
Comparison of Subcutaneous versus Intramuscular Dexmedetomidine–Midazolam–Ketamine–Morphine (DMKM) Mixture as Chemical Restraint for Endoscopic Sex Determination in Aldabra Giant Tortoises (Aldabrachelys gigantea)
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Design
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bour, R. Recherches Sur Des Animaux Doublement Disparus: Les Tortues Gé Antes Subfossiles De Madagascar; Ecole Pratique Des Hautes Études: Montpellier, France, 1994; p. 253. [Google Scholar]
- Bunbury, N.; von Brandis, R.; Currie, J.C.; van de Crommenacker, J.; Accouche, W.; Birch, D.; Chong-Seng, L.; Doak, N.; Haupt, P.; Haverson, P.; et al. Late stage dynamics of a successful feral goat eradication from the UNESCO world heritage site of aldabra atoll, seychelles. Biol. Invasions 2018, 20, 1735–1747. [Google Scholar] [CrossRef]
- Gerlach, J.; Rocamora, G.; Gane, J.; Jolliffe, K.; Vanherck, L. Giant tortoise distribution and abundance in the seychelles islands: Past, present, and future. Chelonian Conserv. Biol. 2013, 12, 70–83. [Google Scholar] [CrossRef]
- Gerlach, J. Giant Tortoises of the Indian Ocean: The Genus Dipsochelys Inhabiting the Seychelles Islands and the Extinct Giants of Madagascar and the Mascarenes; Edition Chimaira: Frankfurt, Germany, 2004; p. 207. [Google Scholar]
- Griffiths, O.; Andre, A.; Meunier, A. Tortoise breeding and ‘re-wilding’on Rodrigues Island. Chelonian Res. Monogr. 2013, 6, 178–182. [Google Scholar] [CrossRef]
- Gerlach, J. Development of distinct morphotypes in captive seychelles–aldabra giant tortoises. Chelonian Conserv. Biol. 2011, 10, 102–112. [Google Scholar] [CrossRef]
- Kuchling, G.; Griffiths, O. Endoscopic imaging of gonads, sex ratios, and occurrence of intersexes in juvenile captive-bred aldabra giant tortoises. Chelonian Conserv. Biol. 2012, 11, 91–96. [Google Scholar] [CrossRef]
- Swingland, I.R. Reproductive effort and life history strategy of the aldabran giant tortoise. Nature 1977, 269, 402–404. [Google Scholar] [CrossRef]
- Hernandez-Divers, S.J.; Stahl, S.J.; Farrell, R. An endoscopic method for identifying sex of hatchling Chinese box turtles and comparison of general versus local anesthesia for coelioscopy. J. Am. Vet. Med. Assoc. 2009, 234, 800–804. [Google Scholar] [CrossRef]
- Sladky, K.K.; Mans, C. Clinical anesthesia in reptiles. J. Exot. Pet Med. 2012, 21, 17–31. [Google Scholar] [CrossRef]
- Innis, C.J.; Hernandez-Divers, S.; Martinez-Jimenez, D. Coelioscopic-assisted prefemoral oophorectomy in chelonians. J. Am. Vet. Med. Assoc. 2007, 230, 1049–1052. [Google Scholar] [CrossRef]
- Read, M.R. Evaluation of the use of anesthesia and analgesia in reptiles. J. Am. Vet. Med. Assoc. 2004, 224, 547–552. [Google Scholar] [CrossRef]
- Sleeman, J.M.; Gaynor, J. Sedative and cardiopulmonary effects of medetomidine and reversal with atipamezole in desert tortoises (Gopherus agassizii). J. Zoo Wildl. Med. 2000, 31, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Vigani, A. Chelonia (tortoises, turtles, and terrapins). Zoo Anim. Wildl. Immobil. Anesth. 2014, 25, 365–387. [Google Scholar] [CrossRef]
- Dundee, J.W.; Halliday, N.J.; Harper, K.W.; Brogden, R.N. Midazolam. A review of its pharmacological properties and therapeutic use. Drugs 1984, 28, 519–543. [Google Scholar] [CrossRef] [PubMed]
- Aitken-Palmer, C.; Heard, D.; Jacobson, E.; Hall, N.; Thieman, K.; Ellison, G. Clinical Management of Cloacal Prolapse in an Adult Galapagos Tortoise (Geochelone nigra); Veterinary Medical Center, University of Florida: Gainesville, FL, USA, 2010; p. 47. [Google Scholar]
- Mans, C.; Sladky, K.K. Endoscopically guided removal of cloacal calculi in three African spurred tortoises (Geochelone sulcata). J. Am. Vet. Med. Assoc. 2012, 240, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Bienzle, D.; Boyd, C.J. Sedative effects of ketamine and midazolam in snapping turtles (Chelydra serpentina). J. Zoo Wildl. Med. 1992, 23, 201–204. [Google Scholar]
- Chittick, E.J.; Stamper, M.A.; Beasley, J.F.; Lewbart, G.A.; Horne, W.A. Medetomidine, ketamine, and sevoflurane for anesthesia of injured loggerhead sea turtles: 13 cases (1996–2000). J. Am. Vet. Med. Assoc. 2002, 221, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Sladky, K.K.; Kinney, M.E.; Johnson, S.M. Analgesic efficacy of butorphanol and morphine in bearded dragons and corn snakes. JAMA 2008, 233, 267. [Google Scholar] [CrossRef]
- Sladky, K.K.; Miletic, V.; Paul-Murphy, J.; Kinney, M.E.; Dallwig, R.K.; Johnson, S.M. Analgesic efficacy and respiratory effects of butorphanol and morphine in turtles. J. Am. Vet. Med. Assoc. 2007, 230, 1356–1362. [Google Scholar] [CrossRef]
- Mosley, C. Pain and nociception in reptiles. Vet. Clin. Exot. Anim. Pract. 2011, 14, 45–60. [Google Scholar] [CrossRef]
- Scheelings, T.F.; Gatto, C.; Reina, R.D. Anaesthesia of hatchling green sea turtles (Chelonia mydas) with intramuscular ketamine-medetomidine-tramadol. Aust. Vet. J. 2020, 98, 511–516. [Google Scholar] [CrossRef]
- Mans, C.; Foster, J.D. Endoscopy-guided ectopic egg removal from the urinary bladder in a leopard tortoise (Stigmochelys pardalis). Can. Vet. J. 2014, 55, 569–572. [Google Scholar] [PubMed]
- Schumacher, J.; Yelen, T. Anesthesia and Analgesia; Reptile Medicine and Surgery: London, UK, 2006. [Google Scholar]
- Falcón, W.; Baxter, R.P.; Furrer, S.; Bauert, M.; Hatt, J.M.; Schaepman-Strub, G.; Ozgul, A.; Bunbury, N.; Clauss, M.; Hansen, D.M. Patterns of activity and body temperature of Aldabra giant tortoises in relation to environmental temperature. Ecol. Evol. 2018, 8, 2108–2121. [Google Scholar] [CrossRef] [PubMed]
- White, N.M.; Balasubramaniam, T.; Nayak, R.; Barnett, A.G. An observational analysis of the trope “A p-value of < 0.05 was considered statistically significant” and other cut-and-paste statistical methods. PLoS ONE 2022, 17, e0264360. [Google Scholar] [PubMed]
- Mc Shane, B.B.; Gal, D.; Gelman, A.; Robert, C.; Tackett, J.L. Abandon statistical significance. Am. Stat. 2019, 73, 235–245. [Google Scholar] [CrossRef]
- Hurlbert, S.H.; Levine, R.A.; Utts, J. Coup de grâce for a tough old bull: “Statistically significant” expires. Am. Stat. 2019, 73, 352–357. [Google Scholar] [CrossRef]
- Divers, S.J.; Stahl, S.J. Mader’s Reptile and Amphibian Medicine and Surgery; Elsevier Health Sciences: London, UK, 2018; p. 1537. [Google Scholar]
- Lahner, L.; Mans, C.; Sladky, K. Comparison of Anesthetic Induction and Recovery Times After Intramuscular, Subcutaneous or Intranasal Dexmedetomidine-Ketamine Administration in Red-Eared Slider Turtles (Trachemys Scripta Elegans). In Proceedings of the Conference American Association of Zoo Veterinarians, Kansas City, MO, USA, 27 October 2011. [Google Scholar]
- Doss, G.A.; Fink, D.M.; Sladky, K.K.; Mans, C. Comparison of subcutaneous dexmedetomidine-midazolam versus alfaxalone-midazolam sedation in leopard geckos (Eublepharis macularius). Vet. Anaesth. Analg. 2017, 44, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, S.J.; Cox, S.; Yaw, T.J.; Sladky, K. Pharmacokinetics of subcutaneously administered hydromorphone in bearded dragons (Pogona vitticeps) and red-eared slider turtles (Trachemys scripta elegans). Vet. Anaesth. Analg. 2019, 46, 352–359. [Google Scholar] [CrossRef]
- Abdala, V.; Manzano, A.S.; Herrel, A. The distal forelimb musculature in aquatic and terrestrial turtles: Phylogeny or environmental constraints? J. Anat. 2008, 213, 159–172. [Google Scholar] [CrossRef]
- Eshar, D.; Rooney, T.A.; Gardhouse, S.; Beaufrère, H. Evaluation of the effects of a dexmedetomidine-midazolam-ketamine combination administered intramuscularly to captive red-footed tortoises (Chelonoidis carbonaria). Am. J. Vet. Res. 2021, 82, 858–864. [Google Scholar] [CrossRef]
- Berry, S.H. Injectable anesthetics. In Veterinary Anesthesia and Analgesia: The Fifth Edition of Lumb and Jones; Wiley: Hoboken, NJ, USA, 2015; pp. 277–296. [Google Scholar]
- Murrell, J.C.; Hellebrekers, L.J. Medetomidine and dexme-detomidine: A review of cardiovascular effects and antinociceptive properties in the dog. Vet. Anaesth. Analg. 2005, 32, 117–127. [Google Scholar] [CrossRef]
- Kallio-Kujala, I.J.; Raekallio, M.R.; Honkavaara, J.; Bennett, R.C.; Turunen, H.; Scheinin, M.; Vainio, O. Peripheral alpha-2-ad- renoreceptor antagonism affects the absorption of intramuscularly coadministered drugs. Vet. Anaesth. Analg. 2018, 45, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Porters, N.; De Rooster, H.; Bosmans, T.; Baert, K.; Cherlet, M.; Croubels, S.; Polis, I. Pharmacokinetics of oral transmucosal and in-tramuscular dexmedetomidine combined with buprenorphine in cats. J. Vet. Pharmacol. Ther. 2014, 38, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, J. Chelonians (turtles, tortoises, and terrapins). In Zoo Animal and Wildlife Immobilization and Anaesthesia; Blackwell Publishing: Hoboken, NJ, USA, 2007; pp. 259–266. [Google Scholar]
- Girling, S.J.; Raiti, P. BSAVA Manual of Reptiles, 3rd ed.; British Small Animal Veterinary Association: Quedgeley, UK, 2019. [Google Scholar]
- Rodney, W.; Molly, S. Section 6 Anesthesia. Chapter 48 Sedation. In Mader’s Reptile and Amphibian Medicine and Surgery; Divers, S.J., Stahl, S.J., Eds.; Elsevier Health Sciences: London, UK, 2018; p. 1537. [Google Scholar]
- Divers, S.J. Endoscopic sex identification in chelonians and birds (psittacines, passerines, and raptors). Vet. Clin. N. Am. Exot. Anim. Pract. 2015, 18, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Meireles, Y.S.; Shinike, F.S.; Matte, D.R.; Morgado, T.O.; Kempe, G.V.; Corrêa, S.H.R.; Souza, R.L.d.; Néspoli, P.B. Ultrasound characterization of the coelomic cavity organs of the red-footed tortoise (Chelonoidis carbonaria). Ciênc. Rural 2016, 46, 1811–1817. [Google Scholar] [CrossRef]
- Scarabelli, S.; Di Girolamo, N. Chelonian sedation and anesthesia. Vet. Clin. Exot. Anim. Pract. 2022, 25, 49–72. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.L.; Bertelsen, M.F. Assessment of the effects of intramuscular administration of alfaxalone with and without medetomidine in Horsfield’s tortoises (Agrionemys horsfieldii). Vet. Anaesth. Analg. 2013, 40, e68–e75. [Google Scholar] [CrossRef] [PubMed]
- Karklus, A.A.; Sladky, K.K.; Johnson, S.M. Respiratory and antinociceptive effects of dexmedetomidine and doxapram in ball pythons (Python regius). Am. J. Vet. Res. 2021, 82, 11–21. [Google Scholar] [CrossRef]







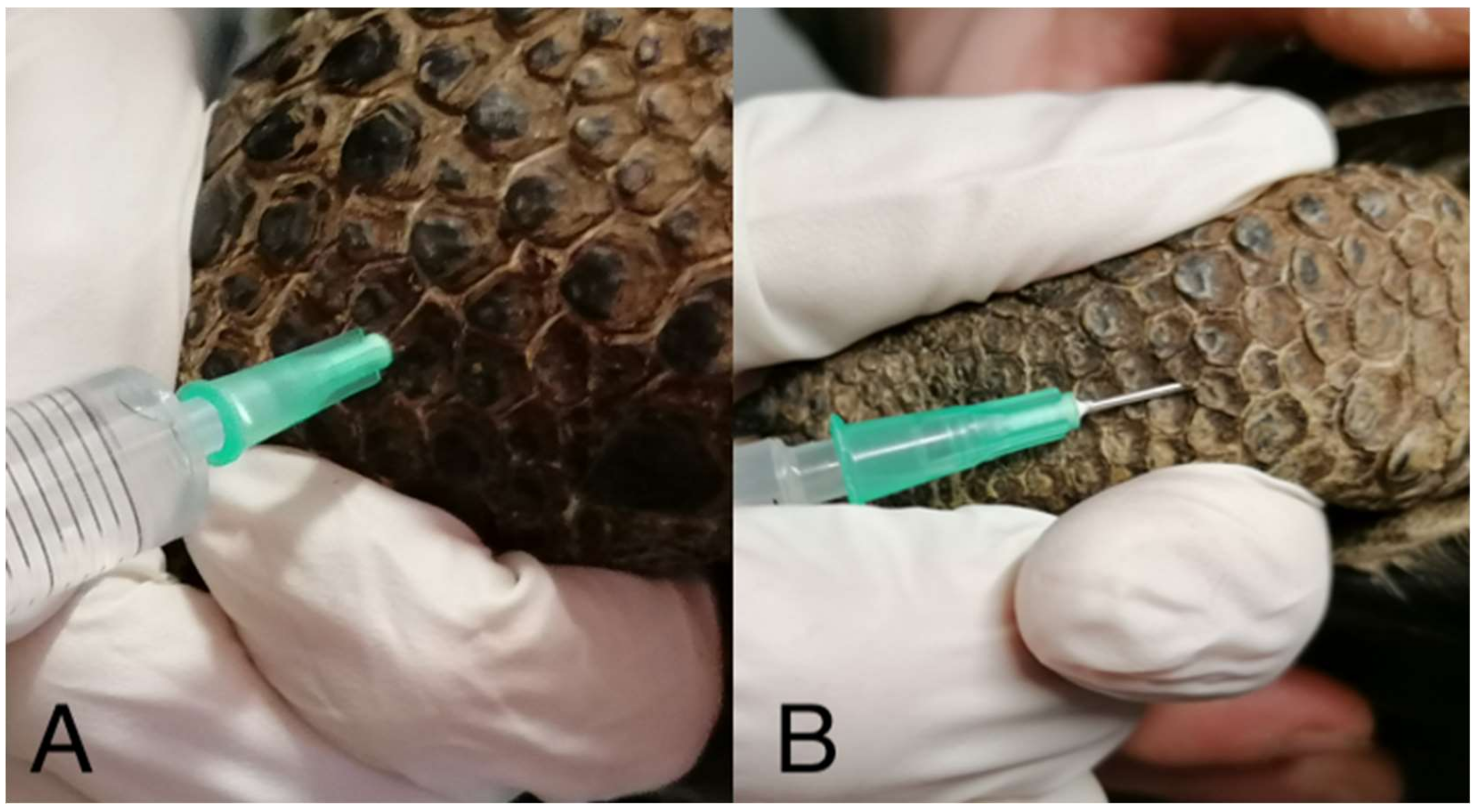{kind=link}
{kind=link}
{kind=link}
{kind=link}
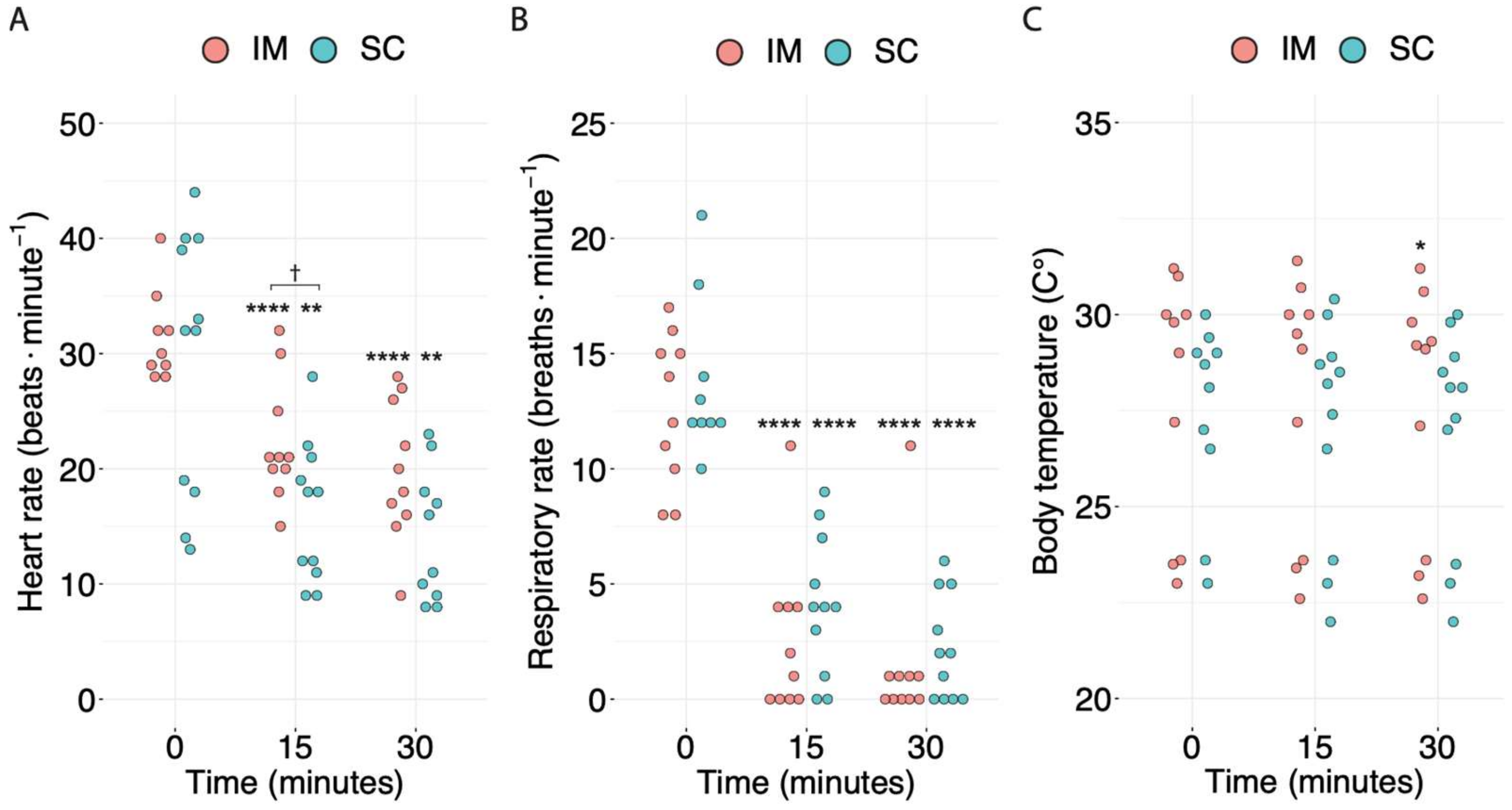{kind=link}
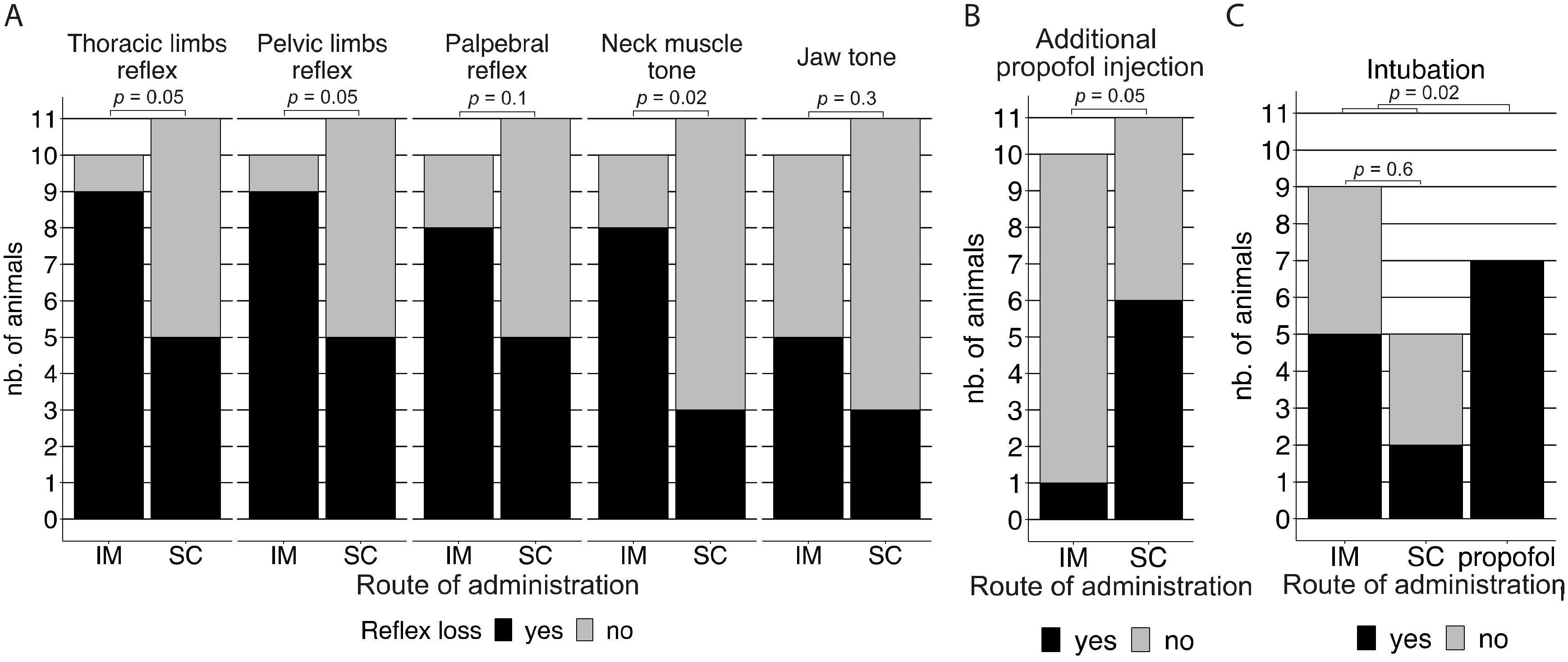{kind=link}
| Parameter | IM DMKM | SC DMKM | p-Value |
|---|---|---|---|
| Time to loss of the thoracic limb reflex for positive cases (minutes) | 16.9 (8.4–25.3) (n = 9/9) | 12.5 (6.3–20.6) (n = 5/5) | 0.6 |
| Time to loss of the pelvic limb reflex for positive cases (minutes) | 16.9 (8.4–25.3) (n = 9/9) | 17.5 (9.4–23.8) (n = 5/5) | 1 |
| Time to loss of the palpebral reflex for positive cases (minutes) | 18.8 (10–26.25) (n = 8/9) | 18.8 (9.4–28.1) (n = 5/5) | 1 |
| Time to loss of neck muscle tone for positive cases (minutes) | 30 (15.0–37.5) (n = 8/9) | 7.5 (3.8–21.2) (n = 3/5) | 0.06 |
| Time to loss of jaw tone for positive cases (minutes) | 22.5 (9.4–36.6) (n = 5/9) | 18.8 (11.3–24.4) (n = 3/5) | 0.8 |
| Parameter | IM DMKM | SC DMKM | p-Value |
|---|---|---|---|
| Time to resumption of the thoracic limb reflex (minutes) | 8.8 (4.4–13.1) (n = 7) | 22.5 (11.3–33.8) (n = 3) | 0.2 |
| Time to resumption of the pelvic limb reflex (minutes) | 7.5 (3.8–11.3) (n = 8) | 11.25 (5.6–18.8) (n = 3) | 0.2 |
| Time to resumption of the palpebral reflex (minutes) | 10.5 (5.3–16.9) (n =7) | 9.4 (4.7–14.1) (n = 5) | 0.7 |
| Time to resumption of neck muscle tone (minutes) | 10.5 (5.3–16.9) (n = 7) | 11.3 (5.6–18.8) (n = 3) | 0.9 |
| Time to resumption of jaw tone (minutes) | 9.4 (4.7–14.1) (n = 5) | 7.5 (3.8–11.6) (n = 2) | 0.8 |
| Time to complete resumption of all reflexes (minutes) | 13.1 (6.6–24.4) (n = 8) | 12.5 (6.3–26.3) (n = 5) | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masi, M.; Vetere, A.; Casalini, J.; Corsi, F.; Di Ianni, F.; Nardini, G. Comparison of Subcutaneous versus Intramuscular Dexmedetomidine–Midazolam–Ketamine–Morphine (DMKM) Mixture as Chemical Restraint for Endoscopic Sex Determination in Aldabra Giant Tortoises (Aldabrachelys gigantea). Animals 2023, 13, 3626. https://doi.org/10.3390/ani13233626
Masi M, Vetere A, Casalini J, Corsi F, Di Ianni F, Nardini G. Comparison of Subcutaneous versus Intramuscular Dexmedetomidine–Midazolam–Ketamine–Morphine (DMKM) Mixture as Chemical Restraint for Endoscopic Sex Determination in Aldabra Giant Tortoises (Aldabrachelys gigantea). Animals. 2023; 13(23):3626. https://doi.org/10.3390/ani13233626
Chicago/Turabian StyleMasi, Marco, Alessandro Vetere, Jacopo Casalini, Flavia Corsi, Francesco Di Ianni, and Giordano Nardini. 2023. "Comparison of Subcutaneous versus Intramuscular Dexmedetomidine–Midazolam–Ketamine–Morphine (DMKM) Mixture as Chemical Restraint for Endoscopic Sex Determination in Aldabra Giant Tortoises (Aldabrachelys gigantea)" Animals 13, no. 23: 3626. https://doi.org/10.3390/ani13233626