samples and Does Not Correlate with Gene Methylation. BMC Vet. Res. 2015, 11, 230. [Google Scholar] [CrossRef] [Green Version]
- Bryan, J.N.; Jabbes, M.; Berent, L.M.; Arthur, G.L.; Taylor, K.H.; Rissetto, K.C.; Henry, C.J.; Rahmatpanah, F.; Rankin, W.V.; Villamil, J.A.; et al. Hypermethylation of the DLC1 CpG Island Does Not Alter Gene Expression in Canine Lymphoma. BMC Genet. 2009, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, S.; Mori, T.; Nakagawa, T.; Itamoto, K.; Haraguchi, T.; Mizuno, T. DNA Methylation Contributes toward Silencing of Antioncogenic MicroRNA-203 in Human and Canine Melanoma Cells. Melanoma Res. 2015, 25, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Bronzini, I.; Aresu, L.; Paganin, M.; Marchioretto, L.; Comazzi, S.; Cian, F.; Riondato, F.; Marconato, L.; Martini, V.; Te Kronnie, G. DNA Methylation and Targeted Sequencing of Methyltransferases Family Genes in Canine Acute Myeloid Leukaemia, Modelling Human Myeloid Leukaemia. Vet. Comp. Oncol. 2017, 15, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Nam, A.-R.; Lee, K.-H.; Hwang, H.-J.; Schabort, J.J.; An, J.-H.; Won, S.-H.; Cho, J.-Y. Alternative Methylation of Intron Motifs Is Associated with Cancer-Related Gene Expression in Both Canine Mammary Tumor and Human Breast Cancer. Clin. Epigenet. 2020, 12, 110. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Yamazaki, J.; Jelinek, J.; Ishizaki, T.; Kagawa, Y.; Yokoyama, N.; Nagata, N.; Sasaki, N.; Takiguchi, M. Genome-Wide DNA Methylation Analysis in Canine Gastrointestinal Lymphoma. J. Vet. Med. Sci. 2020, 82, 632–638. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-H.; Tomiyasu, H.; Lee, J.-J.; Tung, C.-W.; Liao, C.-H.; Chuang, C.-H.; Huang, L.-Y.; Liao, K.-W.; Chou, C.-H.; Liao, A.T.C.; et al. Genome-Wide DNA Methylation Analysis Using MethylCap-Seq in Canine High-Grade B-Cell Lymphoma. J. Leukoc. Biol. 2021, 109, 1089–1103. [Google Scholar] [CrossRef]
- Ferraresso, S.; Aricò, A.; Sanavia, T.; Da Ros, S.; Milan, M.; Cascione, L.; Comazzi, S.; Martini, V.; Giantin, M.; Di Camillo, B.; et al. DNA Methylation Profiling Reveals Common Signatures of Tumorigenesis and Defines Epigenetic Prognostic Subtypes of Canine Diffuse Large B-Cell Lymphoma. Sci. Rep. 2017, 7, 11591. [Google Scholar] [CrossRef] [Green Version]
- Epiphanio, T.M.F.; de Azevedo Fernandes, N.C.C.; de Oliveira, T.F.; Lopes, P.A.; Réssio, R.A.; Gonçalves, S.; Scattone, N.V.; Tedardi, M.V.; Kulikowski, L.D.; Damasceno, J.; et al. Global DNA Methylation of Peripheral Blood Leukocytes from Dogs Bearing Multicentric Non-Hodgkin Lymphomas and Healthy Dogs: A Comparative Study. PLoS ONE 2019, 14, e0211898. [Google Scholar] [CrossRef] [Green Version]
- Chengxiao, Z.; Ze, Y. Biological Function and Molecular Mechanism of Twist2. Yi Chuan 2015, 37, 17–24. [Google Scholar] [CrossRef]
- Yamazaki, J.; Jelinek, J.; Hisamoto, S.; Tsukamoto, A.; Inaba, M. Dynamic Changes in DNA Methylation Patterns in Canine Lymphoma Cell Lines Demonstrated by Genome-Wide Quantitative DNA Methylation Analysis. Vet. J. 2018, 231, 48–54. [Google Scholar] [CrossRef]
- Ferraresso, S.; Bresolin, S.; Aricò, A.; Comazzi, S.; Gelain, M.E.; Riondato, F.; Bargelloni, L.; Marconato, L.; te Kronnie, G.; Aresu, L. Epigenetic Silencing of TFPI-2 in Canine Diffuse Large B-Cell Lymphoma. PLoS ONE 2014, 9, e92707. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Mochizuki, H.; Goto-Koshino, Y.; Fujiwara-Igarashi, A.; Takahashi, M.; Ohno, K.; Tsujimoto, H. Prognostic Significance of Hypermethylation of Death-Associated Protein Kinase (DAPK) Gene CpG Island in Dogs with High-Grade B-Cell Lymphoma. Vet. Comp. Oncol. 2018, 16, 409–415. [Google Scholar] [CrossRef]
- Liggett, W.H.; Sidransky, D. Role of the P16 Tumor Suppressor Gene in Cancer. J. Clin. Oncol. 1998, 16, 1197–1206. [Google Scholar] [CrossRef]
- Fujiwara-Igarashi, A.; Goto-Koshino, Y.; Mochizuki, H.; Sato, M.; Fujino, Y.; Ohno, K.; Tsujimoto, H. Inhibition of P16 Tumor Suppressor Gene Expression via Promoter Hypermethylation in Canine Lymphoid Tumor Cells. Res. Vet. Sci. 2014, 97, 60–63. [Google Scholar] [CrossRef]
- Fujiwara-Igarashi, A.; Goto-Koshino, Y.; Sato, M.; Maeda, S.; Igarashi, H.; Takahashi, M.; Fujino, Y.; Ohno, K.; Tsujimoto, H. Prognostic Significance of the Expression Levels of the P16, P15, and P14 Genes in Dogs with High-Grade Lymphoma. Vet. J. 2014, 199, 236–244. [Google Scholar] [CrossRef]
- Maylina, L.; Kambayashi, S.; Baba, K.; Okuda, M. Simultaneous Analysis of the P16 Gene and Protein in Canine Lymphoma Cells and Their Correlation with PRb Phosphorylation. Vet. Sci. 2022, 9, 393. [Google Scholar] [CrossRef]
- Fosmire, S.P.; Thomas, R.; Jubala, C.M.; Wojcieszyn, J.W.; Valli, V.E.O.; Getzy, D.M.; Smith, T.L.; Gardner, L.A.; Ritt, M.G.; Bell, J.S.; et al. Inactivation of the P16 Cyclin-Dependent Kinase Inhibitor in High-Grade Canine Non-Hodgkin’s T-Cell Lymphoma. Vet. Pathol. 2007, 44, 467–478. [Google Scholar] [CrossRef]
- Sato, M.; Mochizuki, H.; Goto-Koshino, Y.; Fujiwara-Igarashi, A.; Takahashi, M.; Fujino, Y.; Ohno, K.; Tsujimoto, H. Hypermethylation of the Death-Associated Protein Kinase CpG Island in Canine B-Cell Lymphoid Tumors. Vet. Immunol. Immunopathol. 2014, 161, 222–231. [Google Scholar] [CrossRef]
- Aresu, L. Canine Lymphoma, More Than a Morphological Diagnosis: What We Have Learned about Diffuse Large B-Cell Lymphoma. Front. Vet. Sci. 2016, 3, 77. [Google Scholar] [CrossRef] [Green Version]
- Zorzan, E.; Elgendy, R.; Guerra, G.; Da Ros, S.; Gelain, M.E.; Bonsembiante, F.; Garaffo, G.; Vitale, N.; Piva, R.; Marconato, L.; et al. Hypermethylation-Mediated Silencing of CIDEA, MAL and PCDH17 Tumour Suppressor Genes in Canine DLBCL: From Multi-Omics Analyses to Mechanistic Studies. Int. J. Mol. Sci. 2022, 23, 4021. [Google Scholar] [CrossRef]
- Chu, S.; Avery, A.; Yoshimoto, J.; Bryan, J.N. Genome Wide Exploration of the Methylome in Aggressive B-Cell Lymphoma in Golden Retrievers Reveals a Conserved Hypermethylome. Epigenetics 2022, 17, 2022–2038. [Google Scholar] [CrossRef]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of Small RNAs in Animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Doench, J.G.; Sharp, P.A. Specificity of MicroRNA Target Selection in Translational Repression. Genes Dev. 2004, 18, 504–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morlando, M.; Ballarino, M.; Gromak, N.; Pagano, F.; Bozzoni, I.; Proudfoot, N.J. Primary MicroRNA Transcripts Are Processed Co-Transcriptionally. Nat. Struct. Mol. Biol. 2008, 15, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Bohnsack, M.T.; Czaplinski, K.; Gorlich, D. Exportin 5 Is a RanGTP-Dependent DsRNA-Binding Protein That Mediates Nuclear Export of Pre-MiRNAs. RNA 2004, 10, 185–191. [Google Scholar] [CrossRef] [Green Version]
- Lund, E.; Güttinger, S.; Calado, A.; Dahlberg, J.E.; Kutay, U. Nuclear Export of MicroRNA Precursors. Science 2004, 303, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Yi, R.; Qin, Y.; Macara, I.G.; Cullen, B.R. Exportin-5 Mediates the Nuclear Export of Pre-MicroRNAs and Short Hairpin RNAs. Genes Dev. 2003, 17, 3011–3016. [Google Scholar] [CrossRef] [Green Version]
- Hutvágner, G.; McLachlan, J.; Pasquinelli, A.E.; Bálint, E.; Tuschl, T.; Zamore, P.D. A Cellular Function for the RNA-Interference Enzyme Dicer in the Maturation of the Let-7 Small Temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Ketting, R.F.; Fischer, S.E.; Bernstein, E.; Sijen, T.; Hannon, G.J.; Plasterk, R.H. Dicer Functions in RNA Interference and in Synthesis of Small RNA Involved in Developmental Timing in C. Elegans. Genes Dev. 2001, 15, 2654–2659. [Google Scholar] [CrossRef] [Green Version]
- Altuvia, Y.; Landgraf, P.; Lithwick, G.; Elefant, N.; Pfeffer, S.; Aravin, A.; Brownstein, M.J.; Tuschl, T.; Margalit, H. Clustering and Conservation Patterns of Human MicroRNAs. Nucleic Acids Res. 2005, 33, 2697–2706. [Google Scholar] [CrossRef] [Green Version]
- Gourbault, O.; Llobat, L. MicroRNAs as Biomarkers in Canine Osteosarcoma: A New Future? Vet. Sci. 2020, 7, 146. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Heneghan, H.M.; Miller, N.; Kerin, M.J. MiRNAs as Biomarkers and Therapeutic Targets in Cancer. Curr. Opin. Pharmacol. 2010, 10, 543–550. [Google Scholar] [CrossRef]
- Macfarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [Green Version]
- Conkrite, K.; Sundby, M.; Mukai, S.; Thomson, J.M.; Mu, D.; Hammond, S.M.; MacPherson, D. MiR-17∼92 Cooperates with RB Pathway Mutations to Promote Retinoblastoma. Genes Dev. 2011, 25, 1734–1745. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Wang, P.; Wang, Q.; Wang, B.; Mu, J.; Zhuang, X.; Zhang, L.; Yan, J.; Miller, D.; Zhang, H.-G. Quantitatively Controlling Expression of MiR-17∼92 Determines Colon Tumor Progression in a Mouse Tumor Model. Am. J. Pathol. 2014, 184, 1355–1368. [Google Scholar] [CrossRef] [Green Version]
- Hong, L.; Lai, M.; Chen, M.; Xie, C.; Liao, R.; Kang, Y.J.; Xiao, C.; Hu, W.-Y.; Han, J.; Sun, P. The MiR-17-92 Cluster of MicroRNAs Confers Tumorigenicity by Inhibiting Oncogene-Induced Senescence. Cancer Res. 2010, 70, 8547–8557. [Google Scholar] [CrossRef] [Green Version]
- Brockway, S.; Zeleznik-Le, N.J. WEE1 Is a Validated Target of the MicroRNA MiR-17-92 Cluster in Leukemia. Cancer Genet. 2015, 208, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Urtasun, R.; Elizalde, M.; Azkona, M.; Latasa, M.U.; García-Irigoyen, O.; Uriarte, I.; Fernández-Barrena, M.G.; Vicent, S.; Alonso, M.M.; Muntané, J.; et al. Splicing Regulator SLU7 Preserves Survival of Hepatocellular Carcinoma Cells and Other Solid Tumors via Oncogenic MiR-17-92 Cluster Expression. Oncogene 2016, 35, 4719–4729. [Google Scholar] [CrossRef]
- Smith, A.L.; Iwanaga, R.; Drasin, D.J.; Micalizzi, D.S.; Vartuli, R.L.; Tan, A.-C.; Ford, H.L. The MiR-106b-25 Cluster Targets Smad7, Activates TGF-β Signaling, and Induces EMT and Tumor Initiating Cell Characteristics Downstream of Six1 in Human Breast Cancer. Oncogene 2012, 31, 5162–5171. [Google Scholar] [CrossRef] [Green Version]
- Savita, U.; Karunagaran, D. MicroRNA-106b-25 Cluster Targets β-TRCP2, Increases the Expression of Snail and Enhances Cell Migration and Invasion in H1299 (Non Small Cell Lung Cancer) Cells. Biochem. Biophys. Res. Commun. 2013, 434, 841–847. [Google Scholar] [CrossRef]
- Xiao, C.; Calado, D.P.; Galler, G.; Thai, T.-H.; Patterson, H.C.; Wang, J.; Rajewsky, N.; Bender, T.P.; Rajewsky, K. MiR-150 Controls B-Cell Differentiation by Targeting the Transcription Factor c-Myb. Cell 2016, 165, 1027. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-D.; Li, X.-M.; Gu, J.-W.; Sun, X.-C. MiR-155 Regulates Lymphoma Cell Proliferation and Apoptosis through Targeting SOCS3/JAK-STAT3 Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7577. [Google Scholar] [CrossRef] [PubMed]
- Fuertes, T.; Ramiro, A.R.; de Yebenes, V.G. MiRNA-Based Therapies in B-Cell Non-Hodgkin Lymphoma. Trends Immunol. 2020, 41, 932–947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, M. Epigenetic Alterations and Advancement of Treatment in Peripheral T-Cell Lymphoma. Clin. Epigenet. 2020, 12, 169. [Google Scholar] [CrossRef]
- Mirna Therapeutics, Inc. A Multicenter Phase I Study of MRX34, MicroRNA MiR-RX34 Liposomal Injection; Mirna Therapeutics, Inc.: Austin, TX, USA, 2016. Available online: clinicaltrials.gov (accessed on 27 November 2022).
- miRagen Therapeutics, Inc. A Phase 1 Dose-Ranging Study to Investigate the Safety, Tolerability, and Pharmacokinetics of MRG-106 Following Local Intratumoral, Subcutaneous, and Intravenous Administration in Subjects with Various Lymphomas and Leukemias; miRagen Therapeutics, Inc.: Waltham, MA, USA, 2020. Available online: clinicaltrials.gov (accessed on 27 November 2022).
- Craig, K.K.L.; Wood, G.A.; Keller, S.M.; Mutsaers, A.J.; Wood, R.D. MicroRNA Profiling in Canine Multicentric Lymphoma. PLoS ONE 2019, 14, e0226357. [Google Scholar] [CrossRef] [Green Version]
- Joos, D.; Leipig-Rudolph, M.; Weber, K. Tumour-Specific MicroRNA Expression Pattern in Canine Intestinal T-Cell-Lymphomas. Vet. Comp. Oncol. 2020, 18, 502–508. [Google Scholar] [CrossRef] [Green Version]
- Olsen, E.A.; Kim, Y.H.; Kuzel, T.M.; Pacheco, T.R.; Foss, F.M.; Parker, S.; Frankel, S.R.; Chen, C.; Ricker, J.L.; Arduino, J.M.; et al. Phase IIb Multicenter Trial of Vorinostat in Patients with Persistent, Progressive, or Treatment Refractory Cutaneous T-Cell Lymphoma. J. Clin. Oncol. 2007, 25, 3109–3115. [Google Scholar] [CrossRef] [Green Version]
- Piekarz, R.L.; Frye, R.; Turner, M.; Wright, J.J.; Allen, S.L.; Kirschbaum, M.H.; Zain, J.; Prince, H.M.; Leonard, J.P.; Geskin, L.J.; et al. Phase II Multi-Institutional Trial of the Histone Deacetylase Inhibitor Romidepsin as Monotherapy for Patients with Cutaneous T-Cell Lymphoma. J. Clin. Oncol. 2009, 27, 5410–5417. [Google Scholar] [CrossRef] [Green Version]
- Foss, F.; Advani, R.; Duvic, M.; Hymes, K.B.; Intragumtornchai, T.; Lekhakula, A.; Shpilberg, O.; Lerner, A.; Belt, R.J.; Jacobsen, E.D.; et al. A Phase II Trial of Belinostat (PXD101) in Patients with Relapsed or Refractory Peripheral or Cutaneous T-Cell Lymphoma. Br. J. Haematol. 2015, 168, 811–819. [Google Scholar] [CrossRef]
- EMA Farydak. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/farydak (accessed on 27 November 2022).
- Drug Approval Package. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2015/205353Orig1s000TOC.cfm (accessed on 27 November 2022).
- Panobinostat May Be Active in Select Patients With Refractory DLBCL—Cancer Therapy Advisor. Available online: https://www.cancertherapyadvisor.com/home/cancer-topics/lymphoma/panobinostat-may-be-active-in-select-patients-with-refractory-dlbcl/ (accessed on 27 November 2022).
- Batlevi, C.L.; Crump, M.; Andreadis, C.; Rizzieri, D.; Assouline, S.E.; Fox, S.; van der Jagt, R.H.C.; Copeland, A.; Potvin, D.; Chao, R.; et al. A Phase 2 Study of Mocetinostat, a Histone Deacetylase Inhibitor, in Relapsed or Refractory Lymphoma. Br. J. Haematol. 2017, 178, 434–441. [Google Scholar] [CrossRef] [Green Version]
- Dias, J.N.R.; Aguiar, S.I.; Pereira, D.M.; André, A.S.; Gano, L.; Correia, J.D.G.; Carrapiço, B.; Rütgen, B.; Malhó, R.; Peleteiro, C.; et al. The Histone Deacetylase Inhibitor Panobinostat Is a Potent Antitumor Agent in Canine Diffuse Large B-Cell Lymphoma. Oncotarget 2018, 9, 28586–28598. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, X.; Yuan, B.; Ren, L.; Zhang, T.; Lu, G. Subchronic Toxicities of HZ1006, a Hydroxamate-Based Histone Deacetylase Inhibitor, in Beagle Dogs and Sprague-Dawley Rats. Int. J. Environ. Res. Public Health 2016, 13, 1190. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Lan, F.; Matson, C.; Mulligan, P.; Whetstine, J.R.; Cole, P.A.; Casero, R.A.; Shi, Y. Histone Demethylation Mediated by the Nuclear Amine Oxidase Homolog LSD1. Cell 2004, 119, 941–953. [Google Scholar] [CrossRef] [Green Version]
- Metzger, J.; Nolte, A.; Uhde, A.-K.; Hewicker-Trautwein, M.; Distl, O. Whole Genome Sequencing Identifies Missense Mutation in MTBP in Shar-Pei Affected with Autoinflammatory Disease (SPAID). BMC Genom. 2017, 18, 348. [Google Scholar] [CrossRef] [Green Version]
- Wobser, M.; Weber, A.; Glunz, A.; Tauch, S.; Seitz, K.; Butelmann, T.; Hesbacher, S.; Goebeler, M.; Bartz, R.; Kohlhof, H.; et al. Elucidating the Mechanism of Action of Domatinostat (4SC-202) in Cutaneous T Cell Lymphoma Cells. J. Hematol. Oncol. 2019, 12, 30. [Google Scholar] [CrossRef] [Green Version]
- Hollebecque, A.; Salvagni, S.; Plummer, R.; Niccoli, P.; Capdevila, J.; Curigliano, G.; Moreno, V.; de Braud, F.; de Villambrosia, S.G.; Martin-Romano, P.; et al. Clinical Activity of CC-90011, an Oral, Potent, and Reversible LSD1 Inhibitor, in Advanced Malignancies. Cancer 2022, 128, 3185–3195. [Google Scholar] [CrossRef]
- Hollebecque, A.; Salvagni, S.; Plummer, R.; Isambert, N.; Niccoli, P.; Capdevila, J.; Curigliano, G.; Moreno, V.; Martin-Romano, P.; Baudin, E.; et al. Phase I Study of Lysine-Specific Demethylase 1 Inhibitor, CC-90011, in Patients with Advanced Solid Tumors and Relapsed/Refractory Non-Hodgkin Lymphoma. Clin. Cancer Res. 2021, 27, 438–446. [Google Scholar] [CrossRef]
- Roboz, G.J.; Yee, K.; Verma, A.; Borthakur, G.; de la Fuente Burguera, A.; Sanz, G.; Mohammad, H.P.; Kruger, R.G.; Karpinich, N.O.; Ferron-Brady, G.; et al. Phase I Trials of the Lysine-Specific Demethylase 1 Inhibitor, GSK2879552, as Mono- and Combination-Therapy in Relapsed/Refractory Acute Myeloid Leukemia or High-Risk Myelodysplastic Syndromes. Leuk. Lymphoma 2022, 63, 463–467. [Google Scholar] [CrossRef]
- Liu, H.; Wei, J.; Sang, N.; Zhong, X.; Zhou, X.; Yang, X.; Zhang, J.; Zuo, Z.; Zhou, Y.; Yang, S.; et al. The Novel LSD1 Inhibitor ZY0511 Suppresses Diffuse Large B-Cell Lymphoma Proliferation by Inducing Apoptosis and Autophagy. Med. Oncol. 2021, 38, 124. [Google Scholar] [CrossRef]
- Mathur, R.; Sehgal, L.; Havranek, O.; Köhrer, S.; Khashab, T.; Jain, N.; Burger, J.A.; Neelapu, S.S.; Davis, R.E.; Samaniego, F. Inhibition of Demethylase KDM6B Sensitizes Diffuse Large B-Cell Lymphoma to Chemotherapeutic Drugs. Haematologica 2017, 102, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Franci, G.; Sarno, F.; Nebbioso, A.; Altucci, L. Identification and Characterization of PKF118-310 as a KDM4A Inhibitor. Epigenetics 2017, 12, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shen, L.; Stupack, D.G.; Bai, N.; Xun, J.; Ren, G.; Han, J.; Li, L.; Luo, Y.; Xiang, R.; et al. JMJD3 Promotes Survival of Diffuse Large B-Cell Lymphoma Subtypes via Distinct Mechanisms. Oncotarget 2016, 7, 29387–29399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flesner, B.K.; Kumar, S.R.; Bryan, J.N. 6-Thioguanine and Zebularine down-Regulate DNMT1 and Globally Demethylate Canine Malignant Lymphoid Cells. BMC Vet. Res. 2014, 10, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, S.; Yamazaki, J.; Iwahana, M.; Tsukamoto, A. Olsalazine Inhibits Cell Proliferation and DNA Methylation in Canine Lymphoid Tumor Cell Lines. Pol. J. Vet. Sci. 2021, 24, 515–523. [Google Scholar] [CrossRef]
- Da Ros, S.; Aresu, L.; Ferraresso, S.; Zorzan, E.; Gaudio, E.; Bertoni, F.; Dacasto, M.; Giantin, M. Validation of Epigenetic Mechanisms Regulating Gene Expression in Canine B-Cell Lymphoma: An in Vitro and in Vivo Approach. PLoS ONE 2018, 13, e0208709. [Google Scholar] [CrossRef]
- Sloan, S.L.; Renaldo, K.A.; Long, M.; Chung, J.-H.; Courtney, L.E.; Shilo, K.; Youssef, Y.; Schlotter, S.; Brown, F.; Klamer, B.G.; et al. Validation of Protein Arginine Methyltransferase 5 (PRMT5) as a Candidate Therapeutic Target in the Spontaneous Canine Model of Non-Hodgkin Lymphoma. PLoS ONE 2021, 16, e0250839. [Google Scholar] [CrossRef]
- Tomiyasu, H.; Fujiwara-Igarashi, A.; Goto-Koshino, Y.; Fujino, Y.; Ohno, K.; Tsujimoto, H. Evaluation of DNA Methylation Profiles of the CpG Island of the ABCB1 Gene in Dogs with Lymphoma. Am. J. Vet. Res. 2014, 75, 835–841. [Google Scholar] [CrossRef]
- Kambayashi, S.; Minami, K.; Ogawa, Y.; Hamaji, T.; Hwang, C.C.; Igase, M.; Hiraoka, H.; Miyama, T.S.; Noguchi, S.; Baba, K.; et al. Expression of O(6)-Methylguanine-DNA Methyltransferase Causes Lomustine Resistance in Canine Lymphoma Cells. Can. J. Vet. Res. 2015, 79, 201–209. [Google Scholar]
- Parachini-Winter, C.; Bracha, S.; Ramsey, S.A.; Yang, L.; Ho, E.; Leeper, H.J.; Curran, K.M. Prospective Evaluation of the Lymph Node Proteome in Dogs with Multicentric Lymphoma Supplemented with Sulforaphane. J. Vet. Intern. Med. 2020, 34, 2036–2047. [Google Scholar] [CrossRef]
- Clarke, J.D.; Hsu, A.; Yu, Z.; Dashwood, R.H.; Ho, E. Differential Effects of Sulforaphane on Histone Deacetylases, Cell Cycle Arrest and Apoptosis in Normal Prostate Cells versus Hyperplastic and Cancerous Prostate Cells. Mol. Nutr. Food. Res. 2011, 55, 999–1009. [Google Scholar] [CrossRef] [Green Version]
- Suppipat, K.; Park, C.S.; Shen, Y.; Zhu, X.; Lacorazza, H.D. Sulforaphane Induces Cell Cycle Arrest and Apoptosis in Acute Lymphoblastic Leukemia Cells. PLoS ONE 2012, 7, e51251. [Google Scholar] [CrossRef] [Green Version]
- Jakubikova, J.; Cervi, D.; Ooi, M.; Kim, K.; Nahar, S.; Klippel, S.; Cholujova, D.; Leiba, M.; Daley, J.F.; Delmore, J.; et al. Anti-Tumor Activity and Signaling Events Triggered by the Isothiocyanates, Sulforaphane and Phenethyl Isothiocyanate, in Multiple Myeloma. Haematologica 2011, 96, 1170–1179. [Google Scholar] [CrossRef]
- Cui, H.; Hong, Q.; Wei, R.; Li, H.; Wan, C.; Chen, X.; Zhao, S.; Bu, H.; Zhang, B.; Yang, D.; et al. Design and Synthesis of HDAC Inhibitors to Enhance the Therapeutic Effect of Diffuse Large B-Cell Lymphoma by Improving Metabolic Stability and Pharmacokinetic Characteristics. Eur. J. Med. Chem. 2022, 229, 114049. [Google Scholar] [CrossRef]
- Sender, S.; Sultan, A.W.; Palmer, D.; Koczan, D.; Sekora, A.; Beck, J.; Schuetz, E.; Taher, L.; Brenig, B.; Fuellen, G.; et al. Evaluation of the Synergistic Potential of Simultaneous Pan- or Isoform-Specific BET and SYK Inhibition in B-Cell Lymphoma: An In Vitro Approach. Cancers 2022, 14, 4691. [Google Scholar] [CrossRef]
- Kong, W.; Sender, S.; Perez, S.V.; Sekora, A.; Ruetgen, B.; Junghanss, C.; Nolte, I.; Murua Escobar, H. Pan- and Isoform-Specific Inhibition of the Bromodomain and Extra-Terminal Proteins and Evaluation of Synergistic Potential With Entospletinib in Canine Lymphoma. Anticancer Res. 2020, 40, 3781–3792. [Google Scholar] [CrossRef]
- Hammond, S.M. An Overview of MicroRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Dutta, A. MicroRNAs in Cancer. Annu. Rev. Pathol. 2009, 4, 199–227. [Google Scholar] [CrossRef]
- Bryan, J.N. The Current State of Clinical Application of Serum Biomarkers for Canine Lymphoma. Front. Vet. Sci. 2016, 3, 87. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, N.O.; do Nascimento, N.C.; Lichti, N.I.; Kasinski, A.L.; Childress, M.O.; Santos, A.P.D. MicroRNA Biomarkers in Canine Diffuse Large B-Cell Lymphoma. Vet. Pathol. 2021, 58, 34–41. [Google Scholar] [CrossRef]
- Uhl, E.; Krimer, P.; Schliekelman, P.; Tompkins, S.M.; Suter, S. Identification of Altered MicroRNA Expression in Canine Lymphoid Cell Lines and Cases of B- and T-Cell Lymphomas. Genes Chromosomes Cancer 2011, 50, 950–967. [Google Scholar] [CrossRef]
- Albonico, F.; Mortarino, M.; Avallone, G.; Gioia, G.; Comazzi, S.; Roccabianca, P. The Expression Ratio of MiR-17-5p and MiR-155 Correlates with Grading in Canine Splenic Lymphoma. Vet. Immunol. Immunopathol. 2013, 155, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Mortarino, M.; Gioia, G.; Gelain, M.E.; Albonico, F.; Roccabianca, P.; Ferri, E.; Comazzi, S. Identification of Suitable Endogenous Controls and Differentially Expressed MicroRNAs in Canine Fresh-Frozen and FFPE Lymphoma Samples. Leuk. Res. 2010, 34, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara-Igarashi, A.; Igarashi, H.; Mizutani, N.; Goto-Koshino, Y.; Takahashi, M.; Ohno, K.; Tsujimoto, H. Expression Profile of Circulating Serum MicroRNAs in Dogs with Lymphoma. Vet. J. 2015, 205, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ding, L.; Hu, Q.; Xia, J.; Sun, J.; Wang, X.; Xiong, H.; Gurbani, D.; Li, L.; Liu, Y.; et al. MicroRNA-218 Functions as a Tumor Suppressor in Lung Cancer by Targeting IL-6/STAT3 and Negatively Correlates with Poor Prognosis. Mol. Cancer 2017, 16, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagawa, H.; Ikeda, S.; Sawada, K. Role of MicroRNA in the Pathogenesis of Malignant Lymphoma. Cancer Sci. 2013, 104, 801–809. [Google Scholar] [CrossRef]
- Chen, A.-H.; Qin, Y.-E.; Tang, W.-F.; Tao, J.; Song, H.-M.; Zuo, M. MiR-34a and MiR-206 Act as Novel Prognostic and Therapy Biomarkers in Cervical Cancer. Cancer Cell Int. 2017, 17, 63. [Google Scholar] [CrossRef] [Green Version]
- Nie, K.; Zhang, T.; Allawi, H.; Gomez, M.; Liu, Y.; Chadburn, A.; Wang, Y.L.; Knowles, D.M.; Tam, W. Epigenetic Down-Regulation of the Tumor Suppressor Gene PRDM1/Blimp-1 in Diffuse Large B-Cell Lymphomas: A Potential Role of the MicroRNA Let-7. Am. J. Pathol. 2010, 177, 1470–1479. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J. MiR-223 Suppresses Proliferation and Promotes Apoptosis of Diffuse Large B-Cell Lymphoma Cells Through Lmo2 and MAPK Signaling Pathway. J. BUON 2021, 26, 580–586. [Google Scholar]
- Wu, T.; Hu, H.; Zhang, T.; Jiang, L.; Li, X.; Liu, S.; Zheng, C.; Yan, G.; Chen, W.; Ning, Y.; et al. MiR-25 Promotes Cell Proliferation, Migration, and Invasion of Non-Small-Cell Lung Cancer by Targeting the LATS2/YAP Signaling Pathway. Oxid. Med. Cell Longev. 2019, 2019, 9719723. [Google Scholar] [CrossRef] [Green Version]
- Elliott, E.K.; Hopkins, L.N.; Hensen, R.; Sutherland, H.G.; Haupt, L.M.; Griffiths, L.R. Epigenetic Regulation of MiR-92a and TET2 and Their Association in Non-Hodgkin Lymphoma. Front. Genet. 2021, 12, 768913. [Google Scholar] [CrossRef]
- Mazzoccoli, L.; Robaina, M.C.; Bacchi, C.E.; Soares Lima, S.C.; Klumb, C.E. MiR-29 Promoter and Enhancer Methylation Identified by Pyrosequencing in Burkitt Lymhoma Cells: Interplay between MYC and MiR-29 Regulation. Oncol. Rep. 2019, 42, 775–784. [Google Scholar] [CrossRef]
- Thompson, M.A.; Edmonds, M.D.; Liang, S.; McClintock-Treep, S.; Wang, X.; Li, S.; Eischen, C.M. MiR-31 and MiR-17-5p Levels Change during Transformation of Follicular Lymphoma. Hum. Pathol. 2016, 50, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Gao, P.; Tchernyshyov, I.; Chang, T.-C.; Lee, Y.-S.; Kita, K.; Ochi, T.; Zeller, K.I.; De Marzo, A.M.; Van Eyk, J.E.; Mendell, J.T.; et al. C-Myc Suppression of MiR-23a/b Enhances Mitochondrial Glutaminase Expression and Glutamine Metabolism. Nature 2009, 458, 762–765. [Google Scholar] [CrossRef]
- Niu, F.; Kazimierska, M.; Nolte, I.M.; Terpstra, M.M.; de Jong, D.; Koerts, J.; van der Sluis, T.; Rutgers, B.; O’Connell, R.M.; Kok, K.; et al. The MiR-26b-5p/KPNA2 Axis Is an Important Regulator of Burkitt Lymphoma Cell Growth. Cancers 2020, 12, 1464. [Google Scholar] [CrossRef]
- Si, X.; Zhang, X.; Hao, X.; Li, Y.; Chen, Z.; Ding, Y.; Shi, H.; Bai, J.; Gao, Y.; Cheng, T.; et al. Upregulation of MiR-99a Is Associated with Poor Prognosis of Acute Myeloid Leukemia and Promotes Myeloid Leukemia Cell Expansion. Oncotarget 2016, 7, 78095–78109. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-W.; Ramasamy, K.; Bouamar, H.; Lin, A.-P.; Jiang, D.; Aguiar, R.C.T. MicroRNAs MiR-125a and MiR-125b Constitutively Activate the NF-ΚB Pathway by Targeting the Tumor Necrosis Factor Alpha-Induced Protein 3 (TNFAIP3, A20). Proc. Natl. Acad. Sci. USA 2012, 109, 7865–7870. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Cheng, Y.; Dong, S.; Du, J.; Gao, F.; Sun, D.; Cui, J.; Ni, J.; Cai, J. Down Regulation of MiR-143 Promotes Radiation—Induced Thymic Lymphoma by Targeting B7H1. Toxicol. Lett. 2017, 280, 116–124. [Google Scholar] [CrossRef]
- Lone, W.; Bouska, A.; Sharma, S.; Amador, C.; Saumyaranjan, M.; Herek, T.A.; Heavican, T.B.; Yu, J.; Lim, S.T.; Ong, C.K.; et al. Genome-Wide MiRNA Expression Profiling of Molecular Subgroups of Peripheral T-Cell Lymphoma. Clin. Cancer Res. 2021, 27, 6039–6053. [Google Scholar] [CrossRef]
- Ramadan, E.S.; Kubesy, A.A.; Baraka, T.A.; Torad, F.A.; Salem, S.I.; Salem, N.Y. Expression of Blood Hepatocyte-Derived MicroRNA-122 in Canine Multicentric Lymphoma with Hepatic Involvement. Vet. Res. Commun. 2019, 43, 231–238. [Google Scholar] [CrossRef]
- Kulka, M.; Brennan, K.; Mc Gee, M. Investigation of Canine Extracellular Vesicles in Diffuse Large B-Cell Lymphomas. PLoS ONE 2022, 17, e0274261. [Google Scholar] [CrossRef]
- Garnica, T.K.; Lesbon, J.C.C.; Ávila, A.C.F.C.M.; Rochetti, A.L.; Matiz, O.R.S.; Ribeiro, R.C.S.; Zoppa, A.; Nishiya, A.T.; Costa, M.T.; de Nardi, A.B.; et al. Liquid Biopsy Based on Small Extracellular Vesicles Predicts Chemotherapy Response of Canine Multicentric Lymphomas. Sci. Rep. 2020, 10, 20371. [Google Scholar] [CrossRef] [PubMed]
- Asada, H.; Tomiyasu, H.; Uchikai, T.; Ishihara, G.; Goto-Koshino, Y.; Ohno, K.; Tsujimoto, H. Comprehensive Analysis of MiRNA and Protein Profiles within Exosomes Derived from Canine Lymphoid Tumour Cell Lines. PLoS ONE 2019, 14, e0208567. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lymphoma Type | Clinical Signs | Observation | References |
|---|---|---|---|
| Multicentric | Generalised lymphadenomegaly. Involvement of the spleen, liver, and bone marrow. | The most predominant type of lymphoma (80–85%). | [6] |
| Gastrointestinal | Vomiting, diarrhoea, melena, and weight loss due to malabsorption and maldigestion of nutrients. It can present as an acute or chronic lymphoma. | The second most prevalent (10%). | [24] |
| Mediastinal | Coughing, breathing difficulty and pleural effusions. Polyuria and polydipsia can be seen due to hypercalcaemia (present in 40% of cases). | The third most common type of canine lymphoma. | [25] |
| Extranodal | Blindness, renal failure, seizures, bone fractures and respiratory disease. Organs typically affected are the eyes, kidneys, lungs, skin, and central nervous system; other areas that can be invaded include the mammary tissue, liver, bones and mouth. Cutaneous lymphoma, present as a skin rash with dry, red, and itchy bumps or solitary or generalised scaly lesions and can also affect the oral cavity, causing ulcers, lesions and nodules on the gums, lips and roof of the mouth. | The rarest form. Cutaneous lymphoma, the most common form of extranodal lymphoma, in early stages usually present as a skin rash with dry, red and itchy bumps or solitary or generalised scaly lesions. Sometimes may be mistaken for allergies or fungal infections. As the disease progresses, cutaneous lesions will become more severe, and large masses or tumours can develop. Cutaneous lymphoma can also affect the oral cavity causing ulcers, lesions and nodules on the gums, lips, and roof of the mouth. | [26] |
| miRNAs | Expression Profiles | Target Genes * | Result of Aberrant miRNA Expression | Possible Uses | References |
|---|---|---|---|---|---|
| miR-155 | Downregulated | HDAC4, PIK3R1, SMAD5, SHIP1 | Induces cell proliferation, activates oncogenic AKT signalling, enhances tumour aggressiveness | Potential therapeutic target | [148] |
| miR-17-5p | Upregulated | PTEN | Disease progression | Potential prognostic biomarker Potential therapeutic target | [148] |
| miR-181 | Downregulated | Bcl-2/TCL-1 | Activation of oncogenes | Potential prognosis biomarker Potential therapeutic target | [110] |
| miR-203 | Upregulated | ABL1 | Tumour cell proliferation | Potential therapeutic target | [110,147] |
| miR-218 | Upregulated | SLIT2/3 | Tumour cell proliferation | Potential therapeutic target | [151] |
| miR-19a | Upregulated | PTEN | Tumour cell proliferation | Potential prognostic biomarker Potential therapeutic target | [152] |
| miR-19b | Upregulated | PTEN | Tumour cell proliferation | Potential prognostic biomarker Potential therapeutic target | [152] |
| miR-34a | Upregulated | MET, SIRT1, CDK6, VEGF1 | Induced cell migration | Potential prognostic biomarker Potential therapeutic target | [153] |
| Let-7 family | Downregulated | PRDM1 ⁄Blimp-1 | Reduces terminal B-cell differentiation | Potential diagnosis biomarker Potential therapeutic target | [154] |
| miR-223 | Downregulated | LMO2 | Increase tumour progression | Potential prognosis biomarker | [155] |
| miR-25 | Downregulated | LATS2 | Tumour cell proliferation | Potential prognosis biomarker Potential therapeutic target | [156] |
| miR-92a | Downregulated | TET2 | Tumour cell proliferation Increase tumour progression | Potential diagnosis biomarker Potential prognosis biomarker | [157] |
| miR-423a | Downregulated | Unknown | Tumour cell proliferation | Potential diagnosis biomarker Potential prognosis biomarker | [150] |
| miR-29a/b/c | Upregulated | MYC, DNMT3B, MCL1, BIM, CDK6, AKT, TCL1 | Tumour B-cell proliferation | Potential diagnosis biomarker Potential therapeutic target | [158] |
| miR-31 | Upregulated | E2F2, PI3KC2A | Tumour B-cell proliferation | Potential diagnosis biomarker | [159] |
| miR-23a | Downregulated | MYC, MTSS1 | Tumour B-cell proliferation Tumour T-cell proliferation | Potential diagnosis biomarker | [160] |
| miR-26b | Downregulated | EZH2, COPS2, KPNA2, MRPL15, NOL12 | Tumour B-cell proliferation Tumour T-cell proliferation | Potential diagnosis biomarker | [161] |
| miR-99a | Downregulated | Unknown | Tumour B-cell proliferation | Potential diagnosis biomarker | [162] |
| miR-125a | Downregulated | TNFAIP3, NF1, Bcl-2 | Tumour B-cell proliferation | Potential diagnosis biomarker | [163] |
| miR-143 | Downregulated | B7H1, Bcl-2 | Tumour B-cell proliferation | Potential diagnosis biomarker | [164] |
| miR-145 | Downregulated | ROCK1, MYC | Tumour T-cell proliferation | Potential diagnosis biomarker | [165] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montaner-Angoiti, E.; Marín-García, P.J.; Llobat, L. Epigenetic Alterations in Canine Malignant Lymphoma: Future and Clinical Outcomes. Animals 2023, 13, 468. https://doi.org/10.3390/ani13030468
Montaner-Angoiti E, Marín-García PJ, Llobat L. Epigenetic Alterations in Canine Malignant Lymphoma: Future and Clinical Outcomes. Animals. 2023; 13(3):468. https://doi.org/10.3390/ani13030468
Chicago/Turabian StyleMontaner-Angoiti, Esperanza, Pablo Jesús Marín-García, and Lola Llobat. 2023. "Epigenetic Alterations in Canine Malignant Lymphoma: Future and Clinical Outcomes" Animals 13, no. 3: 468. https://doi.org/10.3390/ani13030468