
In Search of Suitable Breeding Sites: Habitat Heterogeneity and Environmental Filters Determine Anuran Diversity of Western Madagascar

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Data Acquisition
2.2.1. The Presence of Anuran Species at Breeding Ponds
2.2.2. Environmental Variables of Breeding Ponds
2.3. Statistical Analysis
2.3.1. Influence of Environmental Variables on Species Richness
2.3.2. The Influence of Environmental Variables on Assemblage Composition
3. Results
3.1. Comparison of Forest and Riverbed Ponds
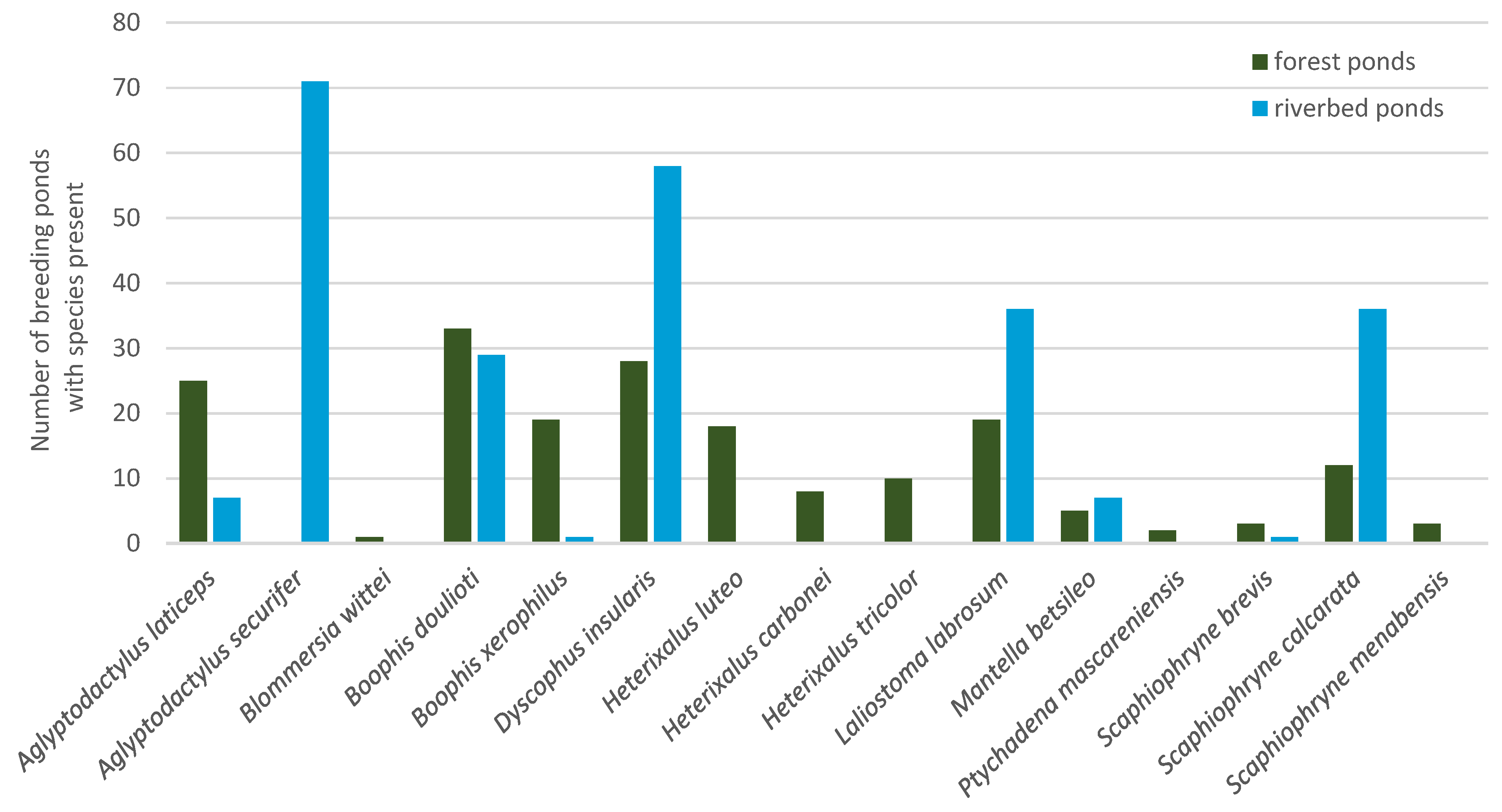
3.2. Distribution of Tadpole Species
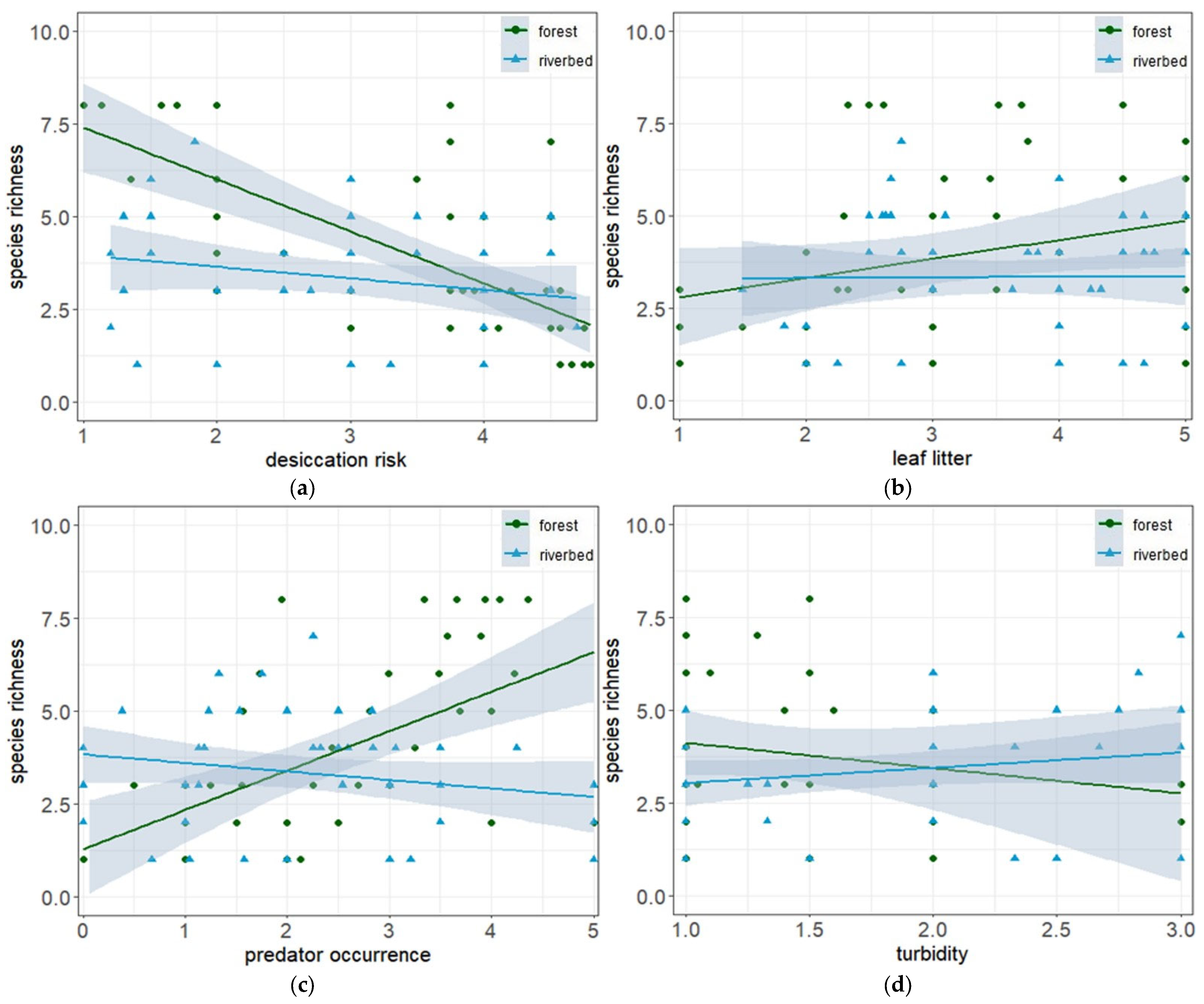
3.3. Environmental Variability and Species Richness
3.4. Species Composition in Different Habitat Types
4. Discussion
5. Conservation Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Araújo, M.B.; Williams, P.H.; Fuller, R.J. Dynamics of extinction and the selection of nature reserves. Proc. R. Soc. Biol. Sci. 2002, 269, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Andreone, F.; Cadle, J.E.; Cox, N.; Glaw, F.; Nussbaum, R.A.; Raxworthy, C.J.; Stuart, S.N.; Vallan, D.; Vences, M. Species review of amphibian extinction risks in Madagascar: Conclusions from the Global Amphibian Assessment. Conserv. Biol. 2005, 19, 1790–1802. [Google Scholar] [CrossRef]
- Glos, J.; Volahy, A.T.; Bourou, R.; Straka, J.; Young, R.; Durbin, J. Amphibian conservation in central Menabe. A Conserv. Strategy Amphib. Madag. 2008, 48, 107–124. [Google Scholar]
- Sodhi, N.S.; Bickford, D.; Diesmos, A.C.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Sekercioglu, C.H.; Bradshaw, C.J.A. Measuring the meltdown: Drivers of global amphibian extinction and decline. PLoS ONE 2008, 3, e1636. [Google Scholar] [CrossRef] [PubMed]
- Bishop, P.J.; Angulo, A.; Lewis, J.P.; Moore, R.D.; Rabb, G.B.; Garcia Moreno, J. The Amphibian Extinction Crisis—What will it take to put the action into the Amphibian Conservation Action Plan? Surv. Perspect. Integr. Environ. Soc. 2012, 5, 97–111. [Google Scholar]
- Jorge, R.F.; Simões, P.I.; Magnusson, W.E.; Lima, A.P. Fine-scale habitat heterogeneity explains the local distribution of two Amazonian frog species of concern for conservation. Biotropica 2016, 48, 694–703. [Google Scholar] [CrossRef]
- Della Bella, V.; Bazzanti, M.; Chiarotti, F. Macroinvertebrate diversity and conservation status of Mediterranean ponds in Italy: Water permanence and mesohabitat influence. Aquat. Conserv. Mar. Freshw. Ecosyst. 2005, 15, 583–600. [Google Scholar] [CrossRef]
- Davies, B.; Biggs, J.; Williams, P.; Whitfield, M.; Nicolet, P.; Sear, D.; Bray, S.; Maund, S. Comparative biodiversity of aquatic habitats in the European agricultural landscape. Agric. Ecosyst. Environ. 2008, 125, 1–8. [Google Scholar] [CrossRef]
- Silver, C.A.; Vamosi, S.M.; Bayley, S.E. Temporary and permanent wetland macroinvertebrate communities: Phylogenetic structure through time. Acta Oecologica 2012, 39, 1–10. [Google Scholar] [CrossRef]
- Keller, A.; Rödel, M.-O.; Linsenmair, K.E.; Grafe, T.U. The importance of environmental heterogeneity for species diversity and assemblage structure in Bornean stream frogs. J. Anim. Ecol. 2009, 78, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Von May, R.; Jacobs, J.M.; Santa-Cruz, R.; Valdivia, J.; Huamán, J.M.; Donnelly, M.A. Amphibian community structure as a function of forest type in Amazonian Peru. J. Trop. Ecol. 2010, 26, 509–519. [Google Scholar] [CrossRef]
- Badgley, C.; Fox, D.L. Ecological biogeography of North American mammals: Species density and ecological structure in relation to environmental gradients. J. Biogeogr. 2000, 27, 1437–1467. [Google Scholar] [CrossRef]
- Werner, E.E.; Skelly, D.K.; Relyea, R.A.; Yurewicz, K.L. Amphibian species richness across environmental gradients. Oikos 2007, 116, 1697–1712. [Google Scholar] [CrossRef]
- Welsh, H.H.; Ollivier, L.M. Stream amphibians as indicators of ecosystem stress: A case study from California’s Redwoods. Ecol. Appl. 1998, 8, 1118. [Google Scholar] [CrossRef]
- Vormisto, J.; Phillips, O.L.; Ruokolainen, K.; Tuomisto, H.; Vásquez, R. A comparison of fine-scale distribution patterns of four plant groups in an Amazonian rainforest. Ecography 2000, 23, 349–359. [Google Scholar] [CrossRef]
- Browne, C.L.; Paszkowski, C.A.; Foote, A.L.; Moenting, A.; Boss, S.M. The relationship of amphibian abundance to habitat features across spatial scales in the Boreal Plains. Écoscience 2009, 16, 209–223. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Burne, M.R.; Griffin, C.R. Habitat associations of pool-breeding amphibians in eastern Massachusetts, USA. Wetl. Ecol. Manag. 2005, 13, 247–259. [Google Scholar] [CrossRef]
- Afonso, L.G.; Eterovick, P.C. Microhabitat choice and differential use by anurans in forest streams in southeastern Brazil. J. Nat. Hist. 2007, 41, 937–948. [Google Scholar] [CrossRef]
- Maseko, M.S.; Zungu, M.M.; Ehlers Smith, D.A.; Ehlers Smith, Y.C.; Downs, C.T. High microhabitat heterogeneity drives high functional traits in forest birds in five protected forest areas in the urban mosaic of Durban, South Africa. Glob. Ecol. Conserv. 2019, 18, e00645. [Google Scholar] [CrossRef]
- Pawar, S.; Koo, M.S.; Kelley, C.; Ahmed, M.F.; Chaudhuri, S.; Sarkar, S. Conservation assessment and prioritization of areas in Northeast India: Priorities for amphibians and reptiles. Biol. Conserv. 2007, 136, 346–361. [Google Scholar] [CrossRef]
- Glos, J.; Dausmann, K.H.; Linsenmair, E.K. Modeling the habitat use of Aglyptodactylus laticeps, an endangered dry-forest frog from Western Madagascar. A Conserv. Strategy Amphib. Madag. 2008, 45, 125–142. [Google Scholar]
- Baldwin, R.F.; de Maynadier, P.G. Assessing threats to pool-breeding amphibian habitat in an urbanizing landscape. Biol. Conserv. 2009, 142, 1628–1638. [Google Scholar] [CrossRef]
- Ochoa-Ochoa, L.; Urbina-Cardona, J.N.; Vázquez, L.-B.; Flores-Villela, O.; Bezaury-Creel, J. The effects of governmental protected areas and social initiatives for land protection on the conservation of Mexican amphibians. PLoS ONE 2009, 4, e6878. [Google Scholar] [CrossRef] [PubMed]
- D’Amen, M.; Bombi, P.; Pearman, P.B.; Schmatz, D.R.; Zimmermann, N.E.; Bologna, M.A. Will climate change reduce the efficacy of protected areas for amphibian conservation in Italy? Biol. Conserv. 2011, 144, 989–997. [Google Scholar] [CrossRef]
- de Pous, P.; Beukema, W.; Weterings, M.; Dümmer, I.; Geniez, P. Area prioritization and performance evaluation of the conservation area network for the Moroccan herpetofauna: A preliminary assessment. Biodivers. Conserv. 2011, 20, 89–118. [Google Scholar] [CrossRef]
- Glos, J.; Erdmann, G.; Dausmann, K.H.; Linsenmair, K.E. A comparative study of predator-induced social aggregation of tadpoles in two anuran species from western Madagascar. Herpetol. J. 2007, 17, 261–268. [Google Scholar]
- Ganzhorn, J.U.; Sorg, J.-P. Ecology and economy of a tropical dry forest in Madagascar. Primate Rep. 1996, 46. [Google Scholar]
- Nelson, R.; Horning, N. AVHRR-LAC estimates of forest area in Madagascar, 1990. Int. J. Remote Sens. 1993, 14, 1463–1475. [Google Scholar] [CrossRef]
- Glos, J. The amphibian fauna of the Kirindy dry forest in western Madagascar. Salamandra Rheinb. 2003, 39, 75–90. [Google Scholar]
- Heyer, W.R.; Donnelly, M.A.; McDiarmid, R.W.; Hayek, L.-A.; Foster, M.S. Measuring and Monitoring Biological diversity: Standard Methods for Amphibians; Smithsonian Institution Press: Washington, DC, USA, 1994; pp. 57–60. [Google Scholar]
- Glos, J.; Linsenmair, K.E. Descriptions of the tadpoles of Aglyptodactylus laticeps and Aglyptodactylus securifer from Western Madagascar, with notes on life history and ecology. J. Herpetol. 2004, 38, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Glaw, F.; Vences, M. A Fieldguide to the Amphibians and Reptiles of Madagascar; Vences, M., Glaw, F., Eds.; Verlags GbR.: München, Germany, 2007. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 3 March 2023).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-PLUS; Springer: Berlin/Heidelberg, Germany, 2002; Volume 4. [Google Scholar]
- Simpson, G.L. Permute: Functions for Generating Restricted Permutations of Data. 2022. Available online: https://CRAN.R-project.org/package=permute (accessed on 26 February 2023).
- Sarkar, D. Lattice: Multivariate Data Visualization with R; Springer: New York, NY, USA, 2008; Available online: http://lmdvr.r-forge.r-project.org (accessed on 8 March 2023).
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conversat. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Crawley, M.J. Generalized Linear Models. In The Oxford Handbook of Quantitative Methods; Oxford University Press: New York, NY, USA, 2013; pp. 557–578. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Clarke, K.R. Nonmetric multivariate analysis in community-level ecotoxicology. Environ. Toxicol. Chem. 1999, 18, 118. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 4, 325–339. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L.F.J. Numerical Ecology: Developments in environmental modelling. Elsevier Sci. Publ. Co. 1998, 24, 201–205. [Google Scholar]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- Vences, M.; Wollenberg, K.C.; Vieites, D.R.; Lees, D.C. Madagascar as a model region of species diversification. Trends Ecol. Evol. 2009, 24, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D. Flagships, umbrellas, and keystones—Is single-species management passe in the landscape era? Cireat Br. 1998, 83, 247–257. [Google Scholar] [CrossRef]
- Vasconcelos, T.S.; Rossa-Feres, D.C. Diversidade, distribuição espacial e temporal de anfíbios anuros (Amphibia, Anura) na região noroeste do estado de São Paulo, Brasil. Biota Neotrop. 2005, 5, 137–150. [Google Scholar] [CrossRef]
- Bastazini, C.V.; Munduruca, J.F.V.; Rocha Pedro Luís, B.; Napoli Marcelo, F. Which environmental variables better explain changes in anuran community composition? A case study in the Restinga of Mata de São João, Bahia, Brazil. Herpetologica 2007, 63, 459–471. [Google Scholar] [CrossRef]
- Piatti, L.; Souza, F.L.; Filho, P.L. Anuran assemblage in a rice field agroecosystem in the Pantanal of central Brazil. J. Nat. Hist. 2010, 44, 1215–1224. [Google Scholar] [CrossRef]
- Parris, K.M.; McCarthy, M.A. What influences the structure of frog assemblages at forest streams? Austral Ecol. 1999, 24, 495–502. [Google Scholar] [CrossRef]
- Eterovick, P.C. Distribution of anuran species among montane streams in south-eastern Brazil. J. Trop. Ecol. 2003, 19, 219–228. [Google Scholar] [CrossRef]
- Vasconcelos, T.S.; Santos, T.G.; Rossa-Feres, D.C.; Haddad, C. Influence of the environmental heterogeneity of breeding ponds on anuran assemblages from southeastern Brazil. Can. J. Zool. 2009, 87, 699–707. [Google Scholar] [CrossRef]
- Parris, K.M. Environmental and spatial variables influence the composition of frog assemblages in sub-tropical eastern Australia. Ecography 2004, 27, 392–400. [Google Scholar] [CrossRef]
- Julian Glos, K. Eduard Linsenmair. Description of the tadpoles of Boophis doulioti and B. xerophilus from Western Madagascar with notes on larval life history and breeding ecology. Amphib. Reptil. 2005, 26, 459–466. [Google Scholar] [CrossRef]
- Glos, J.; Wegner, F.; Dausmann, K.H.; Linsenmair, K.E. Oviposition-site selection in an endangered Madagascan frog: Experimental evaluation of a habitat model and its implications for conservation. Biotropica 2008, 40, 646–652. [Google Scholar] [CrossRef]
- Hartel, T.; Bancila, R.; Cogalniceanu, D.A. Spatial and temporal variability of aquatic habitat use by amphibians in a hydrologically modified landscape. Freshw. Biol. 2011, 56, 2288–2298. [Google Scholar] [CrossRef]
- Glos, J.; Metzner, A.; Schmidtke, C.; Rödel, M.-O. The mating system of the reed frog Heterixalus tricolor (Anura: Hyperoliidae) from western Madagascar’s dry forests. Salamandra 2019, 55, 151–159. [Google Scholar]
- Snodgrass, J.W.; Komoroski, M.J.; Bryan, A.L.; Burger, J. Relationships among isolated wetland size, hydroperiod, and amphibian species richness: Implications for wetland regulations. Conserv. Biol. 2000, 14, 414–419. [Google Scholar] [CrossRef]
- Paton, P.W.C.; Crouch, W.B., III. Using the phenology of pond-breeding amphibians to develop conservation strategies. Conserv. Biol. 2002, 16, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Babbitt, K.J.; Baber, M.J.; Tarr, T.L. Patterns of larval amphibian distribution along a wetland hydroperiod gradient. Can. J. Zool. 2003, 81, 1539–1552. [Google Scholar] [CrossRef]
- Egan, R.S.; Paton, P.W.C. Within-pond parameters affecting oviposition by wood frogs and spotted salamanders. Wetlands 2004, 24, 1–13. [Google Scholar] [CrossRef]
- Wellborn, G.A.; Skelly, D.K.; Werner, E.E. Mechanisms creating community structure across a freshwater habitat gradient. Annu. Rev. Ecol. Syst. 1996, 27, 337–363. [Google Scholar] [CrossRef]
- Weyrauch, S.L.; Grubb, T.C., Jr. Patch and landscape characteristics associated with the distribution of woodland amphibians in an agricultural fragmented landscape: An information-theorie approach. Biol. Conserv. 2004, 115, 443–450. [Google Scholar] [CrossRef]
- Kats, L.B.; Petranka, J.W.; Sih, A. Antipredator defenses and the persistence of amphibian larvae with fishes. Ecology 1988, 69, 1865–1870. [Google Scholar] [CrossRef]
- Hero, J.-M.; Magnusson, W.E.; Rocha, C.F.D.; Catterall, C.P. Antipredator defenses influence the distribution of amphibian prey species in the Central Amazon Rain Forest. Biotropica 2001, 33, 131. [Google Scholar] [CrossRef]
- Morin, P.J. Predation, competition, and the composition of larval anuran guilds. Ecol. Monogr. 1983, 53, 119–138. [Google Scholar] [CrossRef]
- Wilbur, H.M. Experimental Ecology of food webs: Complex systems in temporary ponds. Ecology 1997, 78, 2279–2302. [Google Scholar] [CrossRef]
- Lawton, J.H. Plant Architecture and the Diversity of Phytophagous Insects. Annu. Rev. Entomol. 1983, 28, 23–39. [Google Scholar] [CrossRef]
- Haddad, C.F.B.; Prado, C.P.A. Reproductive modes in frogs and their unexpected diversity in the Atlantic Forest of Brazil. BioScience 2005, 55, 207. [Google Scholar] [CrossRef]
- Werner, E.E.; Glennemeier, K.S. Influence of forest canopy cover on the breeding pond distributions of several amphibian species. Copeia 1999, 1–12. [Google Scholar] [CrossRef]
- Afonso, L.G.; Eterovick, P.C. Spatial and temporal distribution of breeding anurans in streams in southeastern Brazil. J. Nat. Hist. 2007, 41, 949–963. [Google Scholar] [CrossRef]
- Magurran, A.E.; Higham, A. Information transfer across fish shoals under predator threat. Ethology 1988, 78, 153–158. [Google Scholar] [CrossRef]
- Babbitt, K.J.; Tanner, G.W. Effects of cover and predator identity on predation of Hyla squirella tadpoles. J. Herpetol. 1997, 31, 128–130. [Google Scholar] [CrossRef]
- Fouilloux, C.A.; Yovanovich, C.A.M.; Rojas, B. Tadpole responses to environments with limited visibility: What we (don’t) know and perspectives for a sharper future. Front. Ecol. Evol. 2022, 9, 1–8. [Google Scholar] [CrossRef]
- Gregory, R.S. Effect of Turbidity on the Predator Avoidance Behaviour of Juvenile Chinook Salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquat. Sci. 1993, 50, 241–246. [Google Scholar] [CrossRef]
- Wood, S.L.R.; Richardson, J.S. Impact of sediment and nutrient inputs on growth and survival of tadpoles of the Western Toad. Freshw. Biol. 2009, 54, 1120–1134. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Name | Variable Definition | Unit | Method |
|---|---|---|---|
| Desiccation risk | Desiccation intensity of ponds. | Five categories: 1 (not at risk of desiccation = long hydroperiod) to 5 (extreme risk of desiccation = short hydroperiod) | Estimated by the number of days before desiccation, after a complete filling of the pond and a successive period of no rainfall. |
| Pond size | Absolute surface area of pond when maximally water filled. | m2 | Measured using length, width and shape. |
| Maximum depth | Maximum pond depth when maximally water filled. | cm | Measured at deepest point of water body. |
| Shallow water | Relative pond area shallower than 10 cm. | % | Visual estimation. |
| Submerged vegetation | Relative volume of pond filled with submerged water plants. | Six categories: 0 (0%), 1 (1–20%), 2 (21–40%), 3 (41–60%), 4 (61–80%), 5 (81–100%) | Visual estimation. |
| Surrounding vegetation | Density and complexity of vegetation structure ≤ 2 m from pond edge. | Six categories: 0 (no vegetation) to 5 (high density) | Visual estimation. |
| Leaf litter | Relative area of pond bottom covered with dead leaves. | Six categories: 0 (0%), 1 (1–20%), 2 (21–40%), 3 (41–60%), 4 (61–80%), 5 (81–100%) | Visual estimation. |
| Turbidity | Turbidity of the pond water influenced by dissolved and suspensed matters. | Three categories: 1 (clear water), 2 (slightly dull), 3 (very turbid water) | Visual estimation. |
| Predator occurrence | Visual estimation of invertebrate predators, including: dytiscid beetles > 0.5 cm, larval dytiscid beetles > 1 cm, water bugs (Belostomatidae) > 0.5 cm, water scorpions (Nepidae) > 1 cm, larval dragonflies (Anisoptera) > 0.5 cm, larval damselflies (Zygoptera) > 2 cm. | Six categories: 0 (no predators) to 5 (high density; >5 predators per dip net stroke) | Measured using dip-netting and the box-method [32] and averaged; determined to higher taxonomic levels in the field and subsequently released. |
| Variables | Forest Ponds | Riverbed Ponds | Wilcoxon-Test |
|---|---|---|---|
| Desiccation (1 to 5) | 3.5 ± 1.2 | 2.9 ± 0.9 | 2653.5, p = 0.001 * |
| Pond size (m2) | 1185.3 ± 3617.6 | 29.8 ± 62.3 | 3038.5, p < 0.001 * |
| Submerged vegetation (0 to 5) | 1.4 ± 1.2 | 0.1 ± 0.2 | 3594.0, p < 0.001 * |
| Surrounding vegetation (0 to 5) | 4.2 ± 0.9 | 2.4 ± 0.8 | 3630.0, p < 0.001 * |
| Turbidity (1 to 3) | 1.4 ± 0.5 | 1.5 ± 0.7 | 1997.5, p = 1 |
| Predator occurrence (0 to 5) | 2.5 ± 1.3 | 2.1 ± 1.4 | 1202.5, p = 0.25 |
| Leaf litter (0 to 5) | 3.2 ± 1.3 | 3.6 ± 1.0 | 1595.5, p = 0.07 |
| Coefficients | Estimate | Standard Error | z-Value | p-Value |
|---|---|---|---|---|
| Initial generalized linear model (AIC = 362.3) | ||||
| Habitat | −0.31 | 0.11 | −0.28 | 0.78 |
| Desiccation | −0.22 | 0.12 | −1.90 | 0.06 |
| Leaf litter | 0.11 | 1 | 1.11 | 0.27 |
| Predator occurrence | 0.17 | 0.73 | 2.39 | 0.02 * |
| Pond size | −0.38 | 0.23 | −0.16 | 0.87 |
| Submerged vegetation | 0.65 | 1 | 0.70 | 0.51 |
| Surrounding vegetation | −0.33 | 0.15 | −0.22 | 0.83 |
| Turbidity | 1 | 0.19 | 0.52 | 0.60 |
| Habitat:Desiccation | 0.6 | 0.15 | 0.39 | 0.7 |
| Habitat:Leaf litter | 0.95 | 0.15 | 0.65 | 0.52 |
| Habitat:Predator occurrence | −0.3 | 0.98 | −3.05 | <0.001 ** |
| Habitat:Pond size | 0.15 | 0.42 | 0.35 | 0.73 |
| Habitat:Subermerged vegetation | 0.53 | 0.42 | 1.25 | 0.21 |
| Habitat:Surrounding vegetation | −0.74 | 0.22 | −0.34 | 0.73 |
| Habitat turbidity | 0.2 | 0.22 | 0.85 | 0.73 |
| Final generalized linear model (AIC = 349.9) | ||||
| Habitat | 0.47 | 0.3 | 1.58 | 0.12 |
| Desiccation | −0.18 | 0.07 | −2.71 | <0.01 ** |
| Leaf litter | 0.13 | 0.05 | 2.44 | 0.01 * |
| Turbidity | 0.2 | 0.1 | 2.09 | 0.04 * |
| Predators | 0.18 | 0.07 | 2.61 | 0.01 ** |
| Submerged vegetation | 0.09 | 0.08 | 1.19 | 0.23 |
| Habitat:Submerged vegetation | 0.61 | 0.37 | 1.66 | 0.1 |
| Habitat:Predator occurrence | −0.3 | 0.09 | −3.28 | <0.001 ** |
| Variables | Mantel Statistic r | Significance |
|---|---|---|
| Habitat | 0.43 | p < 0.001 * |
| Desiccation | 0.13 | p < 0.001 * |
| Pond size | 0.19 | p < 0.001 * |
| Submerged vegetation | 0.27 | p < 0.001 * |
| Surrounding vegetation | 0.28 | p < 0.001 * |
| Turbidity | −0.01 | 0.61 |
| Predator occurrence | 0.08 | 0.02 * |
| Leaf litter | 0.12 | p < 0.001 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mausberg, N.; Dausmann, K.H.; Glos, J. In Search of Suitable Breeding Sites: Habitat Heterogeneity and Environmental Filters Determine Anuran Diversity of Western Madagascar. Animals 2023, 13, 3744. https://doi.org/10.3390/ani13233744
Mausberg N, Dausmann KH, Glos J. In Search of Suitable Breeding Sites: Habitat Heterogeneity and Environmental Filters Determine Anuran Diversity of Western Madagascar. Animals. 2023; 13(23):3744. https://doi.org/10.3390/ani13233744
Chicago/Turabian StyleMausberg, Nanäa, Kathrin H. Dausmann, and Julian Glos. 2023. "In Search of Suitable Breeding Sites: Habitat Heterogeneity and Environmental Filters Determine Anuran Diversity of Western Madagascar" Animals 13, no. 23: 3744. https://doi.org/10.3390/ani13233744