

First Molecular-Based Confirmation of Dermacentor marginatus and Associated Rickettsia raoultii and Anaplasma marginale in the Hindu Kush Mountain Range

 ,
,  , ,
, ,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
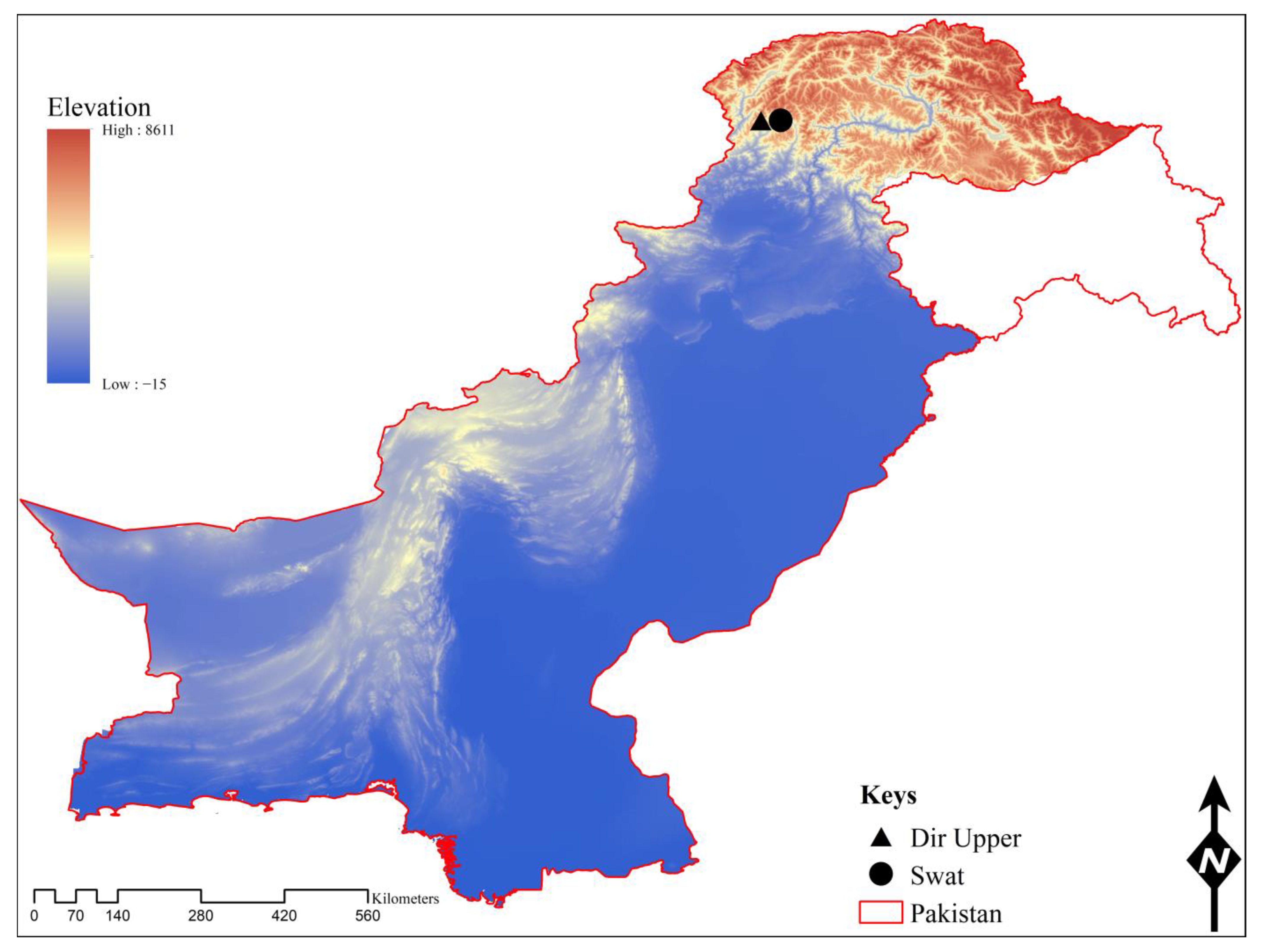
2.2. Study Area
2.3. Ticks Collection and Morphological Identification
2.4. DNA Extraction and PCR
2.5. DNA Sequencing and Phylogenetic Analysis
3. Results
3.1. Tick Record
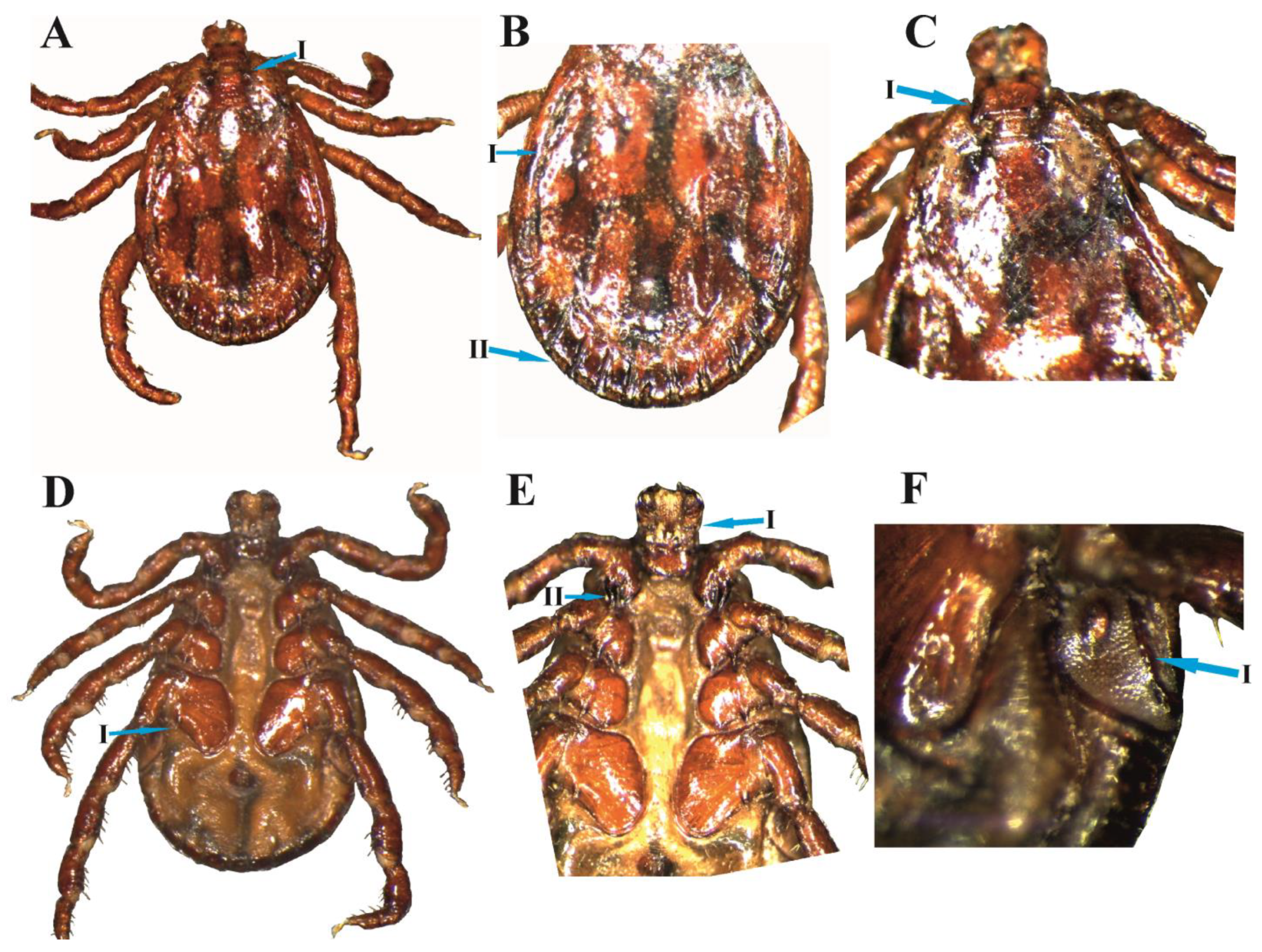
3.2. D. marginatus Male
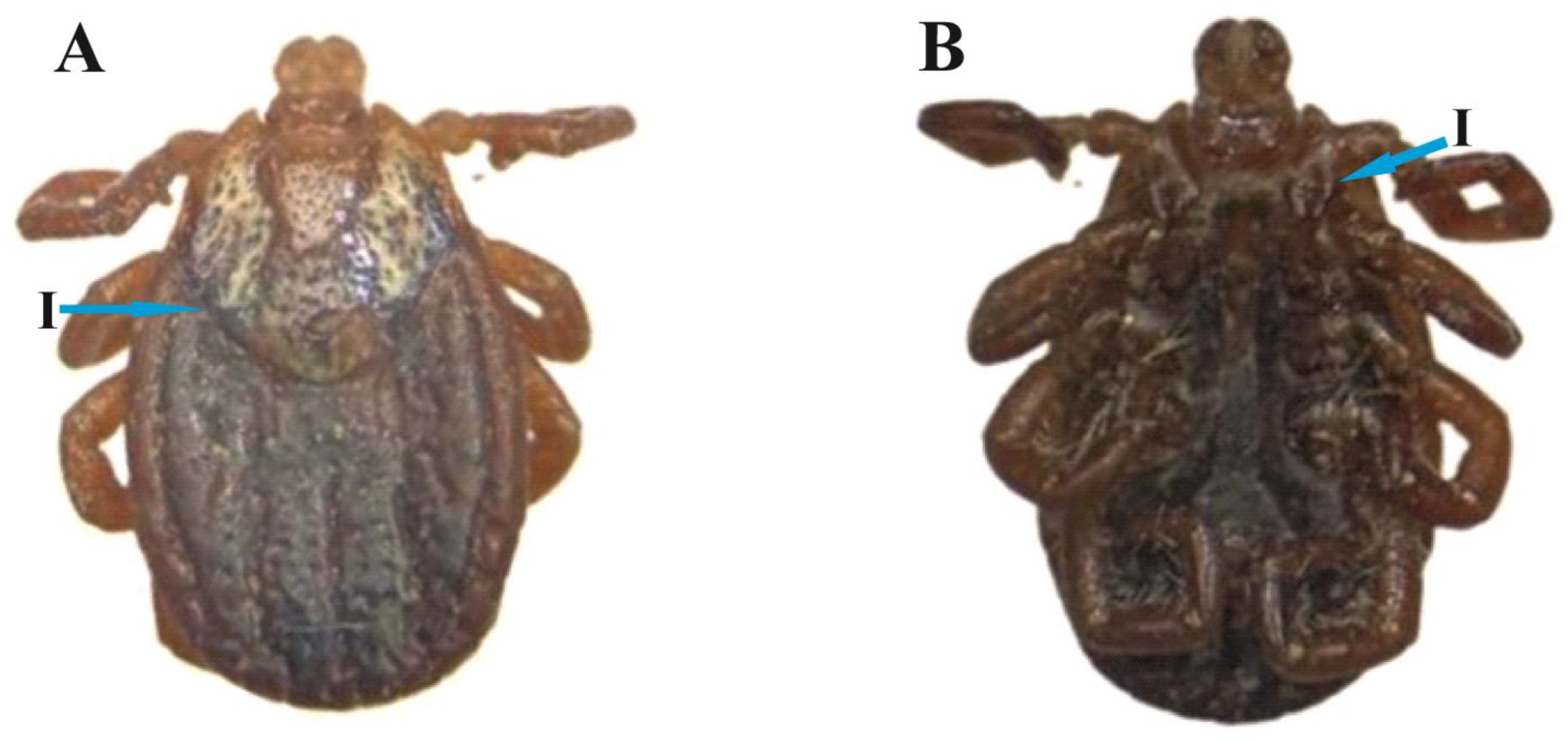
3.3. D. marginatus Female
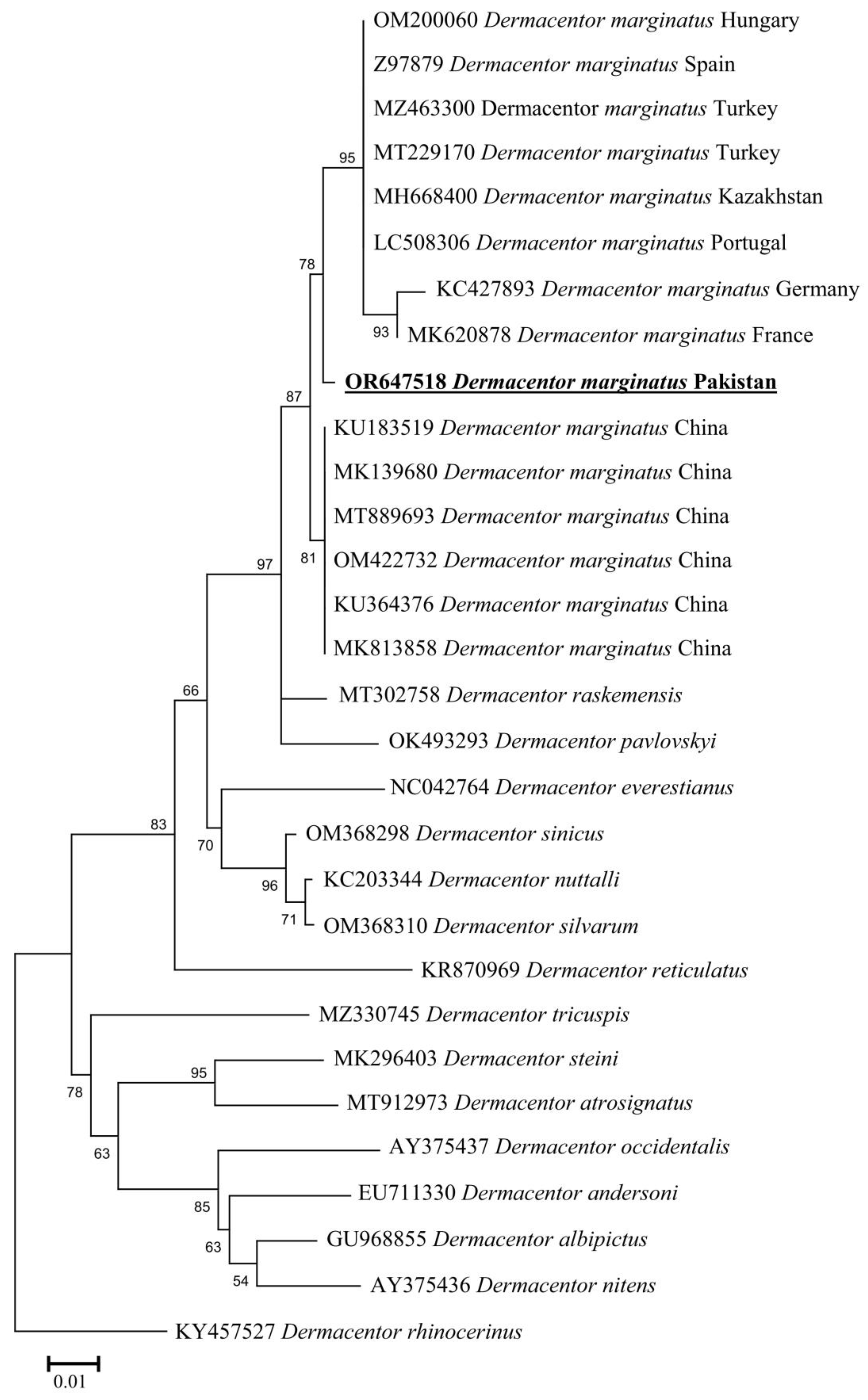
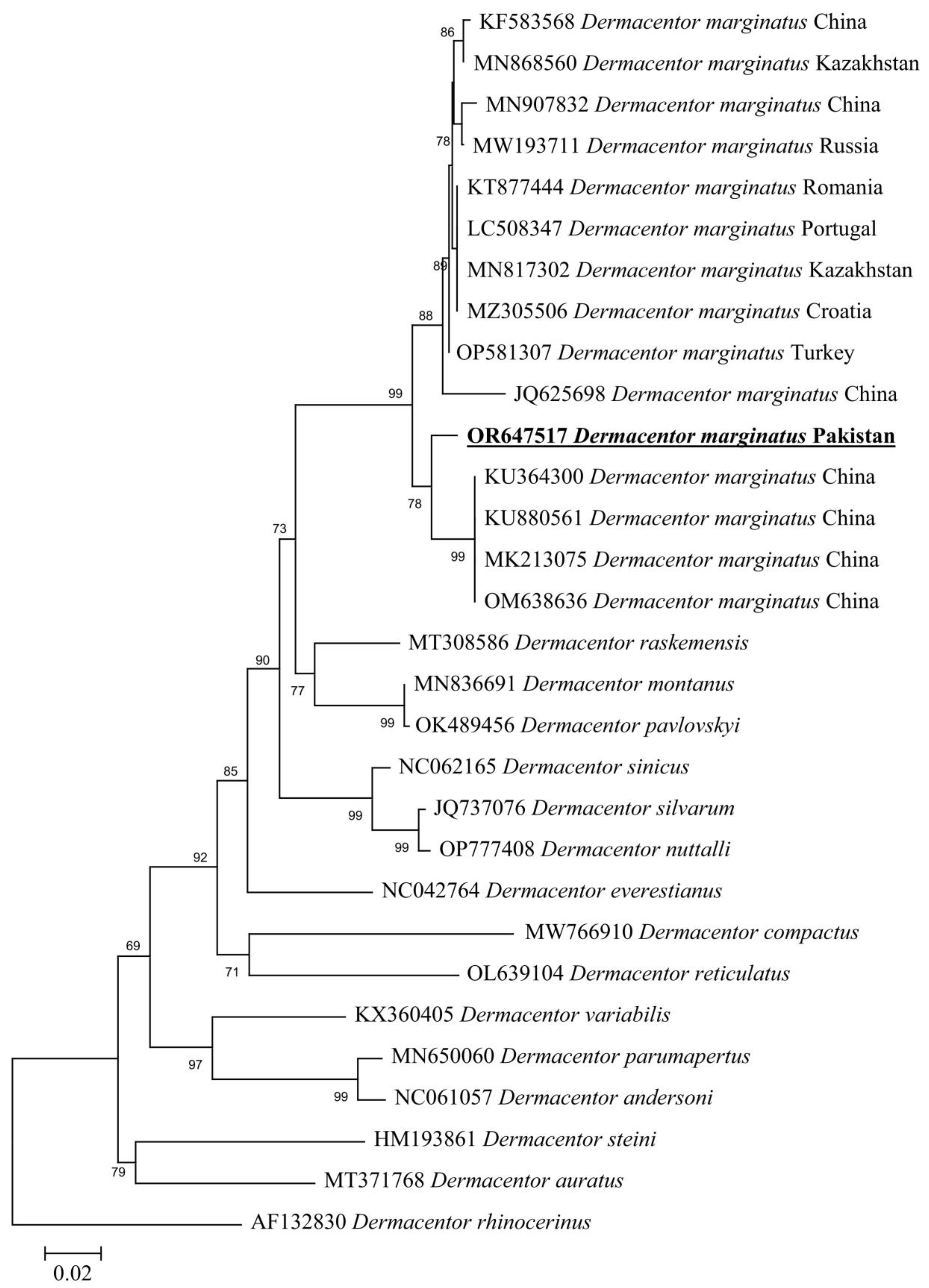
3.4. Molecular and Phylogenetic Analysis of D. marginatus
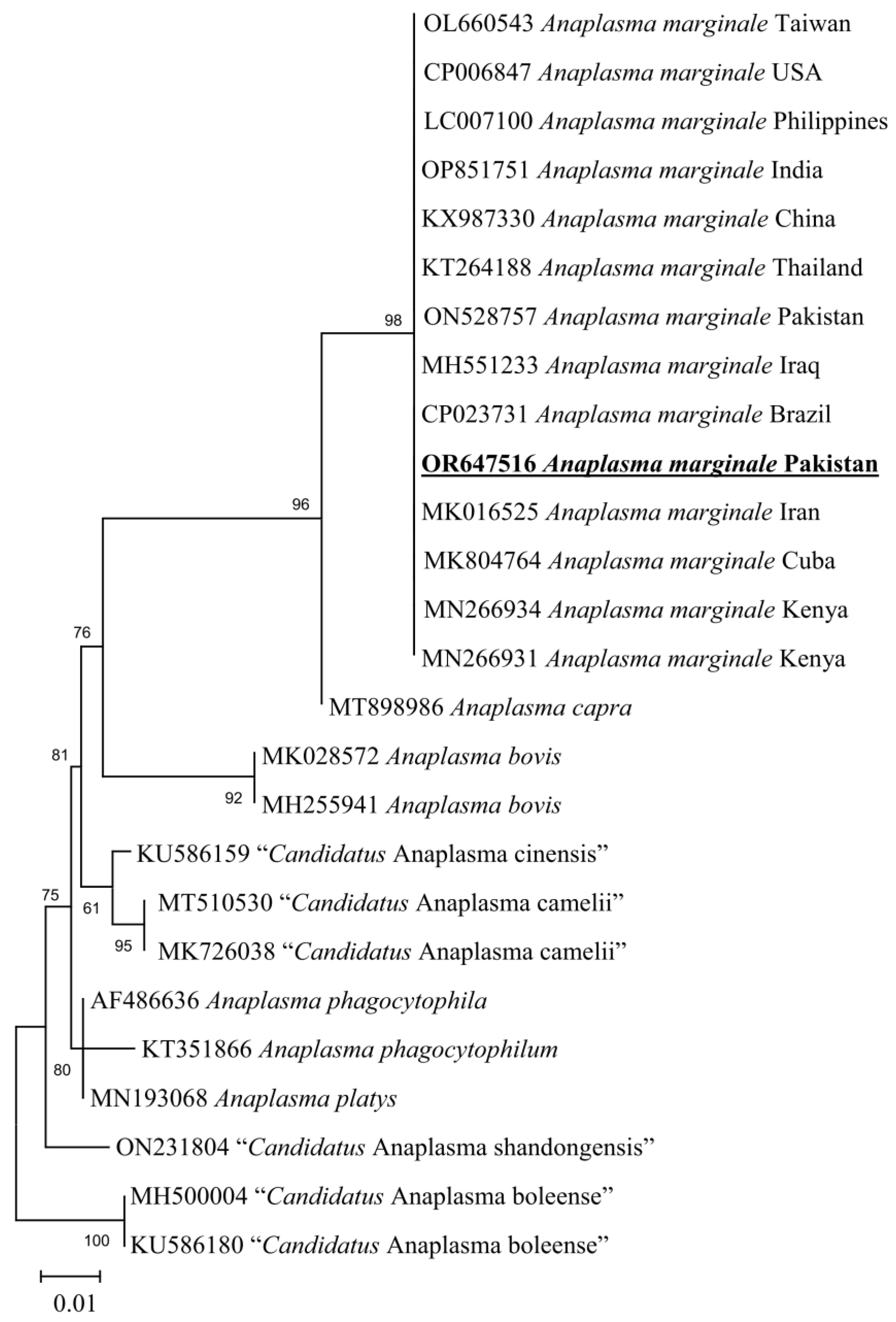
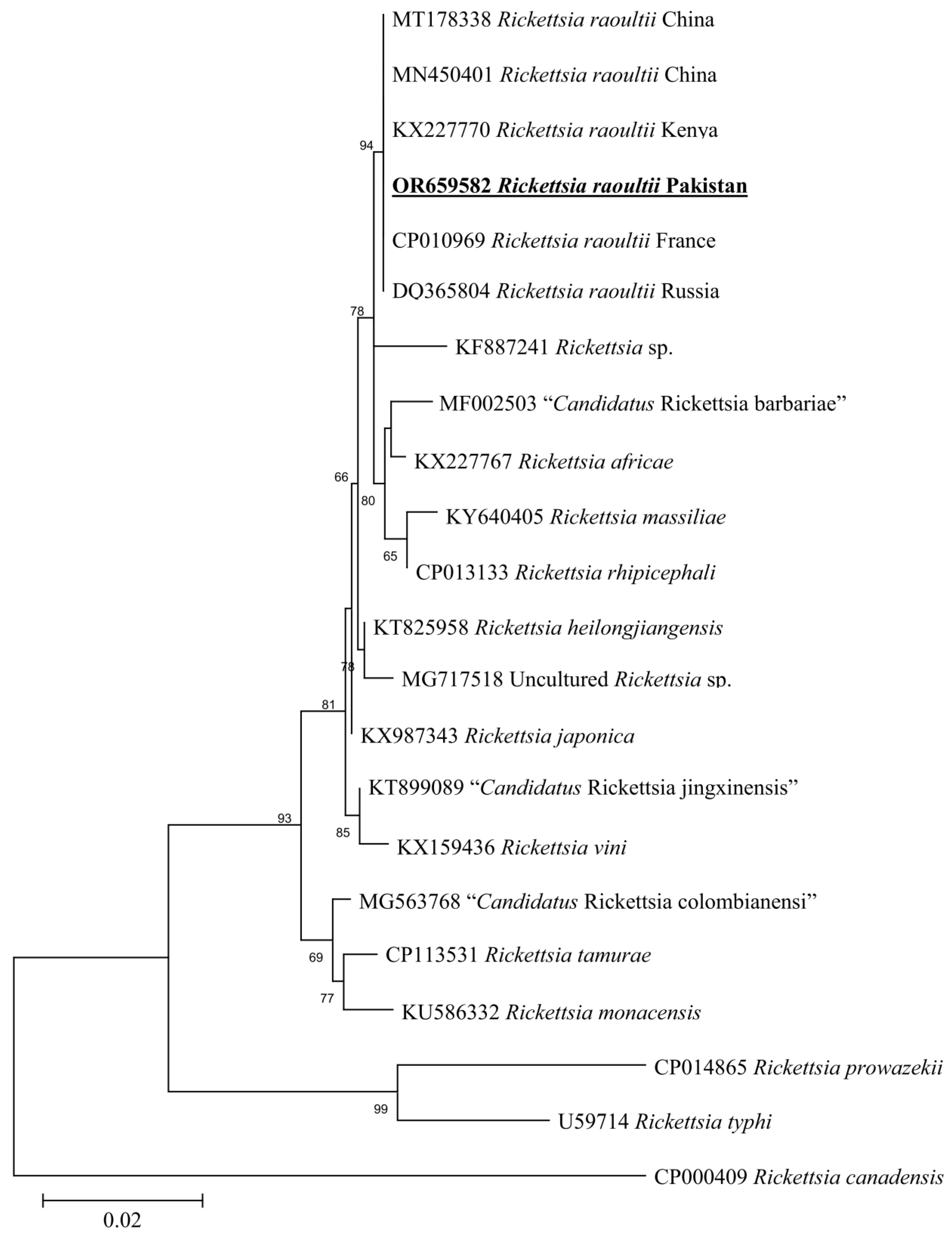
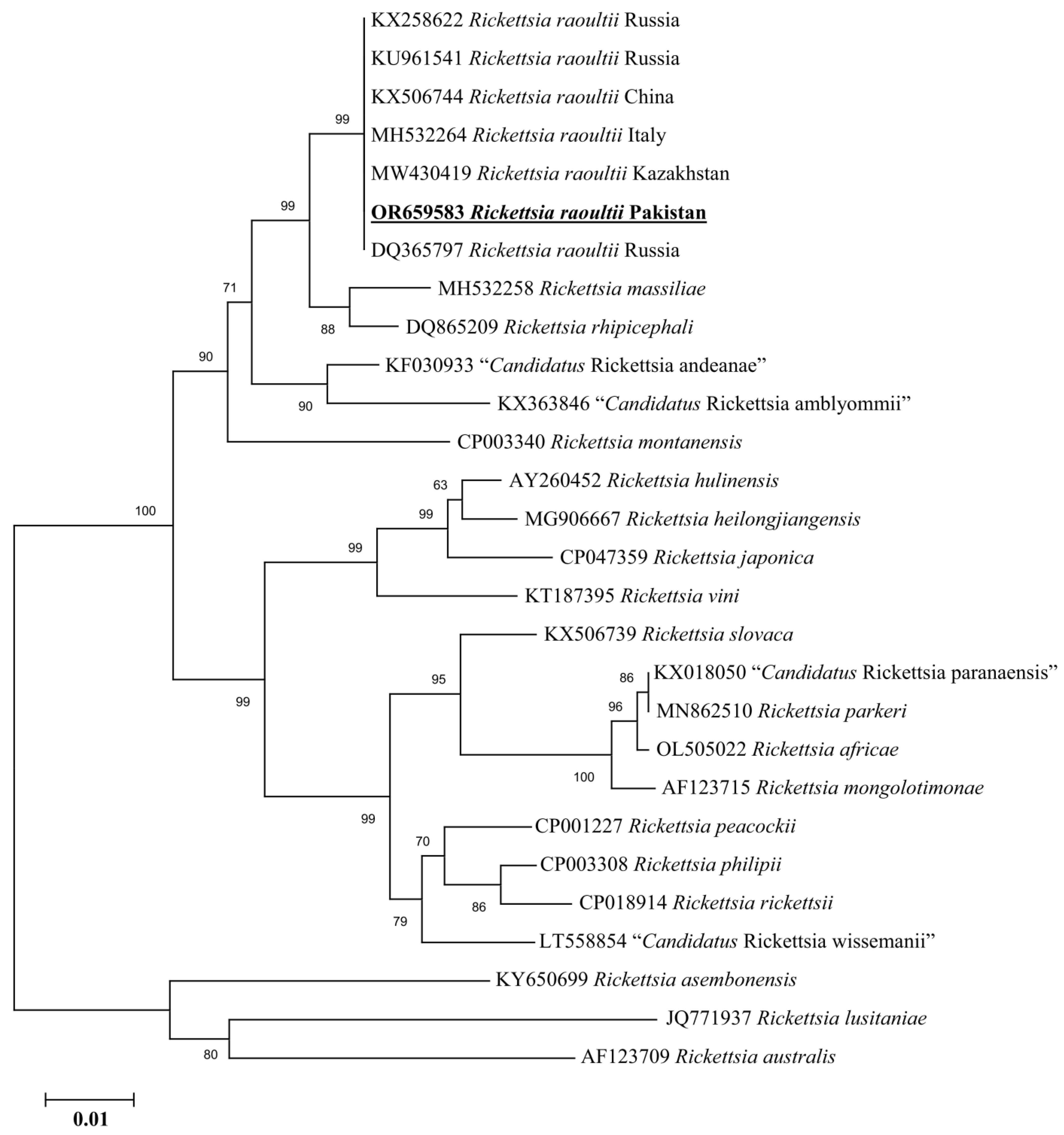
3.5. Molecular Screening and Phylogenetic Analysis of D. marginatus Associated Pathogens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I.G. The Hard Ticks of the World; Springer: Dordrecht, The Netherlands, 2014; Volume 10, pp. 978–994. [Google Scholar]
- Lado, P.; Klompen, H. Evolutionary history of New World ticks of the genus Dermacentor (Ixodida: Ixodidae); the origin of D. variabilis. Biol. J. Linn. Soc. 2019, 127, 863–875. [Google Scholar] [CrossRef]
- Guzmán-Cornejo, C.; Robbins, R.G.; Guglielmone, A.A.; Montiel-Parra, G.; Rivas, G.; Pérez, T.M. The Dermacentor (Acari, Ixodida, Ixodidae) of Mexico: Hosts, geographical distribution and new records. Zookeys 2016, 569, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Estrada-Peña, R. Notes on Dermacentor ticks: Redescription of D. marginatus with the synonymies of D. niveus; D. daghestanicus (Acari: Ixodidae). J. Med. Entomol. 1991, 28, 1–15. [Google Scholar] [CrossRef]
- Raad, M.; Azar, D.; Perotti, M.A. First report of the ticks Haemaphysalis punctata Canestrini et Fanzago, 1878, Haemaphysalis parva (Neumann, 1897) and Dermacentor marginatus (Sulzer, 1776) (Acari, Amblyommidae) from humans in Lebanon. Acta Parasitol. 2020, 65, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Guglielmone, A.A.; Nava, S.; Robbins, R.G. Geographic distribution of the hard ticks (Acari: Ixodida: Ixodidae) of the world by countries and territories. Zootaxa 2023, 5251, 1–274. [Google Scholar] [CrossRef] [PubMed]
- Filippova, N.A.; Plaksina, M.A. Some aspects of intraspecific variability of the closely related species of the Dermacentor marginatus complex (Acari: Ixodidae) as demonstration of microevolutionary process. Parazitologiya 2005, 39, 337–364. [Google Scholar]
- Apanaskevich, D.A.; Apanaskevich, M.A. Description of a new Dermacentor (Acari: Ixodidae) species from Thailand and Vietnam. J. Med. Entomol. 2015, 52, 806–812. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Apanaskevich, M.A.; Nooma, W.; Ahantarig, A.; Trinachartvanit, W. Reinstatement of Dermacentor tricuspis (Schulze, 1933) n. comb.; n. stat. (Acari: Ixodidae) as a valid species, synonymization of D. atrosignatus Neumann, 1906; description of a new species from Indonesia, Malaysia and Thailand. Syst. Parasitol. 2021, 98, 207–230. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Apanaskevich, M.A. Description of two new species of Dermacentor Koch, 1844 (Acari: Ixodidae) from Oriental Asia. Syst. Parasitol. 2016, 93, 159–171. [Google Scholar] [CrossRef]
- Dhanda, V.; Kulkarni, S.M.; Pratt, P. Dermacentor raskemensis (Ixodoidea: Ixodidae), redescription and notes on ecology. J. Parasitol. 1971, 57, 1324–1329. [Google Scholar] [CrossRef]
- Filippova, N.A. Redescription of Dermacentor raskemensis Pomerantzev, 1946 (Ixodidae) a representative of the mountain fauna of the southern regions of the USSR and adjacent territories. Parazitologiia 1983, 17, 283–292. [Google Scholar] [PubMed]
- Apanaskevich, D.A. First description of the nymph and larva of Dermacentor raskemensis (Acari: Ixodidae), parasites of pikas and other small mammals in Central Asia. J. Med. Entomol. 2013, 50, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Masala, G.; Chisu, V.; Satta, G.; Socolovschi, C.; Raoult, D.; Parola, P. Rickettsia slovaca from Dermacentor marginatus ticks in Sardinia, Italy. Ticks Tick Borne Dis. 2012, 3, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Rubel, F.; Brugger, K.; Pfeffer, M.; Chitimia-Dobler, L.; Didyk, Y.M.; Leverenz, S.; Dautel, H.; Kahl, O. Geographical distribution of Dermacentor marginatus and Dermacentor reticulatus in Europe. Ticks Tick Borne Dis. 2016, 7, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, S.; De La Fuente, J.; Nicollet, P.; Liu, X.; Madani, N.; Blanchard, B.; Maingourd, C.; Alongi, A.; Torina, A.; Fernández de Mera, I.G.; et al. Prevalence of tick-borne pathogens in adult Dermacentor spp. ticks from nine collection sites in France. Vector Borne Zoonotic Dis. 2013, 13, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Rubel, F.; Brugger, K.; Belova, O.A.; Kholodilov, I.S.; Didyk, Y.M.; Kurzrock, L.; García-Pérez, A.L.; Kahl, O. Vectors of disease at the northern distribution limit of the genus Dermacentor in Eurasia: D. reticulatus and D. silvarum. Exp. Appl. Acarol. 2020, 82, 95–123. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Qiu, Z.; Zhang, T.; Li, Y.; Wang, W.; Li, M.; Yu, Z.; Liu, J. Microbial diversity in the tick Argas japonicus (Acari: Argasidae) with a focus on Rickettsia pathogens. Med. Vet. Entomol. 2019, 33, 327–335. [Google Scholar] [CrossRef]
- Špitalská, E.; Štefanidesová, K.; Kocianová, E.; Boldiš, V. Rickettsia slovaca; Rickettsia raoultii in Dermacentor marginatus and Dermacentor reticulatus ticks from Slovak Republic. Exp. Appl. Acarol. 2012, 57, 189–197. [Google Scholar] [CrossRef]
- Remesar, S.; Díaz, P.; Portillo, A.; Santibáñez, S.; Prieto, A.; Díaz-Cao, J.M.; López, C.M.; Panadero, R.; Fernández, G.; Díez-Baños, P.; et al. Prevalence and molecular characterization of Rickettsia spp. in questing ticks from north-western Spain. Exp. Appl. Acarol. 2019, 79, 267–278. [Google Scholar] [CrossRef]
- Alkishe, A.; Cobos, M.E.; Osorio-Olvera, L.; Peterson, A.T. Ecological niche and potential geographic distributions of Dermacentor marginatus and Dermacentor reticulatus (Acari: Ixodidae) under current and future climate conditions. Web Ecol. 2022, 22, 33–45. [Google Scholar] [CrossRef]
- Nosek, J.; Sixl, W. Central-European ticks (Ixodoidea). Pomerantzev Mitt. Abt. Zool. Landesmus Joanneum 1972, 1, 480. [Google Scholar]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. (Eds.) Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Sambrook, J.; Fritsch, E.E.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Hoeh, W.R.; Black, M.B.; Vrijenhoek, R.C. Conserved primers for PCR amplification of mitochondrial DNA from different invertebrate phyla. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Inokuma, H.; Raoult, D.; Brouqui, P. Detection of Ehrlichia platys DNA in brown dog ticks (Rhipicephalus sanguineus) in Okinawa Island, Japan. J. Clin. Microbiol. 2000, 38, 4219–4221. [Google Scholar] [CrossRef] [PubMed]
- Labruna, M.B.; Whitworth, T.; Horta, M.C.; Bouyer, D.H.; McBride, J.W.; Pinter, A.; Popov, V.; Gennari, S.M.; Walker, D.H. Rickettsia species infecting Amblyomma cooperi ticks from an area in the state of Sao Paulo, Brazil, where Brazilian spotted fever is endemic. J. Clin. Microbiol. 2004, 42, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rompB (ompB). Int. J. Syst. Evol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensensitivity of progressive multiple sequence alignments through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Apanaskevich, D.A.; Vongphayloth, K.; Jeangkhwoa, P.; Chaloemthanetphong, A.; Ahantarig, A.; Apanaskevich, M.A.; Brey, P.T.; Lakeomany, K.; Trinachartvanit, W. Description of a new species of Dermacentor Koch, 1844 (Acari: Ixodidae) from the mountains of Laos and Thailand. Syst. Parasitol. 2020, 97, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Guglielmone, A.A.; Petney, T.N.; Robbins, R.G. Ixodidae (Acari: Ixodoidea): Descriptions and redescriptions of all known species from 1758 to December 31, 2019. Zootaxa 2020, 4871, 1–322. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, H.R.; Schindler, S.; Adam, M.; Essl, F.; Rabitsch, W. Evidence for changes in the occurrence, frequency or severity of human health impacts resulting from exposure to alien species in Europe: A systematic map. Environ. Evid. 2017, 6, 21. [Google Scholar] [CrossRef]
- Reynolds, S.; Hedberg, M.; Herrin, B.; Chelladurai, J.R.J. Analysis of the complete mitochondrial genomes of Dermacentor albipictus suggests a species complex. Ticks Tick Borne Dis. 2022, 13, 102038. [Google Scholar] [CrossRef] [PubMed]
- Barker, S.C.; Murrell, A. Phylogeny, evolution and historical zoogeography of yicks: A review of recent progress. Exp. Appl. Acarol. 2002, 28, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wu, S.; Zhang, Y.; Chen, Y.; Feng, C.; Yuan, X.; Jia, G.; Deng, J.; Wang, C.; Wang, Q.; et al. Assessment of four DNA fragments (COI, 16S rDNA, ITS2, 12S rDNA) for species identification of the Ixodida (Acari: Ixodida). Parasit. Vectors 2014, 7, 93. [Google Scholar] [CrossRef]
- Shehla, S.; Ullah, F.; Alouffi, A.; Almutairi, M.M.; Khan, Z.; Tanaka, T.; Labruna, M.B.; Tsai, K.H.; Ali, A. Association of SFG Rickettsia massiliae and Candidatus Rickettsia shennongii with different hard ticks infesting livestock hosts. Pathogens 2023, 12, 1080. [Google Scholar] [CrossRef]
- Ali, A.; Obaid, M.K.; Almutairi, M.M.; Alouffi, A.; Numan, M.; Ullah, S.; Rehman, G.; Islam, Z.U.; Khan, S.B.; Tanaka, T. Molecular detection of Coxiella spp. in ticks (Ixodidae and Argasidae) infesting domestic and wild animals: With notes on the epidemiology of tick-borne Coxiella burnetii in Asia. Front. Microbiol. 2023, 14, 1229950. [Google Scholar] [CrossRef]
- Khan, M.; Islam, N.; Khan, A.; Islam, Z.U.; Muñoz-Leal, S.; Labruna, M.B.; Ali, A. New records of Amblyomma gervaisi from Pakistan, with detection of a reptile-associated Borrelia sp. Ticks Tick Borne Dis. 2022, 13, 102047. [Google Scholar] [CrossRef]
- Khan, Z.; Shehla, S.; Alouffi, A.; Kashif Obaid, M.; Zeb Khan, A.; Almutairi, M.M.; Numan, M.; Aiman, O.; Alam, S.; Ullah, S.; et al. Molecular survey and genetic characterization of Anaplasma marginale in ticks collected from livestock hosts in Pakistan. Animals 2022, 12, 1708. [Google Scholar] [CrossRef]
- Ullah, K.; Numan, M.; Alouffi, A.; Almutairi, M.M.; Zahid, H.; Khan, M.; Islam, Z.U.; Kamil, A.; Safi, S.Z.; Ahmed, H.; et al. Molecular characterization and assessment of risk factors associated with Theileria annulata infection. Microorganisms 2022, 10, 1614. [Google Scholar] [CrossRef]
- Zhang, Y.K.; Yu, Z.J.; Wang, D.; Bronislava, V.; Branislav, P.; Liu, J.Z. The bacterial microbiome of field-collected Dermacentor marginatus and Dermacentor reticulatus from Slovakia. Parasit. Vectors 2019, 12, 325. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Target Genes | Sequences (5′-3′) | Size bp | PCR Conditions | References |
|---|---|---|---|---|---|
| Ticks | 16S rRNA | 16S+1-CCGGTCTGAACTCAGATCAAGT 16S−1-CTCAATGATTTTTTAAATTGCTG | 460 | 95 °C 3 min, 40 × (95 °C 30 s, 55 °C 60 s, 72 °C 1 min), 72 °C 7 min | [26] |
| cox1 | HC02198-TAAACTTCAGGGTGACCAAAAAATCA LCO1490-GGTCAACAAATCATAAAGATATTGG | 712 | 95 °C 30 s, 40 × (95 °C 30 s, 48 °C 30 s, 72 °C 1 min), 72 °C 5 min | [27] | |
| Pathogens | 16S rRNA | EHR16SD-GGTACCYACAGAAGAAGTCC EHR16SR-TAGCACTCATCGTTTACAGC | 344 | 95 °C 5 min, 35 × (95 °C 30 s, 55 °C 30 s, 72 °C 90 s), 72 °C 5 min | [28] |
| gltA | CS-78 GCAAGTATCGGTGAGGATGTAAT CS-323 GCTTCCTTAAAATTCAATAAATCAGGAT | 401 | 95 °C 3 min, 40 × (95 °C 15 s, 48 °C 30 s, 72 °C 30 s) 72 °C 7 min | [29] | |
| ompB | 120-M59-CCGCAGGGTTGGTAACTGC 120-807-CCTTTTAGATTACCGCCTAA | 862 | 95 °C 3 min, 40 × (95 °C 30 s, 50 °C 30 s, 68 °C 1 min 30 s), 68 °C 7 min | [30] |
| District | Host | Sex | Age Group | Temperature and Humidity | Place of Collection and Elevation | Tick Species | |
|---|---|---|---|---|---|---|---|
| Targeted Species: D. marginatus | Accompanied Species: H. montgomeryi | ||||||
| Dir Upper | Goat | Female | Adult (above 1 year) | 12 °C, 78% | Kund Banda, 3347 m | 2M | 2M, 1F * |
| Dir Upper | Goat | Male | Young (above 6 months–1 year) | 13 °C, 78% | Gaedar Banda, 3344 m | 1M, 2F | 3M |
| Dir Upper | Goat | Male | Kid (below 6 months) | 13 °C, 78% | Bend Banda, 3340 m | 2M | 4F |
| Dir Upper | Goat | Female | Young (above 6 months–1 year) | 15 °C, 78% | Jan Shahi, 3340 m | 2M, 1F * | 1M, 3F |
| Dir Upper | Goat | Female | Adult (above 1 year) | 13 °C, 78% | Cherry Banda, 3349 m | 2M | 2M |
| Dir Upper | Goat | Female | Kid (below 6 months) | 14 °C, 78% | Dand Banda, 3352 m | 1M | 4M, 1F * |
| Dir Upper | Goat | Male | Young (above 6 months–1 year) | 14 °C, 78% | Otalshai, 3348 m | 3M | 2M, 4F |
| Swat | Goat | Male | Kid (below 6 months) | 15 °C, 78% | Dirgal, 3340 m | 1M, 3F | 4M |
| Swat | Goat | Female | Adult (above 1 year) | 15 °C, 78% | Gorgal, 3345 m | 2M | 5F |
| Swat | Goat | Female | Young (above 6 months–1 year) | 15 °C, 78% | Landai dara, 3336 m | 1M, 1F | 3M, 1F * |
| Swat | Goat | Male | Adult (above 1 year) | 15 °C, 78% | Dasht e Liala, 3351 m | 2M | 2M, 3F * |
| Total | 11 goats | 5 males, 6 females | 4 adults, 4 young, 3 kids | 19M, 7F | 23M, 22F | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, I.; Ullah, S.; Alouffi, A.; Almutairi, M.M.; Numan, M.; Tanaka, T.; Chang, S.-C.; Chen, C.-C.; Ali, A. First Molecular-Based Confirmation of Dermacentor marginatus and Associated Rickettsia raoultii and Anaplasma marginale in the Hindu Kush Mountain Range. Animals 2023, 13, 3686. https://doi.org/10.3390/ani13233686
Ahmad I, Ullah S, Alouffi A, Almutairi MM, Numan M, Tanaka T, Chang S-C, Chen C-C, Ali A. First Molecular-Based Confirmation of Dermacentor marginatus and Associated Rickettsia raoultii and Anaplasma marginale in the Hindu Kush Mountain Range. Animals. 2023; 13(23):3686. https://doi.org/10.3390/ani13233686
Chicago/Turabian StyleAhmad, Iftikhar, Shafi Ullah, Abdulaziz Alouffi, Mashal M. Almutairi, Muhammad Numan, Tetsuya Tanaka, Shun-Chung Chang, Chien-Chin Chen, and Abid Ali. 2023. "First Molecular-Based Confirmation of Dermacentor marginatus and Associated Rickettsia raoultii and Anaplasma marginale in the Hindu Kush Mountain Range" Animals 13, no. 23: 3686. https://doi.org/10.3390/ani13233686