
Gross, Histologic and Immunohistochemical Characteristics of Keratoacanthomas in Lizards

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Histopathology
2.3. Immunohistochemistry
3. Results
3.1. Clinical History and Gross Pathology
3.2. Histopathology and IHC
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Christman, J.; Devau, M.; Wilson-Robles, H.; Hoppes, S.; Rech, R.; Russell, K.E.; Heatley, J.J. Oncology of Reptiles: Diseases, Diagnosis, and Treatment. Vet. Clin. N. Am. Exot. Anim. Pract. 2017, 20, 87–110. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.M.; Hernandez-Divers, S.M.; Raymond, J.T. Reptile neoplasia: A retrospective study of case submissions to a specialty diagnostic service. Vet. Clin. N. Am. Exot. Anim. Pract. 2004, 7, 653–671. [Google Scholar] [CrossRef] [PubMed]
- Monahan, C.F.; Garner, M.M.; Kiupel, M. Chromatophoromas in Reptiles. Vet. Sci. 2022, 9, 115. [Google Scholar] [CrossRef] [PubMed]
- Hannon, D.E.; Garner, M.M.; Reavill, D.R. Squamous Cell Carcinomas in Inland Bearded Dragons (Pogona vitticeps). J. Herpetol. Med. Surg. 2011, 21, 101–106. [Google Scholar] [CrossRef]
- Hellebuyck, T.; Ducatelle, R.; Bosseler, L.; Van Caelenberg, A.; Versnaeyen, H.; Chiers, K.; Martel, A. Basal cell carcinoma in two Hermann’s tortoises (Testudo hermanni). J. Vet. Diagn. Investig. 2016, 28, 750–754. [Google Scholar] [CrossRef] [Green Version]
- Kubiak, M.; Denk, D.; Stidworthy, M.F. Retrospective review of neoplasms of captive lizards in the United Kingdom. Vet. Rec. 2020, 186, 28. [Google Scholar] [CrossRef]
- Meyer, J.; Kolodziejek, J.; Häbich, A.C.; Dinhopl, N.; Richter, B. Multicentric Squamous Cell Tumors in Panther Chameleons (Furcifer pardalis). J. Exot. Pet Med. 2019, 29, 166–172. [Google Scholar] [CrossRef]
- Murphy, G.; Beer, T.; Cerio, R.; Kao, G.; Nagore, E.; Pulitzer, M. Keratinocytic/epidermal tumours. In World Health Organization (WHO) Classification of Skin Tumours, 4th ed.; Elder, D., Massi, D., Scolyer, R., Willemze, R., Eds.; International Agency for Research on Cancer (IARC): Argonay, France, 2018; Volume 11, pp. 36–38. [Google Scholar]
- Williams, S.M.; Reece, R.L.; Hafner, S. Neoplastic diseases. In Diseases of Poultry, 14th ed.; Swayne, D.E., Glisson, J.R., McDougald, L.R., Nolan, L.K., Suarez, D.L., Nair, V., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2019; Volume 2, pp. 652–653. [Google Scholar]
- Abramo, F.; Pratesi, F.; Cantile, C.; Sozzi, S.; Poli, A. Survey of canine and feline follicular tumours and tumour-like lesions in central Italy. J. Small Anim. Pract. 1999, 40, 479–481. [Google Scholar] [CrossRef]
- Scott, D.W.; Miller, W.H.; Griffin, C.E. Neoplastic and Non-Neoplastic Tumors. In Muller & Kirk’s Small Animal Dermatology, 6th ed.; Scott, D.W., Miller, W.H., Griffin, C.E., Eds.; Elsevier Health Sciences: St. Louis, MO, USA, 2001; Volume 1, pp. 1236–1414. [Google Scholar]
- Mauldin, E.A.; Peters-Kennedy, J. Integumentary System. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals, 6th ed.; Maxie, M.G., Ed.; Elsevier Health Sciences: St. Louis, MO, USA, 2015; Volume 1, pp. 509–736. [Google Scholar]
- Gleich, T.; Chiticariu, E.; Huber, M.; Hohl, D. Keratoacanthoma: A distinct entity? Exp. Dermatol. 2016, 25, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Cerroni, L.; Kerl, H. Keratoacanthoma. In Fitzpatrick’s Dermatology in General Medicine, 7th ed.; Goldsmith, L.A., Katz, S.I., Gilchrest, B.A., Paller, A.S., Leffell, D.J., Wolff, K., Eds.; McGraw-Hill: New York, NY, USA, 2012; Volume 2, pp. 1049–1053. [Google Scholar]
- Lonsdorf, A.; Hadaschick, E. Squamous cell carcinoma and keratoacanthoma. In Fitzpatrick’s Dermatology, 9th ed.; Kang, S., Amagai, M., Bruckner, A.L., Enk, A.H., Margolis, D.J., McMichael, A.J., Orringer, J.S., Eds.; McGraw-Hill: New York, NY, USA, 2019; Volume 2, pp. 1901–1916. [Google Scholar]
- Wobeser, B.K.; Kidney, B.A.; Powers, B.E.; Withrow, S.J.; Mayer, M.N.; Spinato, M.T.; Allen, A.L. Agreement among surgical pathologists evaluating routine histologic sections of digits amputated from cats and dogs. J. Vet. Diagn. Investig. 2007, 19, 439–443. [Google Scholar] [CrossRef] [Green Version]
- Fedchenko, N.; Reifenrath, J. Different approaches for interpretation and reporting of immunohistochemistry analysis results in the bone tissue—A review. Diagn. Pathol. 2014, 9, 221. [Google Scholar] [CrossRef] [Green Version]
- Baines, F.; Chattell, J.; Dale, J.; Garrick, D.; Gill, I.; Goetz, M.; Skelton, T.; Swatman, M. How much UV-B does my reptile need? The UV-Tool, a guide to the selection of UV lighting for reptiles and amphibians in captivity. J. Zoo Aquar. Res. 2016, 4, 42–63. [Google Scholar]
- Boyer, T.H.; Scott, P.W. Nutritional therapy. In Mader’s Reptile and Amphibian Medicine and Surgery, 3rd ed.; Divers, S.J., Stahl, S.J., Eds.; Elsevier Health Sciences: St. Louis, MO, USA, 2019; Volume 1, pp. 1173–1176. [Google Scholar]
- Sánchez Yus, E.; Simón, P.; Requena, L.; Ambrojo, P.; de Eusebio, E. Solitary keratoacanthoma: A self-healing proliferation that frequently becomes malignant. Am. J. Dermatopathol. 2000, 22, 305–310. [Google Scholar]
- Weedon, D.D.; Malo, J.; Brooks, D.; Williamson, R. Squamous cell carcinoma arising in keratoacanthoma: A neglected phenomenon in the elderly. Am. J. Dermatopathol. 2010, 32, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Hardon, T.; Fledelius, B.; Heegaard, S. Keratoacanthoma of the spectacle in a Boa constrictor. Vet. Ophthalmol. 2007, 10, 320–322. [Google Scholar] [CrossRef] [PubMed]
- McGeough, R. Furcifer pardalis (Panther Chameleon)—A brief species description and details on captive husbandry. Biol. Eng. Med. Sci. Reports. 2016, 2, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Schmid-Brunclik, N.; Stefka, S.C.; Madeleine, K.B.; Max, G.; Jean-Michel, H. Liposarcoma in a Veiled Chameleon, Chamaeleo calyptratus. J. Herpetol. Med. Surg. 2008, 17, 132–135. [Google Scholar] [CrossRef] [Green Version]
- Paré, J.A.; Lentini, A.M. Reptile Geriatrics. Vet. Clin. N. Am. Exot. Anim. Pract. 2010, 13, 15–25. [Google Scholar] [CrossRef]
- Zargaran, M.; Baghaei, F. A clinical, histopathological and immunohistochemical approach to the bewildering diagnosis of keratoacanthoma. J. Dent. 2014, 15, 91–97. [Google Scholar]
- Ogita, A.; Ansai, S.I. What is a solitary keratoacanthoma? A benign follicular neoplasm, frequently associated with squamous cell carcinoma. Diagnostics 2021, 11, 1848. [Google Scholar] [CrossRef]
- Mitrache, C.; Benea, V.; Tovaru, M.; Georgescu, S.R.; Tudose., I. Clinical, epidemiological, physiopathological and histopathological aspects of keratoacanthoma. DermatoVenerol 2011, 56, 209–215. [Google Scholar]
- Wiener, D.J. Histologic features of hair follicle neoplasms and cysts in dogs and cats: A diagnostic guide. J. Vet. Diagn. Investig. 2021, 33, 479–497. [Google Scholar] [CrossRef] [PubMed]
- Waldman, A.; Schmults, C. Cutaneous Squamous Cell Carcinoma. Hematol. Oncol. Clin. N. Am. 2019, 33, 1–12. [Google Scholar] [CrossRef]
- Burton, K.A.; Ashack, K.A.; Khachemoune, A. Cutaneous Squamous Cell Carcinoma: A Review of High-Risk and Metastatic Disease. Am. J. Clin. Dermatol. 2016, 17, 491–508. [Google Scholar] [CrossRef] [PubMed]
- Hafner, S.; Harmon, B.G.; Stewart, R.G.; Rowland, G.N. Avian Keratoacanthoma (Dermal Squamous Cell Carcinoma) in Broiler Chicken Carcasses. Vet. Pathol. 1993, 30, 265–270. [Google Scholar] [CrossRef]
- Vîlcea, A.M.; Stoica, L.E.; Georgescu, C.V.; Popescu, F.C.; Ciurea, R.N.; Vîlcea, I.D.; Mirea, C.S. Clinical, histopathological and immunohistochemical study of keratoacanthoma. Rom. J. Morphol. Embryol. 2021, 62, 445–456. [Google Scholar] [CrossRef]
- Schwartz, R.A. Keratoacanthoma. J. Am. Acad. Dermatol. 1994, 30, 1–19. [Google Scholar] [CrossRef]
- Vargo, N. Basal cell and squamous cell carcinoma. Semin. Oncol. Nurs. 2003, 19, 12–21. [Google Scholar] [CrossRef]
- Rook, A.; Whimster, I. Keratoacanthoma—A thirty year retrospect. Br. J. Dermatol. 1979, 100, 41–47. [Google Scholar] [CrossRef]
- Kwiek, B.; Schwartz, R.A. Keratoacanthoma (KA): An update and review. J. Am. Acad. Dermatol. 2016, 74, 1220–1233. [Google Scholar] [CrossRef]
- Kubiak, M. Chameleons. In Handbook of Exotic Pet Medicine, 1st ed.; Kubiak, M., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2020; Volume 1, pp. 263–281. [Google Scholar]
- Ito, Y.; Kurokawa, I.; Nishimura, K.; Hakamada, A.; Isoda, K.I.; Yamanaka, K.I.; Tsubura, A.; Mizutani, H. Keratin and filaggrin expression in keratoacanthoma. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 353–355. [Google Scholar] [CrossRef]
- Chang, C.; Wu, P.; Baker, R.E.; Maini, P.K.; Alibardi, L.; Chuong, C.M. Reptile scale paradigm: Evo-Devo, pattern formation and regeneration. Int. J. Dev. Biol. 2009, 53, 813–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alibardi, L.; Toni, M. Wound keratins in the regenerating epidermis of lizard suggest that the wound reaction is similar in the tail and limb. J. Exp. Zool. A Comp. Exp. Biol. 2005, 303, 845–860. [Google Scholar] [CrossRef]
- Alibardi, L.; Maurizii, M.; Taddei, C. Immunocytochemical and electrophoretic distribution of cytokeratins in the resting stage epidermis of the lizard Podarcis sicula. J. Exp. Zool. 2001, 289, 409–418. [Google Scholar] [CrossRef]
- Jones, A.L.; Suárez-Bonnet, A.; Mitchell, J.A.; Ramirez, G.A.; Stidworthy, M.F.; Priestnall, S.L. Avian Papilloma and Squamous Cell Carcinoma: A Histopathological, Immunohistochemical and Virological study. J. Comp. Pathol. 2020, 175, 13–23. [Google Scholar] [CrossRef]
- Papadavid, E.; Pignatelli, M.; Zakynthinos, S.; Krausz, T.; Chu, A.C. The potential role of abnormal E-cadherin and a-, b- and g-catenin immunoreactivity in the determination of the biological behaviour of keratoacanthoma. Br. J. Dermatol. 2001, 145, 582–589. [Google Scholar] [CrossRef]
- Orós, J.; López-Yánez, M.; Rodríguez, F.; Calabuig, P.; Castro, P.L. Immunohistochemical staining patterns of alpha-keratins in normal tissues from two reptile species: Implications for characterization of squamous cell carcinomas. BMC Vet. Res. 2018, 14, 219. [Google Scholar] [CrossRef]
- Slater, M.; Barden, J.A. Differentiating keratoacanthoma from squamous cell carcinoma by the use of apoptotic and cell adhesion markers. Histopathology 2005, 47, 170–178. [Google Scholar] [CrossRef]
- Putti, T.C.; Teh, M.; Lee, Y.S. Biological behavior of keratoacanthoma and squamous cell carcinoma: Telomerase activity and COX-2 as potential markers. Mod. Pathol. 2004, 17, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Hua, H.K.; Jin, C.; Yang, L.J.; Tao, S.Q.; Zhu, X.H. Expression of Cyclooxygenase-2 in Squamous Cell Carcinoma and Keratoacanthoma and its Clinical Significance. Cell Biochem. Biophys. 2015, 72, 475–480. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
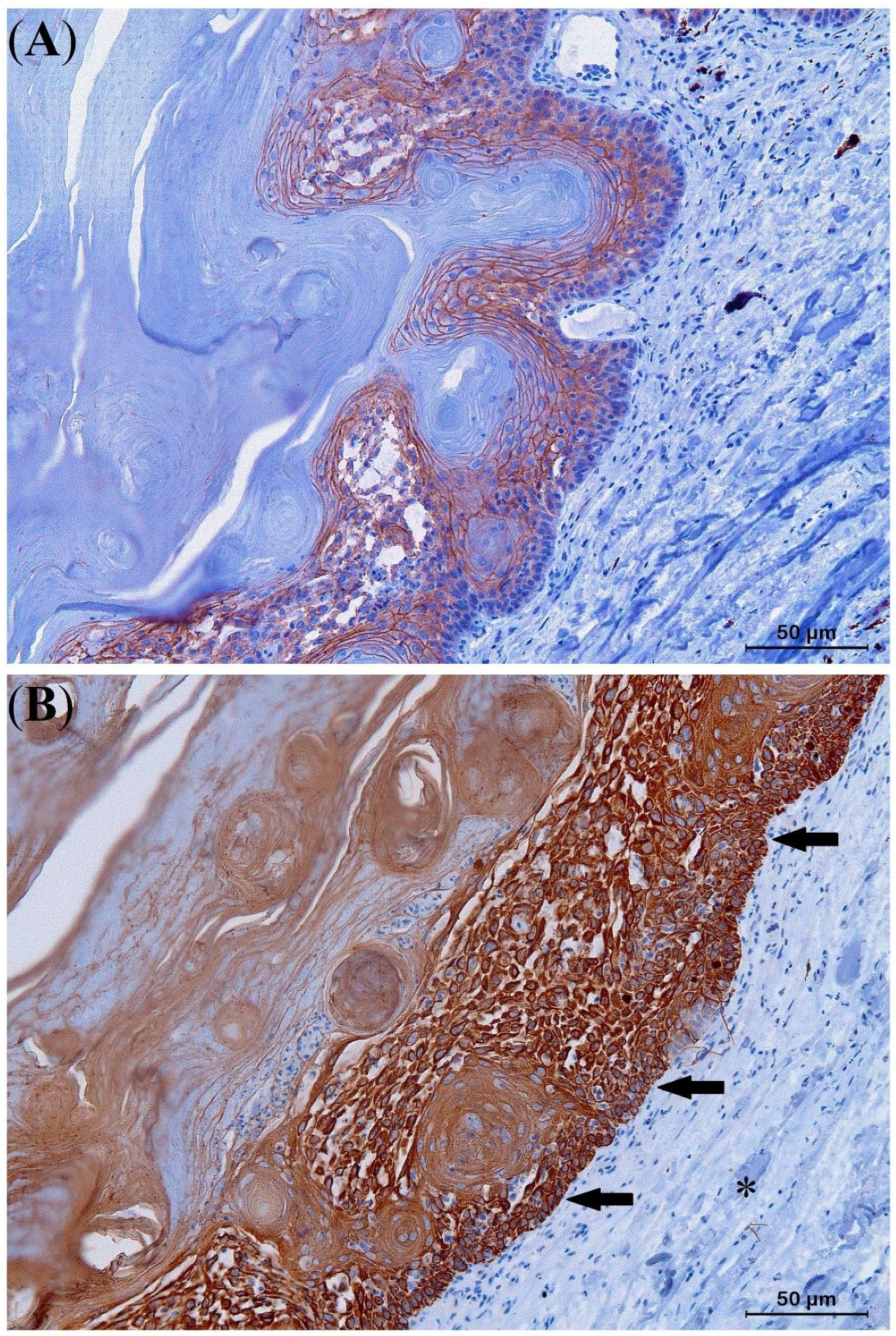{kind=link}
{kind=link}
| Species | Panther Chameleon 1 (Furcifer pardalis) | Panther Chameleon 2 (Furcifer pardalis) | Panther Chameleon 3 (Furcifer pardalis) | Veiled Chameleon (Chamaeleo calyptratus) | Bearded Dragon (Pogona vitticeps) |
|---|---|---|---|---|---|
| Age | 5 years | 4 years | 4 years | 3 years | 6 years |
| N° of nodular skin lesions at initial presentation | 18 | 15 | 3 | 4 | 2 |
| Location of the nodular skin lesions | Eyelid, dorsolateral body wall | Dorsolateral body wall | Eyelid, dorsolateral body wall | Eyelid, head, dorsolateral body wall | Dorsolateral body wall |
| Time between first detection and initial presentation | 2 years | 6 months | 3 months | 2 months | 4 months |
| Follow-up period | 2 years | 2 years | 1 year | 2 years | 1 year |
| Recurrence | No | No | No | 16 | 1 |
| UV-B source | ExoTerra Solar Glo 80 W | ZooMed Powersun 100 W | ZooMed Powersun 100 W | Arcadia D3 Forest | JBL UV-B Spot Plus 80 W |
| Average basking distance | 16 cm | 15 cm | 18 cm | 20 cm | 26 cm |
| A (Percentage of Positive Cells) | B (Intensity of Staining) | IRS Score (Multiplication of A and B) |
|---|---|---|
| 0 = no positive cells | 0 = no color reaction | 0–1 = negative |
| 1 = <10% positive cells | 1 = mild reaction | 2–3 = mild |
| 2 = 10–50% positive cells | 2 = moderate reaction | 4–8 = moderate |
| 3 = 51–80% positive cells | 3 = intense reaction | 9–12 = strongly positive |
| 4 = >80% positive cells | Final IRS score (A × B): 0–12 * | |
| Immunomarker | Percentage of Positive Cells | Intensity of Immunostaining | IRS Score | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 0 | 1 | 2 | 3 | 0–1 | 2–3 | 4–8 | 9–12 | |
| E-cadherin (n = 6) | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 6 |
| COX-2 (n = 6) | 0 | 0 | 1 | 4 | 1 | 0 | 1 | 4 | 1 | 0 | 0 | 6 | 0 |
| Pan-CK (n = 6) | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solanes, F.; Chiers, K.; Kik, M.J.L.; Hellebuyck, T. Gross, Histologic and Immunohistochemical Characteristics of Keratoacanthomas in Lizards. Animals 2023, 13, 398. https://doi.org/10.3390/ani13030398
Solanes F, Chiers K, Kik MJL, Hellebuyck T. Gross, Histologic and Immunohistochemical Characteristics of Keratoacanthomas in Lizards. Animals. 2023; 13(3):398. https://doi.org/10.3390/ani13030398
Chicago/Turabian StyleSolanes, Ferran, Koen Chiers, Marja J. L. Kik, and Tom Hellebuyck. 2023. "Gross, Histologic and Immunohistochemical Characteristics of Keratoacanthomas in Lizards" Animals 13, no. 3: 398. https://doi.org/10.3390/ani13030398