
An Evaluation of Laminarin Additive in the Diets of Juvenile Largemouth Bass (Micropterus salmoides): Growth, Antioxidant Capacity, Immune Response and Intestinal Microbiota

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Design
2.3. Sampling
2.4. Flesh Composition and Antioxidant Capacity Analysis
2.5. Relative Gene Expression Analysis
2.6. Sequencing of Intestinal Microorganisms
2.7. Statistics Analysis
3. Results
3.1. Growth Performance
3.2. Flesh Composition
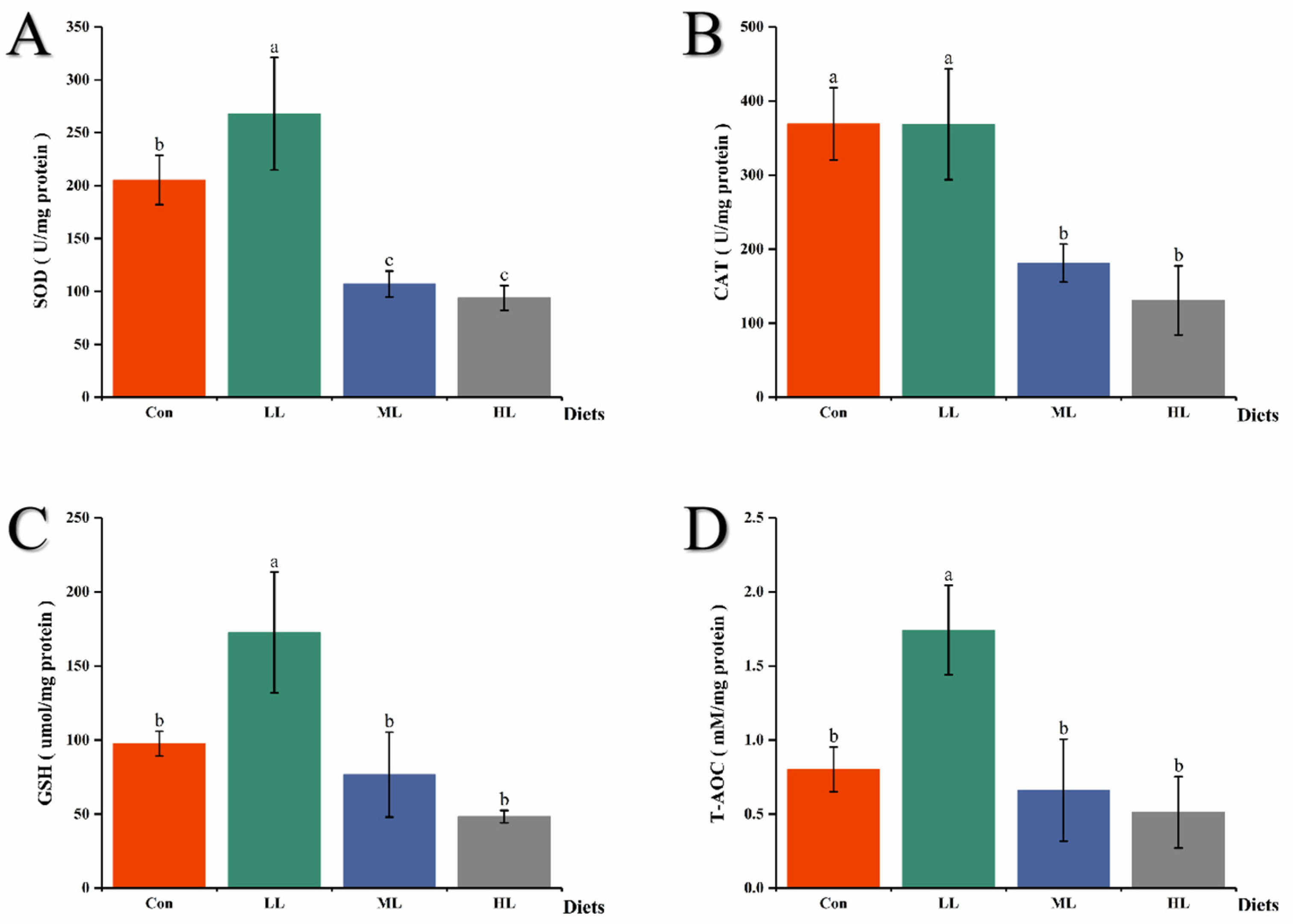
3.3. Antioxidant-related Enzyme Activities
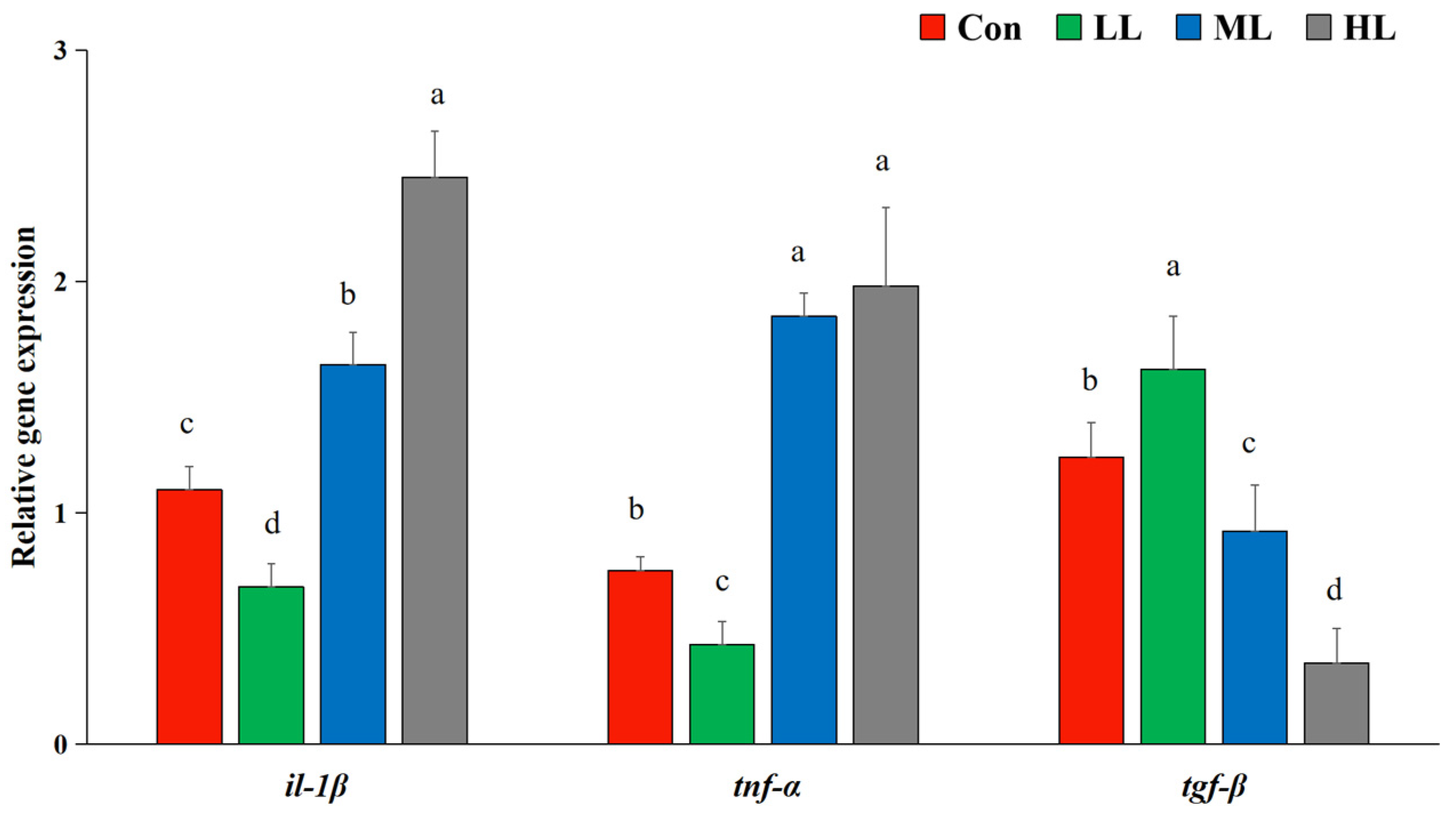
3.4. Relative Expression Levels of Immune Response Related Genes
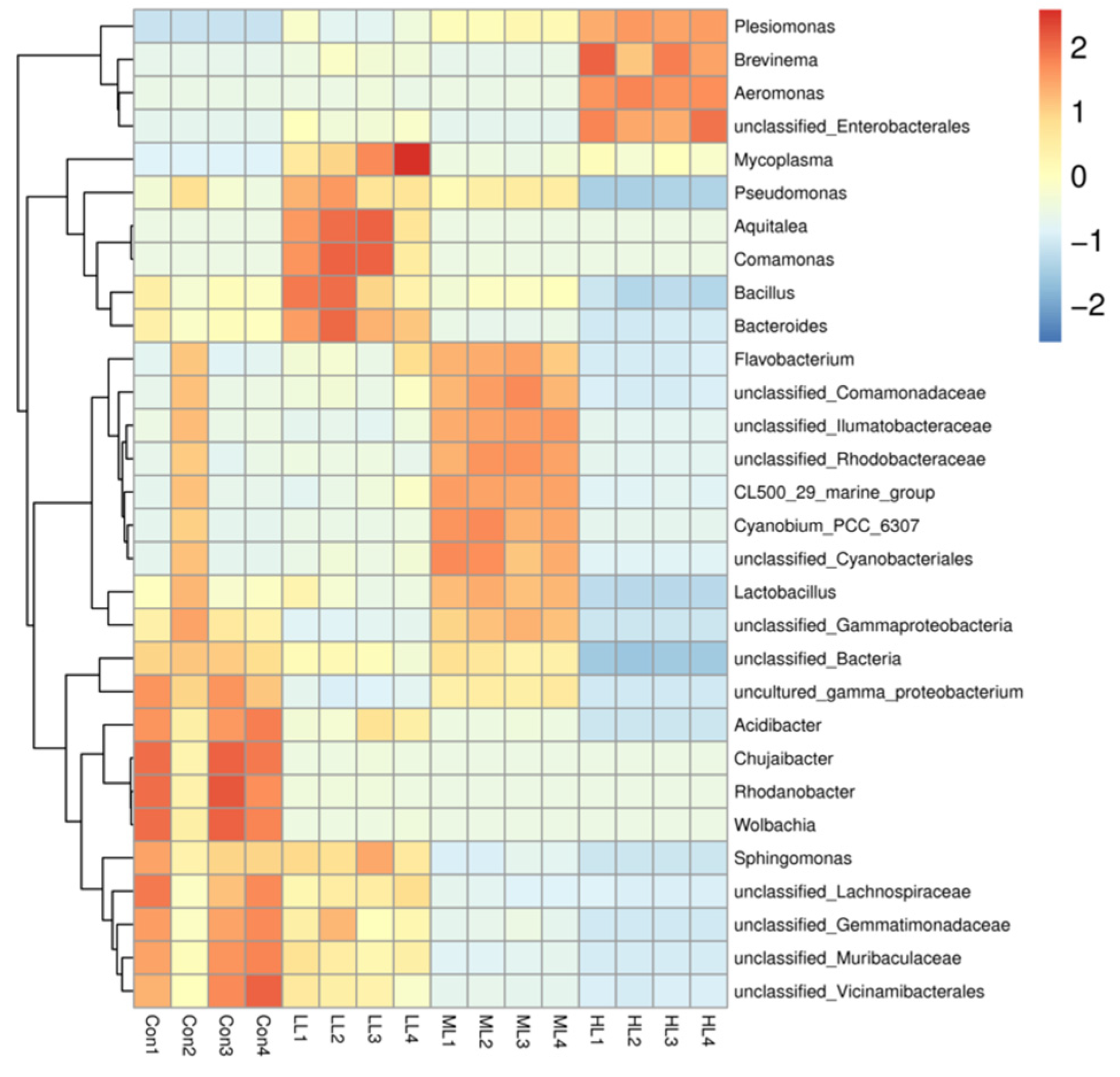
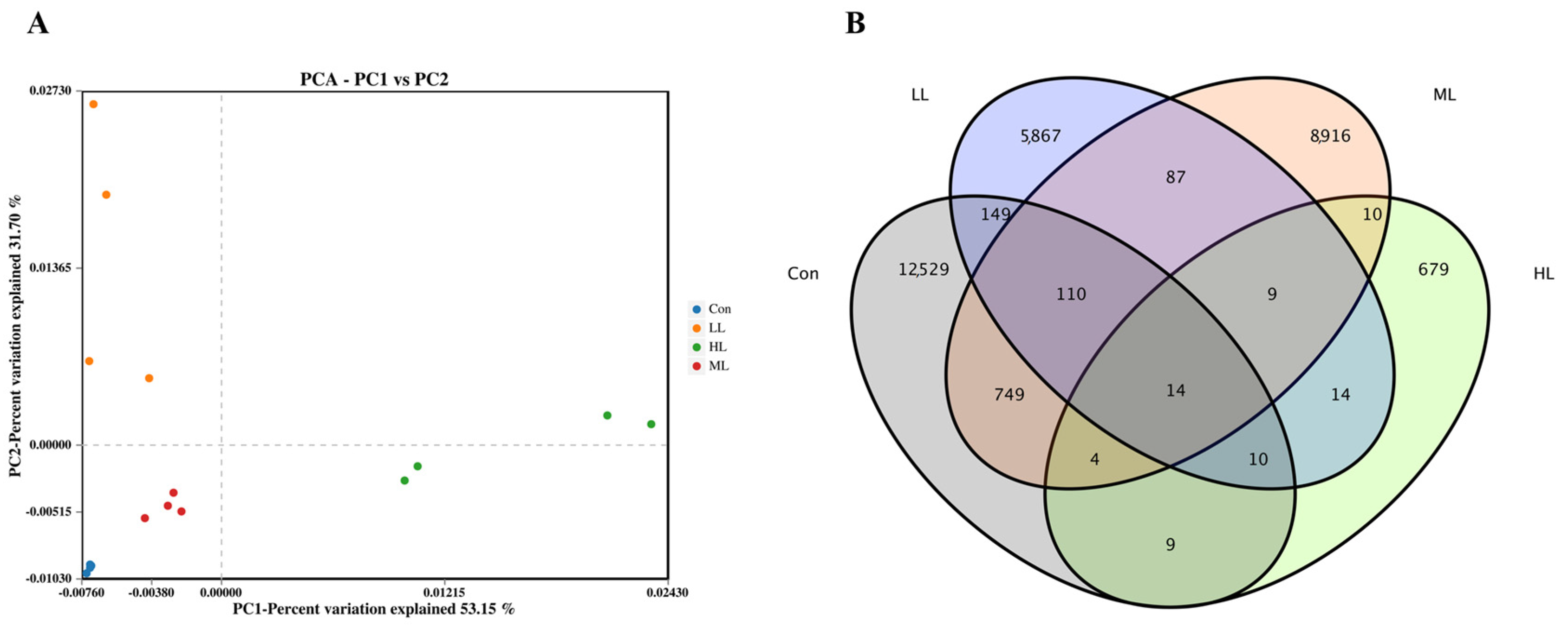
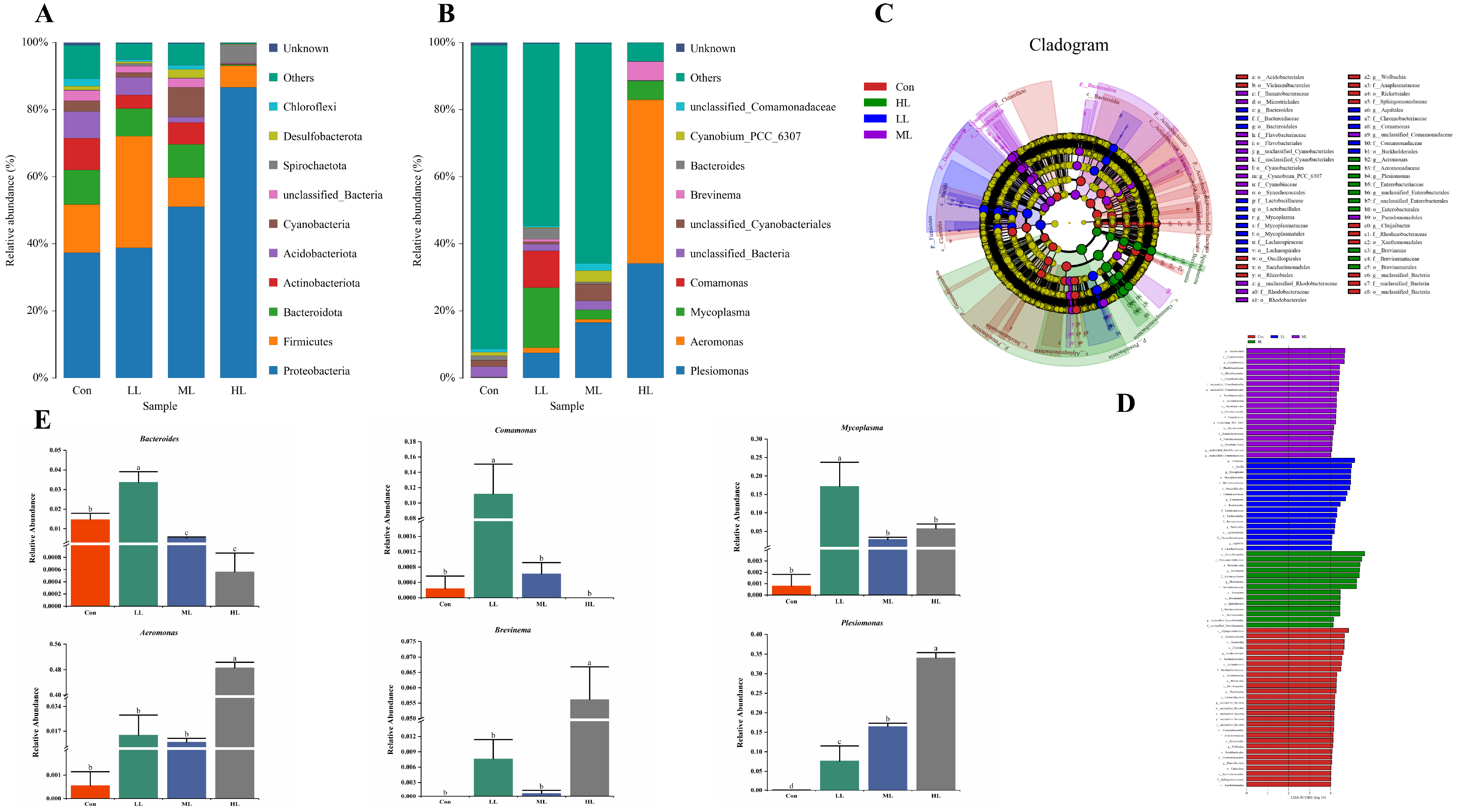
3.5. Intestinal Microbiota Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhong, Y.F.; Shi, C.M.; Zhou, Y.L.; Chen, Y.J.; Lin, S.M.; Tang, R.J. Optimum dietary fiber level could improve growth, plasma biochemical indexes and liver function of largemouth bass, Micropterus salmoides. Aquaculture 2020, 518, 734661. [Google Scholar] [CrossRef]
- Rossi, W., Jr.; Allen, K.M.; Habte-Tsion, H.-M.; Meesala, K.-M. Supplementation of glycine, prebiotic, and nucleotides in soybean mealbased diets for largemouth bass (Micropterus salmoides): Effects on production performance, whole-body nutrient composition and retention, and intestinal histopathology. Aquaculture 2021, 532, 736031. [Google Scholar] [CrossRef]
- Fei, H.; Cheng, Y.; Zhang, H.; Yu, X.; Yi, S.; Huang, M.; Yang, S. Effect of Autolyzed Yarrowia lipolytica on the Growth Performance, Antioxidant Capacity, Intestinal Histology, Microbiota, and Transcriptome Profile of Juvenile Largemouth Bass (Micropterus salmoides). Int. J. Mol. Sci. 2022, 23, 10780. [Google Scholar] [CrossRef]
- Khanjani, M.; Sharifinia, M.; Ghaedi, G. β-glucan as a promising food additive and immunostimulant in aquaculture industry. Ann. Anim. Sci. 2021, 22, 817–827. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Structural characterization of laminaran and galactofucan extracted from the brown seaweed Saccharina longicruris. Phytochemistry 2010, 71, 1586–1595. [Google Scholar] [CrossRef]
- Hadiuzzaman, M.; Moniruzzaman, M.; Shahjahan, M.; Bai, S.C.; Min, T.; Hossain, Z. β-Glucan: Mode of Action and Its Uses in Fish Immunomodulation. Front. Mar. Sci. 2022, 9, 905986. [Google Scholar] [CrossRef]
- Lin, J.B.; Liang, P.; Zhu, Q.G.; Qiu, M.L. Effect of Laminarin on Growth and Immunity of Pearl Gentian Grouper. Anim. Husb. Feed Sci. 2017, 9, 259–262. [Google Scholar]
- Schmitt, P.; Morales-Lange, B.; Bethke, J.; Mercado, L. Immunostimulatory effect of beta-glucans in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immun. 2013, 34, 1736–1737. [Google Scholar] [CrossRef]
- Yin, G.W.; Li, W.W.; Lin, Q.; Lin, X.; Lin, J.B. Dietary administration of laminarin improves the growth performance and immune responses in Epinephelus coioides. Fish Shellfish Immun. 2014, 41, 402–406. [Google Scholar] [CrossRef]
- Carballo, C.; Chronopoulou, E.G.; Letsiou, S.; Maya, C.; Labrou, N.E.; Infante, C.; Power, D.; Manchado, M. Antioxidant capacity and immunomodulatory effects of a chrysolaminarin-enriched extract in Senegalese sole. Fish Shellfish Immun. 2018, 82, 1–8. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, M.; Zheng, Y.; Chen, F.W.; Fu, L.L.; Zhong, L.; Chen, X.; Bian, W. Dietary laminarin administration to enhance the immune responses, promote growing and strengthen physique in Ictalurus punctatus. Aquacul. Nutr. 2021, 27, 1181–1191. [Google Scholar] [CrossRef]
- Kuhlwein, H.; Emery, M.J.; Rawling, M.D.; Harper, G.M.; Merrifield, D.L.; Davies, S.J. Effects of a dietary beta-(1,3)(1,6)-D-glucan supplementation on intestinal microbial communities and intestinal ultrastructure of mirror carp (Cyprinus carpio L.). J. Appl. Microbiol. 2013, 115, 1091–1106. [Google Scholar] [CrossRef]
- Jung-Schroers, V.; Adamek, M.; Jung, A.; Harris, S.; Doza, O.S.; Baumer, A.; Steinhagen, D. Feeding of beta-1,3/1,6-glucan increases the diversity of the intestinal microflora of carp (Cyprinus carpio). Aquacul. Nutr. 2016, 22, 1026–1039. [Google Scholar] [CrossRef]
- Miest, J.J.; Arndt, C.; Adamek, M.; Steinhagen, D.; Reusch, T.B. Dietary beta-glucan (MacroGard) enhances survival of first feeding turbot (Scophthalmus maximus) larvae by altering immunity, metabolism and microbiota. Fish Shellfish Immun. 2016, 48, 94–104. [Google Scholar] [CrossRef]
- de Souza, F.P.; de Lima, E.; Pandolfi, V.; Leite, N.; Furlan-Murari, P.; Leal, C.N.S.; Mainardi, R.M.; Suphoronski, S.A.; Favero, L.M.; Koch, J.F.A.; et al. Effect of beta-glucan in water on growth performance, blood status and intestinal microbiota in tilapia under hypoxia. Aquacult. Rep. 2020, 17, 100369. [Google Scholar] [CrossRef]
- Liang, H.; Li, Y.; Li, M.; Zhou, W.; Chen, J.; Zhang, Z.; Yang, Y.; Ran, C.; Zhou, Z. The effect and underlying mechanism of yeast β-glucan on antiviral resistance of zebrafish against spring viremia of carp virus infection. Front. Immunol. 2022, 13, 1031962. [Google Scholar] [CrossRef]
- Baldridge, M.T.; Nice, T.J.; McCune, B.T.; Yokoyama, C.C.; Kambal, A.; Wheadon, M.; Diamond, M.S.; Ivanova, Y.; Artyomov, M.; Virgin, H.W. Commensal microbes and interferon-λ determine persistence of enteric murine norovirus infection. Science 2015, 347, 266–269. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Jing, X.; Zhang, C.; Hou, Y.; Li, Z.; Yang, X.; Zhou, X.; Xu, P.; Tang, Y.; Zhu, J. Interaction Between the Intestinal Microbial Community and Transcriptome Profile in Common Carp (Cyprinus carpio L.). Front. Microbiol. 2021, 12, 659602. [Google Scholar] [CrossRef]
- Fei, H.; Yi, S.F.; Zhang, H.M.; Cheng, Y.; Zhang, Y.Q.; Yu, X.; Qian, S.C.; Huang, M.M.; Yang, S. Transcriptome and 16S rRNA analysis revealed the response of largemouth bass (Micropterus salmoides) to Rhabdovirus infection. Front. Immunol. 2022, 13, 973422. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, Q.; Xi, L.; Gong, Y.; Su, J.; Han, D.; Zhang, Z.; Liu, H.; Jin, J.; Yang, Y.; et al. Effects of Replacement of Dietary Fishmeal by Cottonseed Protein Concentrate on Growth Performance, Liver Health, and Intestinal Histology of Largemouth Bass (Micropterus salmoides). Front. Physiol. 2021, 12, 764987. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Han, Z.; Turchini, G.M.; Wang, X.; Fang, Z.; Chen, N.; Xie, R.; Zhang, H.; Li, S. Effects of Dietary Phospholipids on Growth Performance, Digestive Enzymes Activity and Intestinal Health of Largemouth Bass (Micropterus salmoides) Larvae. Front. Immunol. 2022, 12, 827946. [Google Scholar] [CrossRef] [PubMed]
- O’Doherty, J.V.; McDonnell, P.; Figat, S. The effect of dietary laminarin and fucoidan in the diet of the weanling piglet on performance and selected faecal microbial populations. Livest. Sci. 2020, 134, 208–210. [Google Scholar] [CrossRef]
- Rattigan, R.; Sweeney, T.; Maher, S.; Thornton, K.; Rajauria, G.; O’Doherty, J.V. Laminarin-rich extract improves growth performance, small intestinal morphology, gene expression of nutrient transporters and the large intestinal microbial composition of piglets during the critical post-weaning period. Br. J. Nutr. 2020, 123, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Rattigan, R.; Sweeney, T.; Maher, S.; Ryan, M.T.; Thornton, K.; O’Doherty, J.V. Effects of reducing dietary crude protein concentration and supplementation with either laminarin or zinc oxide on the growth performance and intestinal health of newly weaned pigs. Anim. Feed Sci. Technol. 2021, 270, 114693. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ke, F.; Zhang, Q.Y. Effect of beta-glucan on activity of antioxidant enzymes and Mx gene expression in virus infected grass carp. Fish Shellfish Immun. 2009, 27, 336–340. [Google Scholar] [CrossRef]
- Yamamoto, F.Y.; Castillo, S.; de Cruz, C.R.; Chen, K.; Hume, M.E.; Gatlin, D.M., III. Synergistic Effects of the β-1,3 Glucan Paramylon and Vitamin C on Immunological Responses of Hybrid Striped Bass (Morone Chrysops × M. Saxatilis) Were Pronounced In Vitro But More Moderate In Vivo. Aquaculture 2020, 526, 735394. [Google Scholar] [CrossRef]
- Yamamoto, F.Y.; Yin, F.; Rossi, W., Jr.; Hume, M.; Gatlin, D.M., III. β-1,3 Glucan Derived from Euglena Gracilis and Algamune™ Enhances Innate Immune Responses of Red Drum (Sciaenops Ocellatus L.). Fish Shellfish Immun. 2018, 77, 273–279. [Google Scholar] [CrossRef]
- El-Murr, A.I.; Abd El Hakim, Y.; Neamat-Allah, A.N.F.; Baeshen, M.; Ali, H.A. Immune-protective, antioxidant and relative genes expression impacts of beta-glucan against fipronil toxicity in Nile tilapia, Oreochromis niloticus. Fish Shellfish Immun. 2019, 94, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, O.; Sevgiler, N. Tissue-specific oxidative stress responses in fish exposed to 2,4-D and azinphosmethyl. Comp. Biochem. Physiol. Part C 2004, 137, 43–51. [Google Scholar] [CrossRef]
- Olsvika, P.A.; Kristensen, T.; Waagbø, R.; Rosseland, B.O.; Tollefsen, K.-E. mRNA expression of antioxidant enzymes (SOD, CAT and GSH-Px) and lipid peroxidative stress in liver of Atlantic salmon (Salmo salar) exposed to hyperoxic water during smoltification. Comp. Biochem. Physiol. Part C 2005, 141, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; Dawood, M.A.O.; Aleya, L.; Alkahtani, S. Effects of fucoidan on the hematic indicators and antioxidative responses of Nile tilapia (Oreochromis niloticus) fed diets contaminated with aflatoxin B(1). Environ. Sci. Pollut. R. 2020, 27, 12579–12586. [Google Scholar] [CrossRef] [PubMed]
- Vinosha, M.; Palanisamy, S.; Anjali, R.; Li, C.; Yelithao, K. Sulphated galactan from Halymenia dilatata enhance the antioxidant properties and prevents Aeromonas hydrophila infection in tilapia fish: In vitro and in vivo study. Int. J. Biol. Macromol. 2020, 158, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, L. Effects of chitosan-supplemented diets on the growth performance, nonspecific immunity and health of loach fish (Misgurnus anguillicadatus). Carbohyd. Polym. 2019, 225, 115227. [Google Scholar] [CrossRef]
- Huang, Q.; Zhu, Y.L.; Yu, J.; Fang, L.; Li, Y.A. Effects of sulfated beta-glucan from Saccharomyces cerevisiae on growth performance, antioxidant ability, nonspecific immunity, and intestinal flora of the red swamp crayfish (Procambarus clarkii). Fish Shellfish Immun. 2022, 127, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Peddie, S.; Scapigliati, G.; Zhang, Y.; Bols, N.C.; Ellis, A.E. Functional Characterisation of the Recombinant Tumor Necrosis Factors in Rainbow Trout, Oncorhynchus mykiss. Dev. Comp. Immunol. 2013, 27, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Falco, A.; Frost, P.; Miest, J.; Pionnier, N.; Irnazarow, I.; Hoole, D. Reduced inflammatoryb response to Aeromonas salmonicida infection in common carp (Cyprinus carpio L.) fed with β-glucan supplements. Fish Shellfish Immun. 2012, 32, 1051–1057. [Google Scholar] [CrossRef]
- Yang, X.; Wei, H.; Qin, L.; Zhang, S.; Wang, X.; Zhang, A. Reciprocal interaction between fish tgf-β1 and il-1β is responsible for restraining il-1β signaling activity in grass carp head kidney leukocytes. Dev. Comp. Immunol. 2014, 47, 197–204. [Google Scholar] [CrossRef]
- Feng, L.; Chen, Y.P.; Jiang, W.D.; Liu, Y.; Jiang, J.; Wu, P. Modulation of Immune Response, Physical Barrier and Related Signaling Factors in the Gills of Juvenile Grass Carp (Ctenopharyngodon Idella) Fed Supplemented Diet with Phospholipids. Fish Shellfish Immun. 2016, 48, 79–93. [Google Scholar] [CrossRef]
- Tan, P.; Dong, X.; Mai, K.; Xu, W.; Ai, Q. Vegetable Oil Induced Inflammatory Response by Altering TLR-NF-kb Signalling, Macrophages Infiltration and Polarization in Adipose Tissue of Large Yellow Croaker (Larimichthys Crocea). Fish Shellfish Immun. 2016, 59, 398–405. [Google Scholar] [CrossRef]
- Zhou, M.; Liang, R.; Mo, J.; Yang, S.; Gu, N.; Wu, Z.; Babu, V.S.; Li, J.; Huang, Y.; Lin, L. Effects of brewer’s yeast hydrolysate on the growth performance and the intestinal bacterial diversity of largemouth bass (Micropterus salmoides). Aquaculture 2018, 484, 139–144. [Google Scholar] [CrossRef]
- Xv, Z.; Zhong, Y.; Wei, Y.; Zhang, T.; Zhou, W.; Jiang, Y.; Chen, Y.; Lin, S. Yeast culture supplementation alters the performance and health status of juvenile largemouth bass (Micropterus salmoides) fed a high-plant protein diet. Aquacul. Nutr. 2021, 27, 2637–2650. [Google Scholar] [CrossRef]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Zhong, Y.; He, G.; Sun, H.; Chen, Y.; Zhou, W.; Lin, S. Yeast culture improved the growth performance, liver function, intestinal barrier and microbiota of juvenile largemouth bass (Micropterus salmoides) fed high-starch diet. Fish Shellfish Immun. 2022, 120, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Behera, B.K.; Bera, A.K.; Paria, P.; Das, A.; Parida, P.K.; Kumari, S.; Bhowmick, S.; Das, B.K. Identification and pathogenicity of Plesiomonas shigelloides in Silver Carp. Aquaculture 2018, 493, 314–318. [Google Scholar] [CrossRef]
- Wang, R.; Pan, X.; Xu, Y. Altered Intestinal Microbiota Composition Associated with Enteritis in Yellow Seahorses Hippocampus kuda (Bleeker, 1852). Curr. Microbiol. 2020, 77, 730–737. [Google Scholar] [CrossRef]
- Yuan, X.Y.; Zhang, X.T.; Xia, Y.T.; Zhang, Y.Q.; Wang, B.; Ye, W.W.; Ye, Z.F.; Qian, S.C.; Huang, M.M.; Yang, S.; et al. Transcriptome and 16S rRNA analyses revealed differences in the responses of largemouth bass (Micropterus salmoides) to early Aeromonas hydrophila infection and immunization. Aquaculture 2021, 541, 736759. [Google Scholar] [CrossRef]
- Chen, W.; Chang, K.; Chen, J.; Zhao, X.; Gao, S. Dietary sodium butyrate supplementation attenuates intestinal inflammatory response and improves gut microbiota composition in largemouth bass (Micropterus salmoides) fed with a high soybean meal diet. Fish Physiol. Biochem. 2021, 47, 1805–1819. [Google Scholar] [CrossRef]
- Xu, F.M.; Hou, S.W.; Wang, G.X.; Gong, J.Y.; Zhou, L.; Gao, S. Effects of zymolytic black soldier fly (Hermetia illucens) pulp as dietary supplementation in largemouth bass (Micropterus salmoides). Aquacul. Rep. 2021, 21, 100823. [Google Scholar] [CrossRef]
- Yang, H.; Bian, Y.; Huang, L.; Lan, Q.; Ma, L.; Li, X.; Leng, X. Effects of replacing fish meal with fermented soybean meal on the growth performance, intestinal microbiota, morphology and disease resistance of largemouth bass (Micropterus salmoides). Aquacul. Rep. 2022, 22, 100954. [Google Scholar] [CrossRef]
- Xie, X.; Wang, J.; Guan, Y.; Xing, S.; Liang, X.; Xue, M.; Wang, J.; Chang, Y.; Leclercq, E. Cottonseed protein concentrate as fishmeal alternative for largemouth bass (Micropterus salmoides) supplemented a yeast-based paraprobiotic: Effects on growth performance, gut health and microbiome. Aquaculture 2022, 551, 737898. [Google Scholar] [CrossRef]
- Lian, C.-A.; Yan, G.-Y.; Huang, J.-M.; Danchin, A.; Wang, Y.; He, L.-S. Genomic Characterization of a Novel Gut Symbiont from the Hadal Snailfish. Front. Microbiol. 2020, 10, 2978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheaib, B.; Yang, P.; Kazlauskaite, R.; Lindsay, E.; Heys, C.; Dwyer, T.; De Noia, M.; Schaal, P.; Sloan, W.; Ijaz, U.; et al. Genome erosion and evidence for an intracellular niche-Exploring the biology of mycoplasmas in Atlantic salmon. Aquaculture 2021, 541, 736772. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 2012, 95, 50–60. [Google Scholar] [CrossRef]
- Zhang, J.; Song, L.; Wang, Y.; Liu, C.; Zhang, L.; Zhu, S.; Liu, S.; Duan, L. Beneficial effect of butyrate-producing Lachnospiraceae on stress-induced visceral hypersensitivity in rats. J. Gastroen. Hepatol. 2019, 34, 1368–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, W.-K.; Koh, C.-B. The utilization and mode of action of organic acids in the feeds of cultured aquatic animals. Rev. Aquacult. 2017, 9, 342–368. [Google Scholar] [CrossRef]
- Zhang, Q.-L.; Li, H.-W.; Wu, W.; Zhang, M.; Guo, J.; Deng, X.-Y.; Wang, F.; Lin, L.-B. The Response of Microbiota Community to Streptococcus agalactiae Infection in Zebrafish Intestine. Front. Microbiol. 2019, 10, 2848. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, C.; Zhou, S. Isolation and identification of a humic substance and Fe(III)-reducing bacterial strain. J. Lanzhou Univ. 2010, 46, 56–65. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g kg−1) | Diets | |||
|---|---|---|---|---|
| Con | LL | ML | HL | |
| Fishmeal | 550 | 550 | 550 | 550 |
| Shrimp meal | 100 | 100 | 100 | 100 |
| Laminarin | 0 | 5 | 10 | 15 |
| Corn protein | 130 | 130 | 130 | 130 |
| Soybean meal | 60 | 60 | 60 | 60 |
| Soluble starch | 40 | 40 | 40 | 40 |
| Fish oil | 80 | 80 | 80 | 80 |
| Ca(H2PO4)2 | 10 | 10 | 10 | 10 |
| Mineral premix 1 | 10 | 10 | 10 | 10 |
| Vitamin premix 2 | 8 | 8 | 8 | 8 |
| Cr2O3 | 4 | 4 | 4 | 4 |
| Choline chloride | 3 | 3 | 3 | 3 |
| Threonine | 3 | 3 | 3 | 3 |
| Methionine | 2 | 2 | 2 | 2 |
| Proximate composition (%) | ||||
| Dry matter | 94.11 | 94.17 | 94.03 | 94.18 |
| Crude protein | 49.77 | 49.43 | 49.21 | 49.09 |
| Crude lipid | 9.53 | 9.50 | 9.44 | 9.39 |
| Ash | 10.78 | 10.62 | 10.72 | 10.69 |
| Gene | Sequence (5′-3′) | Tm (℃) | Product Size (bp) | Accession Number |
|---|---|---|---|---|
| il-1β | F: CGTGACTGACAGCAAAAAGAGG | 60 | 166 | XM_038733429.1 |
| R: GATGCCCAGAGCCACAGTTC | 61 | |||
| tnf-α | F: CTTCGTCTACAGCCAGGCATCG | 63 | 162 | XM_038710731.1 |
| R: TTTGGCCACACCGACCTCACC | 65 | |||
| tgf-β | F: GCTCAAAGAGAGCGAGGATG | 58 | 118 | XM_038693206.1 |
| R: TCCTCTACCATTCGCAATCC | 57 | |||
| β-actin | F: TGGAAGGGACCTCACAGACTAC | 61 | 231 | MH018565 |
| R: GGGCAACGGAACCTCTCAT | 60 |
| Item | Con | LL | ML | HL |
|---|---|---|---|---|
| IBW(g) | 0.72 ± 0.04 | 0.73 ± 0.02 | 0.72 ± 0.04 | 0.71 ± 0.04 |
| FBW(g) | 2.88 ± 0.21 | 3.00 ± 0.15 | 2.84 ± 0.30 | 2.75 ± 0.30 |
| WGR(%) | 297.38 ± 29.58 | 311.81 ± 19.85 | 296.37 ± 42.18 | 285.15 ± 42.30 |
| SGR(%/day) | 4.92 ± 0.56 | 5.05 ± 0.17 | 4.90 ± 0.37 | 4.80 ± 0.40 |
| Total feed intake (g) | 90.72 ± 2.12 | 92.62 ± 1.34 | 88. 19 ± 1.71 | 90.58 ± 1.86 |
| FCR | 1.05 ± 0.02 b | 1.02 ± 0.04 b | 1.04 ± 0.03 b | 1.11 ± 0.02 a |
| CF(g/cm³) | 1.04 ± 0.07 ab | 1.07 ± 0.07 a | 0.98 ± 0.06 b | 1.00 ± 0.05 b |
| HSI(%) | 1.76 ± 0.28 | 1.64 ± 0.04 | 1.67 ± 0.19 | 1.69 ± 0.14 |
| Item | Con | LL | ML | HL |
|---|---|---|---|---|
| Moisture (%) | 73.54 ± 0.36 | 73.97 ± 0.17 | 73.86 ± 0.22 | 73.50 ± 0.31 |
| Crude protein (%) | 15.74 ± 0.45 a | 16.41 ± 0.38 a | 15.72 ± 0.33 ab | 15.42 ± 0.29 b |
| Crude lipid (%) | 3.76 ± 0.14 | 3.84 ± 0.07 | 3.75 ± 0.07 | 3.72 ± 0.09 |
| Item | Con | LL | ML | HL |
|---|---|---|---|---|
| ACE | 3931.84 ± 735.78 a | 1722.66 ± 130.31 c | 2976.99 ± 137.85 b | 221.94 ± 43.89 d |
| Shannon | 10.74 ± 0.16 a | 8.00 ± 0.27 c | 9.03 ± 0.04 b | 2.91 ± 0.14 d |
| Simpson | 1.00 ± 0.00 a | 0.97 ± 0.02 b | 0.99 ± 0.00 a | 0.78 ± 0.01 c |
| Chao1 | 3927.94 ± 732.99 a | 1720.83 ± 130.43 c | 2966.06 ± 138.33 b | 220.46 ± 44.03 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Cheng, Y.; Qian, S.; Zhang, W.; Huang, M.; Yang, S.; Fei, H. An Evaluation of Laminarin Additive in the Diets of Juvenile Largemouth Bass (Micropterus salmoides): Growth, Antioxidant Capacity, Immune Response and Intestinal Microbiota. Animals 2023, 13, 459. https://doi.org/10.3390/ani13030459
Wu Y, Cheng Y, Qian S, Zhang W, Huang M, Yang S, Fei H. An Evaluation of Laminarin Additive in the Diets of Juvenile Largemouth Bass (Micropterus salmoides): Growth, Antioxidant Capacity, Immune Response and Intestinal Microbiota. Animals. 2023; 13(3):459. https://doi.org/10.3390/ani13030459
Chicago/Turabian StyleWu, Youjun, Yan Cheng, Shichao Qian, Wei Zhang, Mengmeng Huang, Shun Yang, and Hui Fei. 2023. "An Evaluation of Laminarin Additive in the Diets of Juvenile Largemouth Bass (Micropterus salmoides): Growth, Antioxidant Capacity, Immune Response and Intestinal Microbiota" Animals 13, no. 3: 459. https://doi.org/10.3390/ani13030459