1. Introduction
The term “organic production” refers to a farming and food-production approach that incorporates optimal environmental practices, promotes extensive biodiversity, conserves natural resources, upholds stringent standards for animal welfare, and aligns with the preferences of consumers seeking products created using environmentally friendly methods [
1]. Organic aquaculture is a modest but growing part of the global food-supply chain [
2]. Its production methods [
3] were adopted due to the increasing interest in sustainable resource utilisation [
4,
5]. Organic aquaculture can be carried out using various technologies, including recirculating aquaculture systems (RAS), net pens, cages, raceways, and tanks [
6]. The appropriate fish stocking density in RAS qualifies these systems for use in organic production, providing safe conditions for most fish in terms of animal welfare and biosecurity concerns [
7]. Nevertheless, transitioning from conventional to organic aquaculture involves an intricate and multifaceted process, encompassing considerations related to consumer safety, ecological and environmental impacts, socioeconomic factors, and animal welfare [
8,
9]. The argument over the use of organic feeds for organic aquaculture is still ongoing because a balance must be struck between the realities of the supply of sources for aquafeeds and the fundamental principles of organic food production. Additionally, feeds must support animal health and growth, provide a final edible product of excellent quality, have a low impact on the environment, and be balanced to meet the nutritional needs of the farmed species [
10].
In 2020, the EU27’s total organic aquaculture production was approximated at 74,032 tonnes, constituting 6.4% of the EU’s total output. This production marks a 60% increase from 2015 (46,341 tonnes at the EU 27 level in 2015), primarily due to increased production of organic mussels [
11]. One of the main species produced organically in the Mediterranean is European seabass, the production of which increased from 2000 tonnes in 2015 to 2750 tonnes in 2020. Greece is the leading EU producer of this fish [
11]. Economic considerations such as increased production expenses and increased retail costs have dissuaded both farmers and consumers, limiting the growth of organic production of seabass [
12]. Research on consumer preferences shows that organic seabass production in the Mediterranean is economically promising [
13,
14], but appropriate marketing strategies still need to be developed [
15,
16]. Furthermore, a major obstacle to expanding organic aquaculture production is the lack of organic feeds, particularly for carnivorous species. The limitations imposed by the EU organic regulations make it difficult to find organic feed ingredients that are rich in protein and thus to design well-balanced organic diets [
4,
17,
18,
19].
More research is required on the organic cultivation of seabass. Previous studies reported that organic fish show improved growth performance, lower feed-conversion ratios, and increased metabolic rates compared with fish grown in conventional aquaculture [
18]. However, other studies have found no differences in stress and immunological indices between fish grown in organic and conventional aquaculture [
18,
20].
The general principles of ecological production, such as the development of processes that are based on environmental systems and that use the system’s natural resources, the restricted use of synthetic substances, and the limited use or non-use of genetically modified organisms (GMOs), apply to organic aquaculture, with some additional limitations regarding the availability of organic resources [
21]. The relevant regulatory limitations stipulate a maximum of 60% organic plant ingredients and the absence of synthetic amino acids [
22]. Continual efforts are required to identify alternative sources of nutritional protein and lipids for organic feeds in organic aquaculture, with a focus on minimising the utilisation of fishmeal (FM) and fish oil in such feeds. However, there is a need to focus on the quality and certification of alternative ingredients for use in organic aquaculture. Research is still being done to investigate novel alternative formulations of ingredients and the quality of the resulting products [
1].
Many researchers have studied the effects of substituting FM with plant-based proteins [
23,
24,
25,
26,
27,
28]. Completely replacing animal proteins with plant proteins has generally not been successful due to concerns about antinutrients, changes in amino-acid absorption, potential micronutrient deficiencies, and immune suppression [
29,
30,
31]. Other potential feed sources, excluding plant proteins, include microbiological organisms (bacteria, microalgae, fungi), byproducts from terrestrial animals (processed animal protein (PAP), blood meal), annelid worms obtained from wild harvesting and cultivation, and the larvae and pupae of insects [
32,
33,
34]. The utilisation of animal byproducts is made possible by PAP, a key ingredient in feeds [
35]. According to several studies, insects can be used as a source of protein for fish [
36,
37,
38,
39]. European seabass can be fed insect meal from
Tenebrio molitor at varying concentrations without adversely affecting growth performance, according to a feeding assay [
40]. Byproduct meals can be highly appealing due to their competitive pricing compared to fish meal, making them a potentially interesting and cost-effective option [
41].
Due to regulatory restrictions, it is difficult to find enough organic protein sources suitable for seabass, one of the main carnivorous fish produced in Europe. Transformed animal proteins (TAPs) from non-ruminant animals, whose use is permitted in conventional aquaculture (RD 578/2014), as well as insects (Regulation EU 893/2017), are suggested. The use of organic-derived TAPs in organic aquaculture does not violate any regulations and facilitates the formulation of organic aquaculture feed without the need for captured FM, relying solely on the recovery of byproducts from organic aquaculture.
On the other hand, there has been considerable interest in the use of in vitro assays to evaluate the digestibility of a prospective feed product for aquatic species, such as fish, prawns, and molluscs [
42]. The in vitro digestibility test is appropriate for preliminary research. It allows many samples to be analysed because it is inexpensive, has no ethical restrictions, and is reasonably simple to carry out [
43]. It also allows the conduction of controlled experiments to investigate how proteins, lipids, and carbohydrates in feed items are hydrolysed [
42]. Research on fish digestion in vitro is still in its infancy, based on the number of relevant publications.
The present work aimed to provide a 100% organic diet for seabass, one of Europe’s most important marine aquaculture species, using alternative organic raw materials such as insects, byproducts from Iberian pigs, and rainbow trout remains. This research may support a dramatic improvement in organic aquaculture.
2. Materials and Methods
2.1. System for Rearing
The trial was conducted in 18 cylindrical fiberglass tanks, each with a volume of 1750 L, as part of a saltwater recirculating system with a total capacity of 75 m3. The system was equipped with a rotating mechanical filter, a gravity biofilter with a capacity of 6 m3, and a skim (September to January). A heat pump was used to ensure that the water temperature remained constant (20.9 °C), and all tanks had aeration. The dissolved oxygen level was 7.7 mg L−1, and salinity was 31.3 g L−1. The pH was maintained at 8.0, with nitrates (NO−3) at a concentration of 33.2 mg L−1, nitrites (NO−2) at 0.13 mg L−1, and ammonium (NH+4) at 0.03 mg L−1. The photoperiod was natural (11 h), and the lighting was the same in all tanks.
2.2. Fish
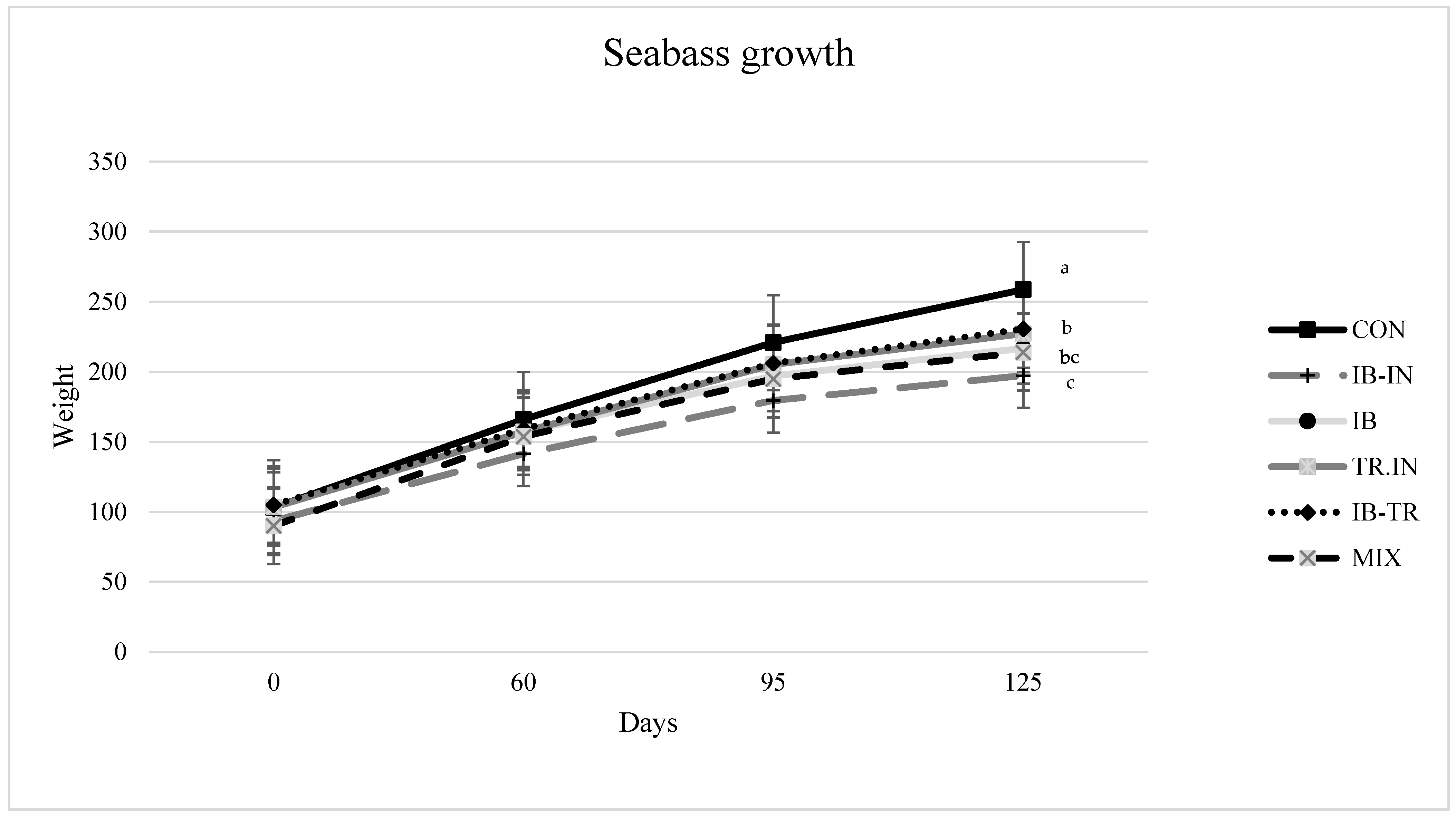
Juvenile organic seabass from the fish farm Sonrionansa S.L. situated in Pesues (Cantabria, Spain) were delivered to the Universitat Politècnica de València and distributed among experimental tanks. Before the feeding experiment, a 15-day acclimatisation period was provided to allow all fish to adapt to the laboratory conditions. There were 486 fish, with an average weight of 90 g, distributed throughout the 18 test tanks (27 fish per tank). The experiment was carried out over 125 days.
2.3. Diets and Feeding
The proximal composition of the raw materials is shown in
Table 1. Six diets were tested in triplicate: (1) a control diet containing FM provided for sustainable fisheries as a protein source (CON); (2) a diet in which the protein source was composed of Iberian pig meal byproduct (diet IB); (3) a diet containing organic Iberian pig meal byproduct and organic insect meal (diet IB-IN); (4) a diet based on organic Iberian pig and organic rainbow-trout byproduct meal (diet IB-TR); (5) a diet with organic rainbow trout byproducts and organic insect meal as protein sources (diet TR-IN); and (6) a MIX diet containing organic insect meal, organic rainbow trout meal and organic Iberian pig byproduct meal (
Table 2). The nutritional compositions of all the diets are represented as the means of five separate analyses conducted during each feed-manufacturing cycle. Formulations were initially designed with different amounts of raw ingredients to maintain a consistent composition of 45% crude protein (CP) and 20% crude lipid (CL); challenges in effectively mixing certain ingredients led to variations in some of these values (
Table 2).
All diets were produced at the Universitat Politècnica de València using a semi-industrial twin-screw extruder (CLEXTRAL BC-45, Firminy, St Etienne, France), under the following processing conditions: a screw speed set at 100 rpm, pressure ranging from 40 to 50 atm, and a temperature maintained at 110 °C. Calcium phosphate and organic vegetable amino acids were included in diets as supplements (lysine and methionine). Diet formulation and manufacture were carried out with organic raw materials labelled and approved by Regulation (EU) 2018/848.
Each experimental diet was evaluated in three randomly distributed tanks. The fish were manually fed twice a day at 9:00 and 17:00, six days per week (from Monday to Saturday).
The fish were fed until they reached satiety, and the pellets were administered gradually. Daily monitoring and weighing occurred every 30 days before anaesthetising the fish were anaesthetised with clove oil, which contained 87% eugenol (Guinama®, Valencia, Spain), at a concentration of 10 mg L−1 of water.
The aim of this process was to evaluate the growth of the fish throughout the experiment, define growth parameters, and assess the overall health of the fish. The fish were starved the day before they were weighed. Ten fish were collected at the start of the experiment and preserved at −30 °C for subsequent analysis of their body composition. Three specimens from each tank were randomly selected for sampling and pooling to determine the approximate composition and amino acid content of their bodies.
2.4. Analysis of Nutritional Composition and Amino Acids
The diets and their approximate composition (
Table 2), as well as the whole fish, were examined using the methods described in [
44] and analysed for the following metrics: dry matter (105 °C to constant weight); ash (incinerated at 550 °C for five hours); crude protein (determined by the direct combustion method DUMAS using LECO CN628, Geleen, The Netherlands); and crude lipid (extracted with methyl-ether using ANKOMXT10 Extractor (Macedon, NY, USA)). Each analysis was carried out in triplicate.
A Waters HPLC system (Waters 474, Waters, Milford, MA, USA) composed of two pumps (Model 515, Waters), an autosampler (Model 717, Waters), a fluorescence detector (Model 474, Waters), and a temperature-control module was used to analyse the levels of amino acids (AA) in the diets and in the fish using the procedure previously described by Bosch et al., 2006 [
45].
Before hydrolyzation, aminobutyric acid was introduced as a internal standard. AQC was used to derivatise AA (6-aminoquinolyl-N-hydroxysuccinimidyl carbamate). After oxidation with performic acid, methionine and cysteine were identified individually as methionine sulphone and cystic acid. AA was converted to methionine and cystine after it was separated with a reverse-phase C-18 column by Waters Acc—Tag (150 mm 3.9 mm).
Table 3 shows the essential amino acids (EAA) content of the experimental diets. All amino acid analyses were carried out in duplicate.
2.5. Indices of Growth
At the end of the trial, the nutrient efficiency indices and growth were determined. Metrics included the survival rate (SR), specific growth rate (SGR), feed intake (FI), feed conversion ratio (FCR), and protein efficiency ratio (PER), considering each tank as an experimental unit. All fish were weighed. Additionally, the productive protein value (PPV%) and productive fat value (PFV%) were calculated. These parameters were calculated using the following equations:
2.6. Digestibility Assay
The digestibility test was performed after the growth experiment ended and was carried out in triplicate, in three tanks. Fifteen seabass were randomly placed in each experimental tank (190 L fibreglass tank, 88 cm high, 62 cm wide, and 188 cm deep) in a semi-closed recirculating system based on the Guelph system (the faecal material being collected in a settling column). The water flow velocity was altered to reduce the settling of faeces in the drainpipe and increase the recovery of faeces in the settling column.
The fish received one meal per day at 10:00 a.m. The diet was offered so as to reduce waste while the fish were actively feeding. The drainpipe and the settling column were dusted an hour after feeding to prevent faeces from being contaminated by column diets. The faeces were gravity-collected in a plastic container from the base of the settling column 6–7 h after feeding.
After collection, the faeces were weighed and dried in a 60 °C oven for 48 h before analysis. Subsequently, they were preserved in sealed plastic containers and analysed for nutritional components and inert markers. Chromic oxide (Cr2O3) was used (5 g kg−1) as an inert and indigestible marker. An atomic absorption spectrometer was used to determine the amount of chromium oxide in diets and faeces after acid digestion (Perkin Elmer 3300, Perkin Elmer, Boston, MA, USA). Additionally, analyses were conducted for crude protein, dry matter, calcium, energy, and phosphorus in diets and faces. All analyses were performed in triplicate.
The apparent digestibility coefficients of the diet (ADC) were determined using [
46]. The ADCs of the dry matter (ADCdm, %) of the diets were determined per Equation (1):
The percentage of ADCs for each dietary nutrient (protein, energy, calcium, and phosphorus) was calculated using Equation (2):
The variables “nutrient diet (g kg−1)” and “nutrient faeces (g kg−1)” in this equation indicate the amounts of a nutrient (such as protein or energy) in the diet and the faeces, respectively. The measurements “marker diet” (g kg−1) and “marker faeces” (g kg−1) indicate the amount of marker in the diet and the faeces, respectively.
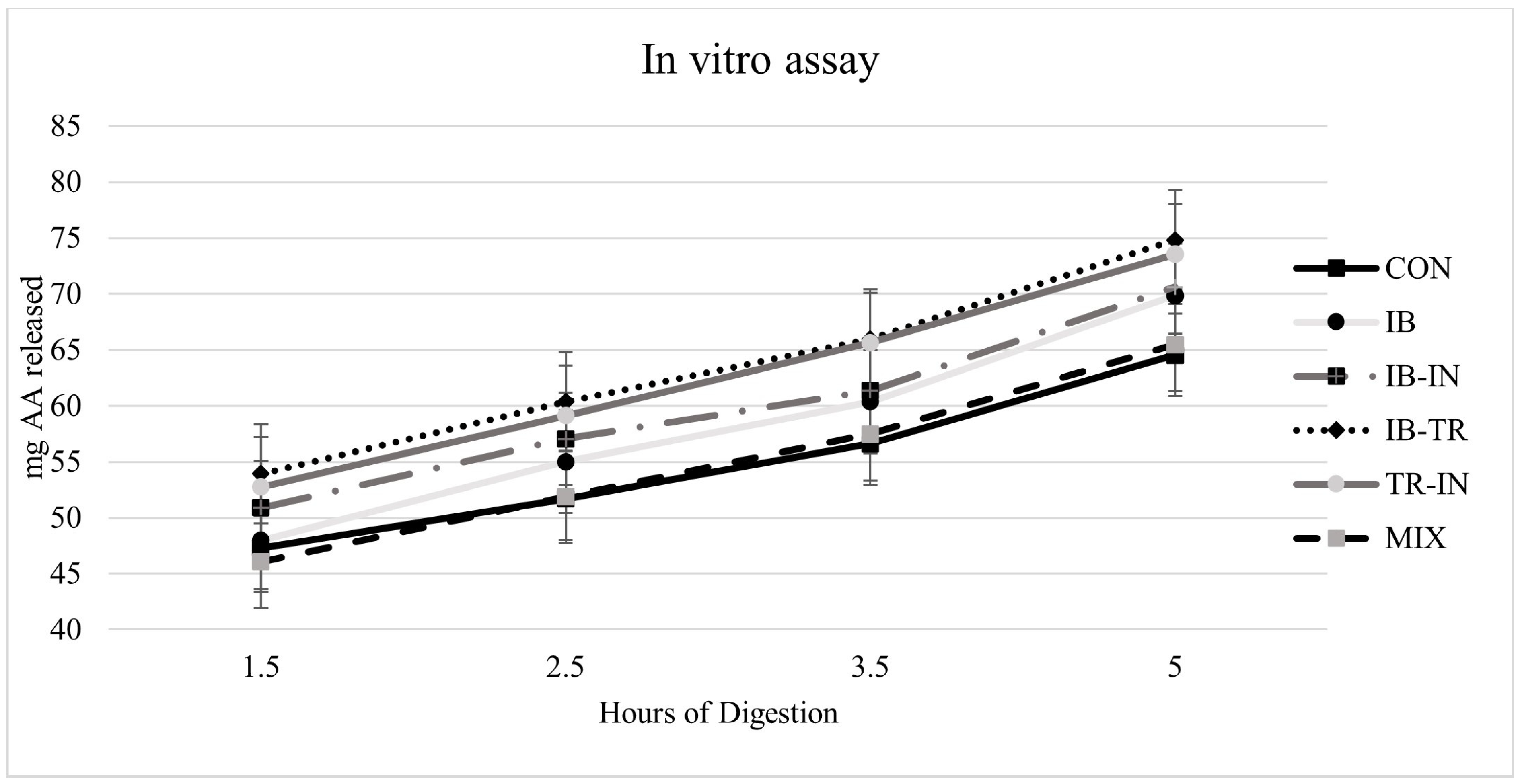
2.7. In Vitro Hydrolysis Assay
The in vitro hydrolysis trial was conducted under conditions that simulated the digestive system of juvenile European seabass [
47]). Ten juvenile seabasses with an average weight of 100 g were used and sampled six hours after feeding to ensure the presence of enzymes in both the stomach and the intestine. The fish were euthanised in ice-cold water with a small quantity of clove oil, which acted as an anaesthetic. Subsequently, the fish were immediately dissected to extract the digestive tract. The digestive tract was divided into two parts: (1) the proximal intestine, which encompassed the diffuse pancreas and pyloric caecum, and (2) the stomach. These tissues were utilised to create extracts for measuring protease activity. The methods used were as follows: acid protease was measured by tyrosine release from haemoglobin hydrolysis at pH 2.5 [
48]; alkaline protease was measured by tyrosine release from casein at pH 8.5 [
49]; and amylase was measured during the preliminary evaluation of the enzymes of the juvenile European seabass at pH 7.5 [
50]. The extracts were prepared by mechanical homogenisation of the tissues in distilled water (1:10
w/
v) and centrifugation (3220×
g, 20 min, 4 °C). The supernatant was then filtered through a dialysis system with a MWCO of 10 kDa (Pellicon XL, Millipore, Burlington, MA, USA), and the concentrated extracts were freeze-dried until they were required for the assays. The activities of acid protease in the stomach (pepsin) and total intestinal alkaline proteases present in the extracts were measured using the methods described in refs. [
47,
48]. Protease activity levels were used as indicators to estimate the amount of extracts required to provide physiological enzyme-substrate ratios in the assays. These ratios were calculated considering, on one hand, the average total production of enzyme measured in several fish in relation to their live weight, and on the other, the average intake per meal of fish of such a size, a value obtained from commercial ration tables.
Based on these findings, the average enzyme production was estimated as follows: acid protease, 37.7 U g
−1 weight; and alkaline protease, 24.7 U g
−1 weight. The conditions are given in
Table 4.
Three assay replicates were conducted for each diet, and a blank sample was also included. The blank sample was created by deactivating the enzymatic extracts through heat treatment before they were added to the bioreactors. This step enabled measurement of the amino acids in the extracts and the diet.
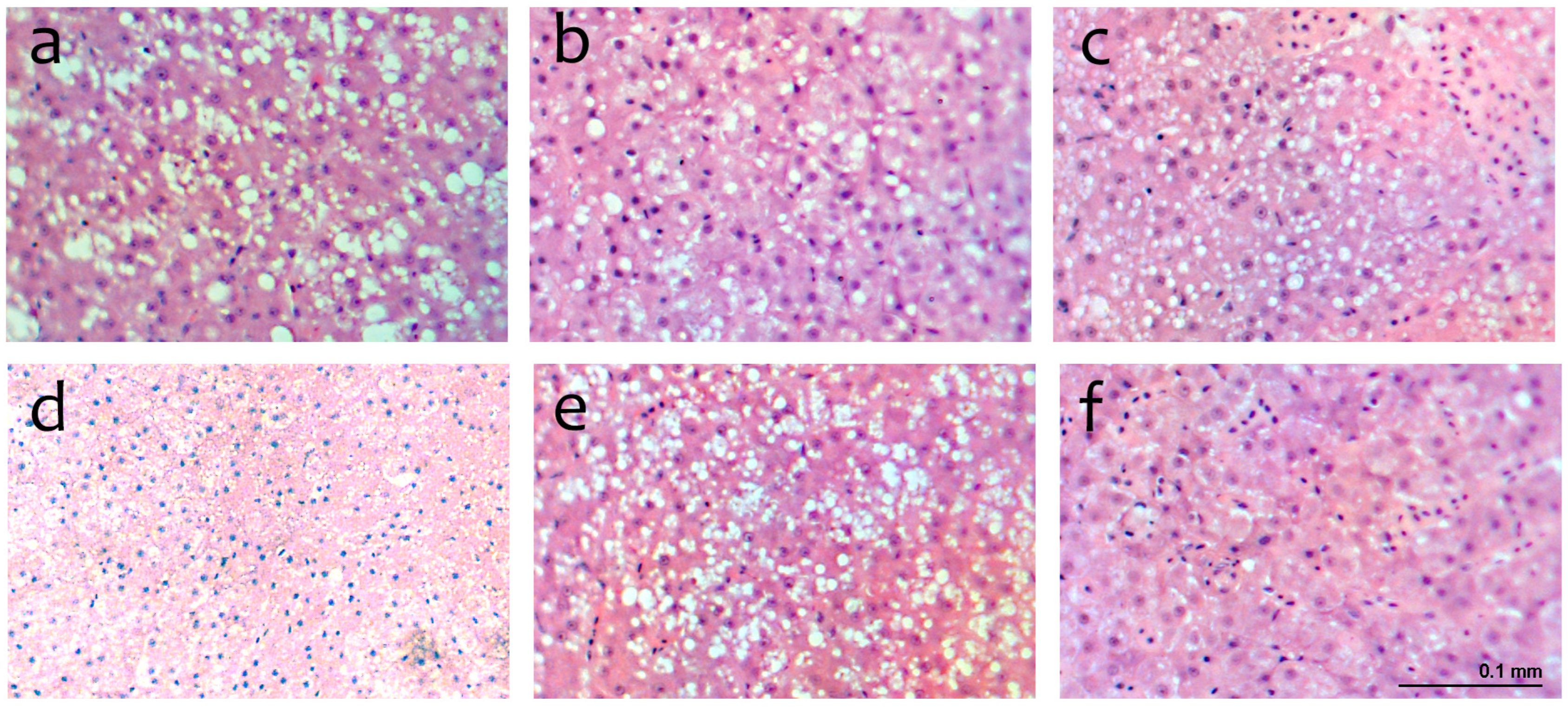
2.8. Histological Analysis of the Liver
The liver was collected from was three fish per tank after the growth experiment ended. Samples were preserved in phosphate-buffered formalin (4%, pH 7.4). According to typical histological procedures, all the formalin-fixed tissues underwent regular dehydration in ethanol, were conditioned in ultra-clean environments, and were embedded in paraffin. Transverse sections from each paraffin block were taken using a Shandon Hypercut microtome, then stained for haematoxylin and eosin analysis.
One hundred sections of the liver were examined using an Eclipse E400 Nikon light microscope from Izasa S.A. in Barcelona, Spain. To determine the effects of different feeds on the liver, the diameters of hepatocytes and nuclei were measured [
51,
52].
2.9. Statistical Analysis
All data were checked for normal distribution and homogeneity of variances. Using the Statgraphics® Plus 5.1 statistical programme (Statistical Graphics Corp, Rockville, MO, USA), various growth and nutrient indices, retention of AA, ADC, in vitro hydrolysis, and histological measurements were analysed using analysis of variance with a Newman-Keul test for multiple comparisons. The initial covariate weight was used to analyse growth indices. The findings are represented as means with standard error (SEM, standard error of the mean). The significance level was established at p < 0.05.


 ,
, 




{kind=link}
{kind=link}
{kind=link}