
Priming European Sea Bass Female Broodstock Improves the Antimicrobial Immunity of Their Offspring

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Nodavirus-Expressing Vector Constructs
2.3. Experimental Design and Sampling Procedure
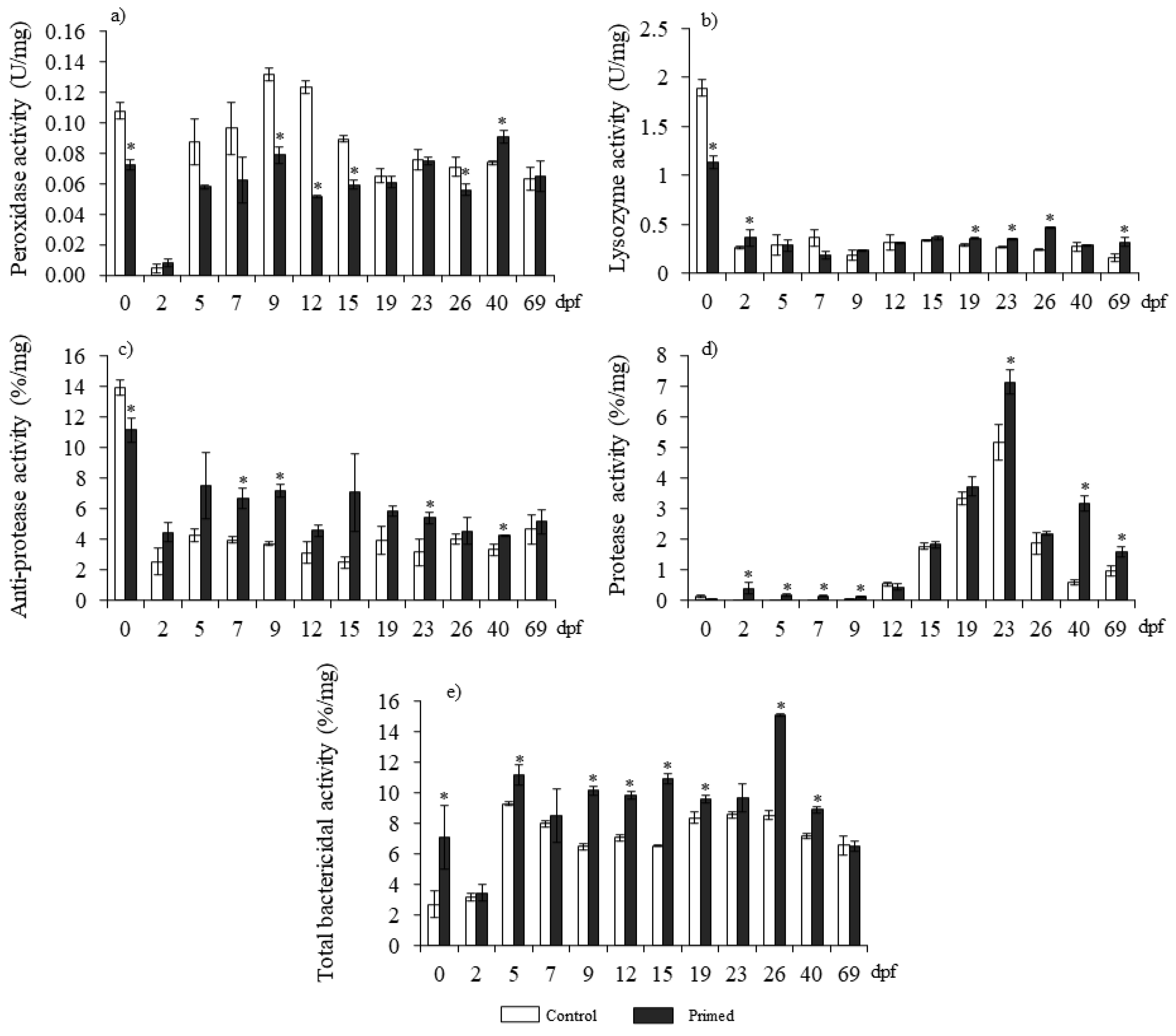
2.4. Functional Determination of Immunity
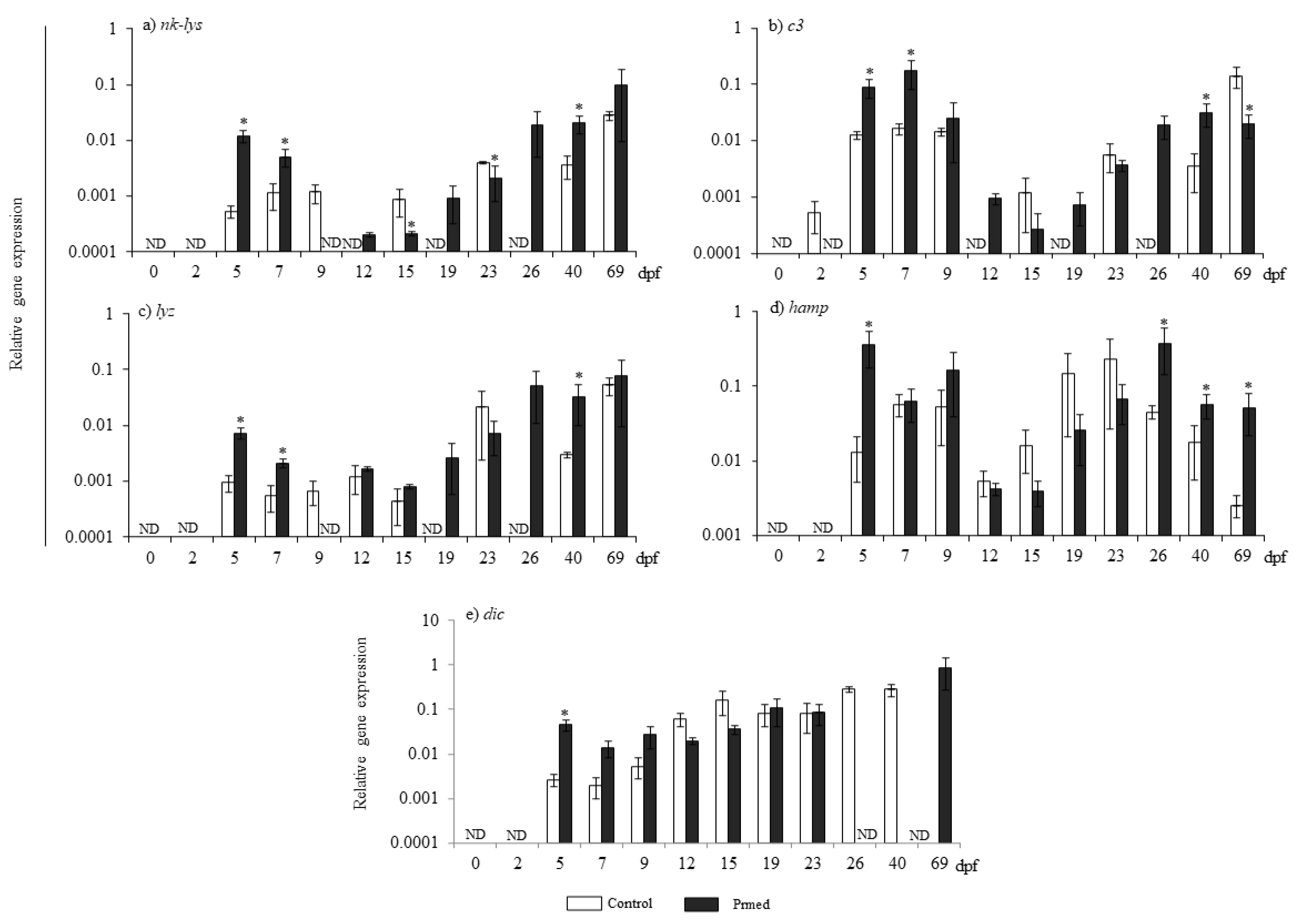
2.5. Gene Expression Analysis
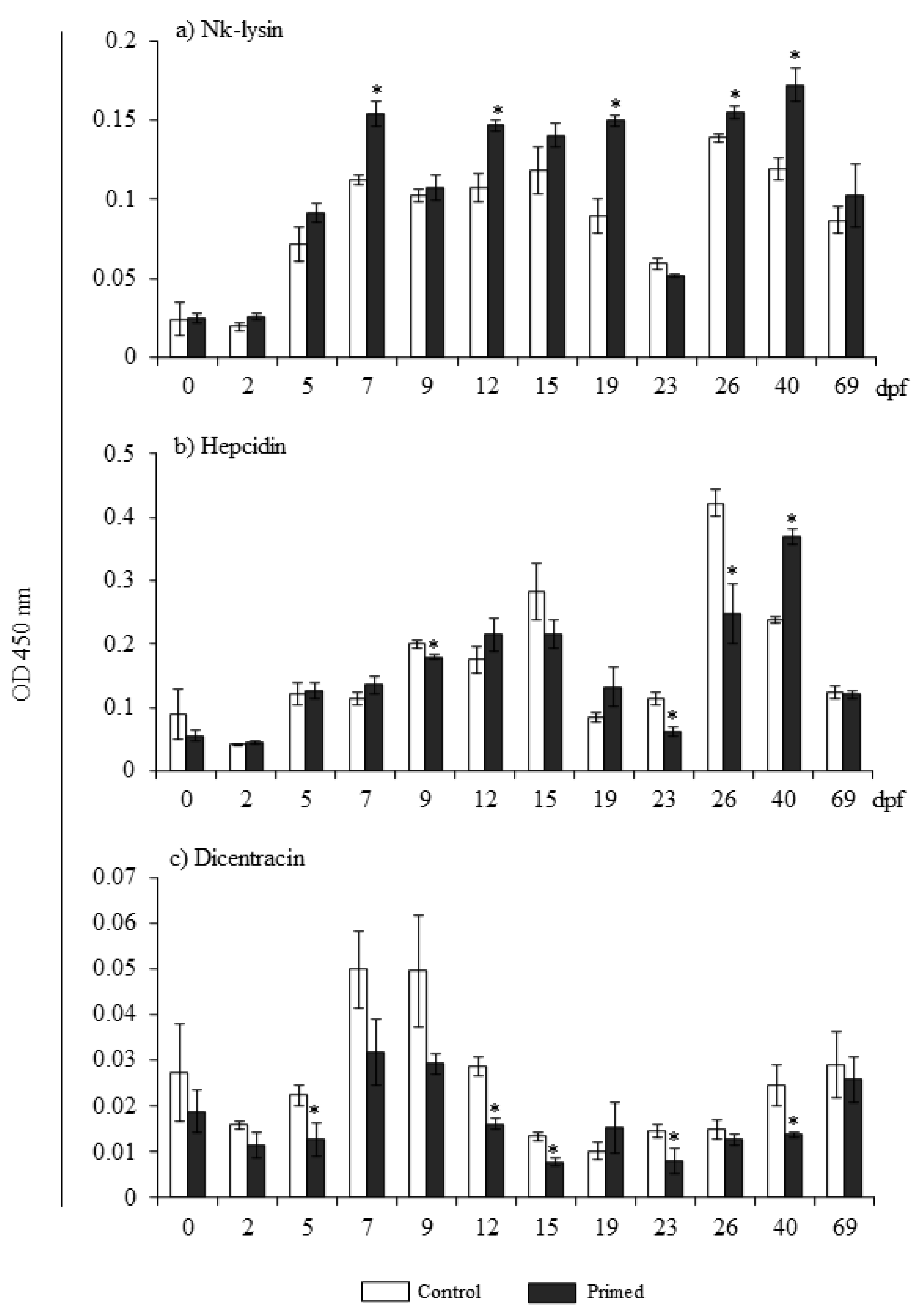
2.6. Peptide Quantification by ELISA
2.7. Nomenclature and Statistical Analysis
3. Results
3.1. Antimicrobial Activities Are Enhanced in Larvae from Vaccinated Females
3.2. AMPs Transcription Was Upregulated in the Progeny of CP-pNNV-Primed Females
3.3. Priming Broodstock Females Enhances Nk-Lysin Levels during Fry Development
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, S.; Wang, Z.; Wang, H. Maternal immunity in fish. Dev. Comp. Immunol. 2013, 39, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Mulero, I.; Garía-Ayala, A.; Meseguer, J.; Mulero, V. Maternal transfer of immunity and ontogeny of autologous immune competence of fish: A mini review. Aquaculture 2007, 268, 244–250. [Google Scholar] [CrossRef]
- DosSantos, N.M.; Romano, N.; deSousa, M.; Ellis, A.E.; Rombout, J.H. Ontogeny of Band T cells in sea bass (Dicentrarchus labrax, L.). Fish Shellfish Immunol. 2000, 10, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, S.; Terova, G.; Caricato, G.; Saroglia, M. Lysozyme activity in embryos and larvae of sea bass (Dicentrarchus labrax L.), spawned by brood stocks fed with vitamin C enriched diets. Bull. Eurn. Assoc. Fish. Pathol. 2000, 20, 120–124. [Google Scholar]
- Carnevali, O.; Mosconi, G.; Cambi, A.; Ridolfi, S.; Zanuy, S.; Polzonetti-Magni, A.M. Changes of lysosomal enzyme activities in sea bass (Dicentrarchus labrax) eggs and developing embryos. Aquaculture 2001, 202, 249–256. [Google Scholar] [CrossRef]
- Cho, J.H.; Park, I.Y.; Kim, M.S.; Kim, S.C. Matrix metalloproteinase 2 is involved in the regulation of the antimicrobial peptide parasin I production in catfish skin mucosa. FEBS Lett. 2002, 531, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Cucchi, P.; Sucré, E.; Santos, R.; Leclère, J.; Charmantier, G.; Castille, R. Embryonic development of the sea bass Dicentrarchus labrax. Helgol. Mar. Res. 2011, 66, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Mondal, H.; Thomas, J. Are view on the recent advances and application of vaccines against fish pathogens in aquaculture. Aquacult. Int. 2022, 30, 1971–2000. [Google Scholar] [CrossRef]
- Hanif, A.; Bakopoulos, V.; Dimitriadis, G.J. Maternal transfer of humoral specific and non-specific immune parameters to seabream (Sparus aurata) larvae. Fish Shellfish Immunol. 2004, 17, 411–435. [Google Scholar] [CrossRef]
- Hanif, A.; Bakopoulos, V.; Leonardos, I.; Dimitriadis, G.J. The effect of seabream (Sparus aurata) broodstock and larval vaccination on the susceptibility by Photobacterium damselae subsp. piscicida and on the humoral immune parameters. Fish Shellfish Immunol. 2005, 19, 345–361. [Google Scholar] [CrossRef]
- Nurani, F.S.; Sukenda, S.; Nuryati, S. Maternal immunity of tilapia broodstock vaccinated with polyvalent vaccine and resistance of their offspring against Streptococcus agalactiae. Aquac. Res. 2020, 51, 1513–1522. [Google Scholar] [CrossRef]
- VanderMeer, J.W.M.; Joosten, L.A.B.; Riksen, N.; Netea, M.G. Trained immunity: A smart way to enhance innate immune defence. Mol. Immunol. 2015, 68, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Quintin, J.; Saeed, S.; Martens, J.H.A.; Giamarellos-Bourboulis, E.J.; Ifrim, D.C.; Logie, C.; Jacobs, L.; Jansen, T.; Kullberg, B.J.; Wijmenga, C.; et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microb. 2012, 12, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Chi, H.; Dalmo, R.A. Trained innate immunity of fish is a viable approach in larval aquaculture. Front. Immunol. 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beemelmanns, A.; Roth, O. Biparental immune priming in the pipefish Syngnathus typhle. Zoology 2016, 119, 262–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roth, O.; Joop, G.; Eggert, H.; Hilbert, J.; Daniel, J.; Schmid-Hempel, P.; Kurtz, J. Paternally derived immune priming for offspring in the redflour beetle, Tribolium castaneum. J. Anim. Ecol. 2010, 79, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Sandalova, E.; Low, D.; Gehring, A.J.; Fieni, S.; Amadei, B.; Urbani, S.; Chong, Y.S.; Guccione, E.; Bertoletti, A. Trained immunity in new born infants of HBV-infected mothers. Nat. Commun. 2015, 6, 6588. [Google Scholar] [CrossRef] [Green Version]
- Norouzitallab, P.; Baruah, K.; Biswas, P.; Vanrompay, D.; Bossier, P. Probing the phenomenon of trained immunity in invertebrates during a transgenerational study, using brine shrimp Artemia as a model system. Sci. Rep. 2016, 6, 21166. [Google Scholar] [CrossRef] [Green Version]
- Green, T.J.; Helbig, K.; Speck, P.; Raftos, D.A. Primedforsuccess:Oyster parents treated with poly(I:C) produce offspring with enhanced protection against Ostreid herpes virus type I infection. Mol. Immunol. 2016, 78, 113–120. [Google Scholar] [CrossRef]
- Beemelmanns, A.; Roth, O. Grand parental immune priming in the pipefish Syngnathus typhle. BMC Evol. Biol. 2017, 17, 44. [Google Scholar] [CrossRef] [Green Version]
- Valero, Y.; Saraiva-Fraga, M.; Costas, B.; Guardiola, F.A. Fish antimicrobial peptides: Beyond the fight against pathogens. Rev. Aquac. 2020, 12, 224–253. [Google Scholar] [CrossRef]
- Meloni, M.; Candusso, S.; Galeotti, M.; Volpatti, D. Preliminary study on expression of antimicrobial peptides in European sea bass (Dicentrarchus labrax) following in vivo infection with Vibrio anguillarum. A time course experiment. Fish Shellfish Immunol. 2015, 43, 82–90. [Google Scholar] [CrossRef]
- Valero, Y.; García-Alcázar, A.; Esteban, M.A.; Cuesta, A.; Chaves-Pozo, E. Antimicrobial response is increased in the testis of European sea bass, but not in gilthead seabream, upon nodavirus infection. Fish Shellfish Immunol. 2015, 44, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; Arizcun, M.; Cortés, J.; Ramírez-Cepeda, F.; Guzmán, F.; Mercado, L.; Esteban, M.Á.; Chaves-Pozo, E.; Cuesta, A. NK-lysin, dicentracin and hepcidin antimicrobial peptides in European sea bass. Ontogenetic development and modulation in juveniles by nodavirus. Dev. Comp. Immunol. 2020, 103, 103516. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.A.; Acosta, F.; Montero, D.; Guzman, F.; Torres, E.; Vega, B.; Mercado, L. Synthetic hepcidin from fish: Uptake and protection against Vibrio anguillarum in sea bass (Dicentrarchus labrax). Fish Shellfish Immunol. 2016, 55, 662–670. [Google Scholar] [CrossRef]
- León, R.; Ruiz, M.; Valero, Y.; Cárdenas, C.; Guzmán, F.; Vila, M.; Cuesta, A. Exploring small cationic peptides of different origin as potential antimicrobial agents in aquaculture. Fish Shellfish Immunol. 2020, 98, 720–727. [Google Scholar] [CrossRef]
- Valero, Y.; Awad, E.; Buonocore, F.; Arizcun, M.; Esteban, M.A.; Meseguer, J.; Chaves-Pozo, E.; Cuesta, A. An oral chitosan DNA vaccine against nodavirus improves transcription of cell-mediated cytotoxicity and interferon genes in the European sea bass juveniles gut and survival upon infection. Dev. Comp. Immunol. 2016, 65, 64–72. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal.Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef]
- Valero, Y.; López-Cánovas, A.E.; Rodenas, M.C.; Cabas, I.; García-Hernández, P.; Arizcun, M.; García-Ayala, A.; Chaves-Pozo, E. Endocrine disrupter chemicals affect the humoral antimicrobial activities of gilthead seabream males even upon the cease of the exposure. Sci. Rep. 2020, 10, 7966. [Google Scholar] [CrossRef]
- Charney, J.; Tomarelli, R.M. A colorimetric method for the determination of the proteolytic activity of duodenal juice. J. Biol. Chem. 1947, 171, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Ellis, A.E. Serum antiproteases in fish. In Techniques in Fish Immunology; Stolen, J.S., Fletcher, T.C., Anderson, D.P., Roberson, B.S., vanMuiswinkel, W.B., Eds.; SOSPublications: FairHaven, NJ, USA, 1990; pp. 95–99. [Google Scholar]
- Parry, R.M., Jr.; Chandan, R.C.; Shahani, K.M. A rapid and sensitive assay of muramidase. Proc. Soc. Exp. Biol. Med. 1965, 119, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Valero, Y.; García-Alcázar, A.; Esteban, M.A.; Cuesta, A.; Chaves-Pozo, E. Seasonal variations of the humoral immune parameters of European sea bass (Dicentrarchus labraxL.). Fish Shellfish Immunol. 2014, 39, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Sunyer, J.O.; Tort, L. Natural haemolytic and bactericidal activities of seabream Sparus aurata serum are affected by the alternative complement pathway. Vet. Immunol. Immunopathol. 1995, 45, 333–345. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Guardiola, F.A.; Meseguer, J.; Esteban, M.A.; Cuesta, A. Nodavirus infection induces a great innate cell-mediated cytotoxic activity in resistant, gilthead seabream, and susceptible, European sea bass, teleost fish. Fish Shellfish Immunol. 2012, 33, 1159–1166. [Google Scholar] [CrossRef]
- Nishizawa, T.; Mori, K.; Nakai, T.; Furusawa, I.; Muroga, K. Polymerase chain reaction (PCR) amplification of RNA of striped jack nervous necrosis virus (SJNNV). Dis.Aquat. Organ 1994, 18, 103–107. [Google Scholar] [CrossRef]
- Santana, P.A.; Álvarez, C.A.; Guzmán, F.; Mercado, L. Development of a sandwich ELISA for quantifying hepcidinin Rainbow trout. Fish Shellfish Immunol. 2013, 35, 748–755. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Swain, P.; Nayak, S.K. Role of maternally derived immunity in fish. Fish Shellfish Immunol. 2009, 27, 89–99. [Google Scholar] [CrossRef]
- Anderon, E.D.; Mourich, D.V.; Leong, J.A. Gene expression in rainbow trout (Oncorhynchus mykiss) following intramuscular injection of DNA. Mol. Mar. Biol.Biotechnol. 1996, 5, 105–113. [Google Scholar]
- Collins, C.; Lorenzen, N.; Collet, B. DNA vaccination for finfish aquaculture. Fish Shellfish Immunol. 2019, 85, 106–125. [Google Scholar] [CrossRef] [PubMed]
- Kanellos, T.; Sylvester, I.D.; Ambali, A.G.; Howard, C.R.; Russell, P.H. The safety and longevity of DNA vaccines for fish. Immunology 1999, 96, 307–313. [Google Scholar] [CrossRef]
- Guo, Y.X.; Wei, T.; Dallmann, K.; Kwang, J. Induction of caspase-dependent apoptosis by betanodaviruses GGNNV and demonstration of protein as an apoptosis inducer. Virology 2003, 308, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Magnadottir, B. Innate immunity of fish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Bugla-Plskonska, G.; Kiersnowski, A.; Futoma-Koloch, B.; Doroszkiewicz, W. Cooperation between lysozyme and complementsystem in bactericidal action of human serum–is everything already clear? Cent. Eur. J. Immunol. 2008, 33, 37–42. [Google Scholar]
- Ogundele, M.O. A novel anti-inflammatory activity of lysozyme: Modulation of serum complement activation. Mediat. Inflamm. 1998, 7, 363–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardlaw, A.C. The complement-dependent bacteriolytic activity of normal human serum. I. The effect of pH and ionic strength and the role of lysozyme. J. Exp. Med. 1962, 115, 1231–1249. [Google Scholar] [CrossRef]
- Huttenhuis, H.B.; Grou, C.; Taverne-Thiele, A.J.; Taverne, N.; Rombout, J.H. Carp (Cyprinus carpio L.) innate immune factors are present before hatching. Fish Shellfish Immunol. 2006, 20, 586–596. [Google Scholar] [CrossRef]
- Chettri, J.K.; Raida, M.K.; Kania, P.W.; Buchmann, K. Differential immune response of rainbow trout (Oncorhynchus mykiss) at early developmental stages (larvae and fry) against the bacterial pathogen Yersinia ruckeri. Dev. Comp. Immunol. 2012, 36, 463–474. [Google Scholar] [CrossRef]
- Santana, P.A.; Guzmán, F.; Forero, J.C.; Luna, O.F.; Mercado, L. Hepcidin, Cathelicidin-1 and IL-8 as immunological markers of responsiveness in early developmental stages of rainbow trout. Dev. Comp. Immunol. 2016, 62, 48–57. [Google Scholar] [CrossRef]
- Katzenback, B.A.; Katakura, F.; Belosevic, M. Regulation of teleost macrophage and neutrophil cell development by growth factors and transcription factors. In New Advances and Contributions to Fish Biology; InTech: Rijeka, Croatia, 2012; pp. 97–150. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Molecule | Gene | Accession Number | Sequence (5′ →3′) | |
|---|---|---|---|---|
| NNV capsid | cp | D38636 | F2 | CGTGTCAGTCATGTGTCGCT |
| R3 | CGAGTCAACACGGGTGAAGA | |||
| Hepcidin | hamp | DQ131605 | F | CCAGTCACTGAGGTGCAAGA |
| R | GCTGTGACGCTTGTGTCTGT | |||
| Complement factor 3-1 and 3-2 | c3 | HM563079 HM563078 | F | ACCAAAGAACTGGCAACCAC |
| R | CTAGCAGTCGGTCAGGGAAC | |||
| Lysozyme | lyz | FN667957 | F | ATTTCCTGGCTGGAACACAG |
| R | GAGCTCTGGCAACAACATCA | |||
| Nk-lysin | nk-lys | KY801205 | F | GAAGAAACACCTCGGGGAAT |
| R | GCAGGTCCAACATCTCCTTC | |||
| Dicentracin | dic | AY303949 | F | GGCAAGTCCATCCACAAACT |
| R | ATATTGCTCCGCTTGCTGAT | |||
| Elongation factor 1 alpha | ef1a | FM019753 | F | CGTTGGCTTCAACATCAAGA |
| R | GAAGTTGTCTGCTCCCTTGG | |||
| Ribosomal protein L13 alpha | l13a | DT044539 | F | GCGAAGGCATCAACATCTCC |
| R | AGACGCACAATCTTGAGAGCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valero, Y.; Mercado, L.; Arizcun, M.; Cuesta, A.; Chaves-Pozo, E. Priming European Sea Bass Female Broodstock Improves the Antimicrobial Immunity of Their Offspring. Animals 2023, 13, 415. https://doi.org/10.3390/ani13030415
Valero Y, Mercado L, Arizcun M, Cuesta A, Chaves-Pozo E. Priming European Sea Bass Female Broodstock Improves the Antimicrobial Immunity of Their Offspring. Animals. 2023; 13(3):415. https://doi.org/10.3390/ani13030415
Chicago/Turabian StyleValero, Yulema, Luis Mercado, Marta Arizcun, Alberto Cuesta, and Elena Chaves-Pozo. 2023. "Priming European Sea Bass Female Broodstock Improves the Antimicrobial Immunity of Their Offspring" Animals 13, no. 3: 415. https://doi.org/10.3390/ani13030415