
Phytobiotic-Prebiotic Feed Additive Containing a Combination of Carob Pulp, Chicory, and Fenugreek Improve Growth Performance, Carcass Traits, and Fecal Microbiota of Fattening Pigs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management and Experimental Design
2.2. Meat Sampling and Carcass Classification
2.3. Fecal Sample Analysis: Enumeration, DNA Extraction and Sequencing
2.4. Bioinformatics
2.5. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Carcass Quality
3.3. Fecal Microbiota Composition
3.3.1. Enumeration
3.3.2. Sequencing
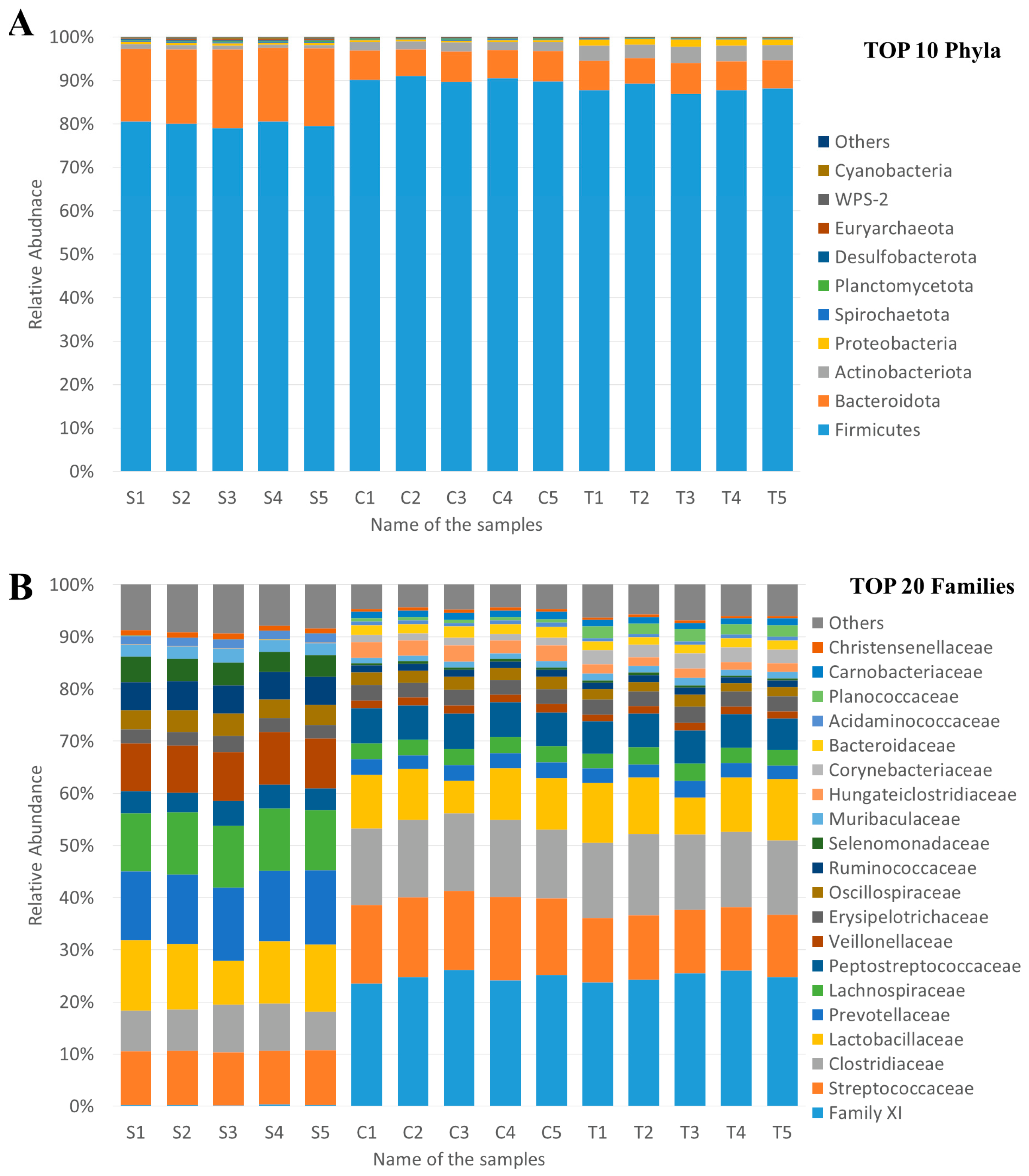
3.3.3. Taxonomic Profiles and Bacterial Diversity
3.3.4. Treatment-Related Differences
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dunker, A.; Rey, A.I.; López-Bote, C.J.; Daza, A. Effect of the feeding level during the fattening phase on the productive parameters, carcass characteristics and quality of fat in heavy pigs. J. Anim. Feed. Sci. 2007, 16, 621–635. [Google Scholar] [CrossRef]
- Fellström, C.; Holmgren, N. Mice as reservoirs for swine dysentery in a fattening herd. Sv VetTidn 2005, 13, 19–21. [Google Scholar]
- Tsinas, A.C.; Kyriakis, S.C.; Lekkas, S.; Sarris, K.; Bourtzi-Hatzopoulou, E.; Saoulidis, K. Control of proliferative enteropathy in growing/fattening pigs using growth promoters. Zentralbl Vet. B 1998, 45, 115–127. [Google Scholar] [CrossRef]
- Paul, S.S.; Rama Rao, S.V.; Hegde, N.; Williams, N.J.; Chatterjee, R.N.; Raju, M.; Reddy, G.N.; Kumar, V.; Phani Kumar, P.S.; Mallick, S.; et al. Effects of Dietary Antimicrobial Growth Promoters on Performance Parameters and Abundance and Diversity of Broiler Chicken Gut Microbiome and Selection of Antibiotic Resistance Genes. Front. Microbiol. 2022, 13, 905050. [Google Scholar] [CrossRef]
- Costa, M.C.; Bessegatto, J.A.; Alfieri, A.A.; Weese, J.S.; Filho, J.A.; Oba, A. Different antibiotic growth promoters induce specific changes in the cecal microbiota membership of broiler chicken. PLoS ONE 2017, 12, e0171642. [Google Scholar] [CrossRef]
- Li, L.; Sun, X.; Zhao, D.; Dai, H. Pharmacological Applications and Action Mechanisms of Phytochemicals as Alternatives to Antibiotics in Pig Production. Front. Immunol. 2021, 12, 798553. [Google Scholar] [CrossRef]
- Mendez-Palacios, N.; Mendez-Mendoza, M.; Vazquez-Flores, F.; Castro-Colombres, J.G.; Ramirez-Bribiesca, J.E. Productive and economic parameters of pigs supplemented from weaning to finishing with prebiotic and probiotic feed additives. Anim. Sci. J. 2018, 89, 994–1001. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Rodríguez-Solana, R.; Romano, A.; Moreno-Rojas, J.M. Carob Pulp: A Nutritional and Functional By-Product Worldwide Spread in the Formulation of Different Food Products and Beverages. A Review. Processes 2021, 9, 1146. [Google Scholar] [CrossRef]
- Biagia, G.; Cipollini, I.; Paulicks, B.R.; Roth, F.X. Effect of tannins on growth performance and intestinal ecosystem in weaned piglets. Arch. Anim. Nutr. 2010, 64, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Inserra, L.; Luciano, G.; Bella, M.; Scerra, M.; Cilione, C.; Basile, P.; Lanza, M.; Priolo, A. Effect of including carob pulp in the diet of fattening pigs on the fatty acid composition and oxidative stability of pork. Meat Sci. 2015, 100, 256–261. [Google Scholar] [CrossRef]
- Kotrotsios, N.; Christaki, E.; Bonos, E.; Florou-Paneri, P. Dietary carob pods on growth performance and meat quality of fattening pigs. Asian-Australas J. Anim. Sci. 2012, 25, 880–885. [Google Scholar] [CrossRef]
- Liu, H.; Ivarsson, E.; Dicksved, J.; Lundh, T.; Lindberg, J.E. Inclusion of chicory (Cichorium intybus L.) in pigs’ diets affects the intestinal microenvironment and the gut microbiota. Appl. Environ. Microbiol. 2012, 78, 4102–4109. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.R.; Awati, A.; Smidt, H.; Williams, B.A.; Akkermans, A.D.; de Vos, W.M. Specific response of a novel and abundant Lactobacillus amylovorus-like phylotype to dietary prebiotics in the guts of weaning piglets. Appl. Environ. Microbiol. 2004, 70, 3821–3830. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chen, D.; Yu, B.; Huang, Z.; Luo, Y.; Zheng, P.; Mao, X.; Yu, J.; Luo, J.; He, J. Effect of Dietary Inulin Supplementation on Growth Performance, Carcass Traits, and Meat Quality in Growing-Finishing Pigs. Animals 2019, 9, 840. [Google Scholar] [CrossRef]
- Windisch, W.; Schedle, K.; Plitzner, C.; Kroismayr, A. Use of phytogenic products as feed additives for swine and poultry1. J. Anim. Sci. 2008, 86, E140–E148. [Google Scholar] [CrossRef]
- Zhai, H.; Liu, H.; Wang, S.; Wu, J.; Kluenter, A.M. Potential of essential oils for poultry and pigs. Anim. Nutr. 2018, 4, 179–186. [Google Scholar] [CrossRef]
- Huang, C.M.; Lee, T.T. Immunomodulatory effects of phytogenics in chickens and pigs—A review. Asian-Australas J. Anim. Sci. 2018, 31, 617–627. [Google Scholar] [CrossRef]
- Michiels, J.; Missotten, J.; Dierick, N.; Fremaut, D.; Maene, P.; De Smet, S. In vitro degradation and in vivo passage kinetics of carvacrol, thymol, eugenol and trans-cinnamaldehyde along the gastrointestinal tract of piglets. J. Sci. Food Agric. 2008, 88, 2371–2381. [Google Scholar] [CrossRef]
- Ren, H.; Vahjen, W.; Dadi, T.; Saliu, E.M.; Boroojeni, F.G.; Zentek, J. Synergistic Effects of Probiotics and Phytobiotics on the Intestinal Microbiota in Young Broiler Chicken. Microorganisms 2019, 7, 684. [Google Scholar] [CrossRef] [PubMed]
- Zentek, J.; Gartner, S.; Tedin, L.; Manner, K.; Mader, A.; Vahjen, W. Fenugreek seed affects intestinal microbiota and immunological variables in piglets after weaning. Br. J. Nutr. 2013, 109, 859–866. [Google Scholar] [CrossRef]
- Lin, B.; Gong, J.; Wang, Q.; Cui, S.; Yu, H.; Huang, B. In-vitro assessment of the effects of dietary fibers on microbial fermentation and communities from large intestinal digesta of pigs. Food Hydrocoll. 2011, 25, 180–188. [Google Scholar] [CrossRef]
- Hossain, M.M.; Begum, M.; Kim, I.H. Effects of fenugreek (Trigonella foenum-graecum L.) seed extract supplementation in different energy density diets on growth performance, nutrient digestibility, blood characteristics, fecal microbiota, and fecal gas emission in growing pigs. Can. J. Anim. Sci. 2018, 98, 289–298. [Google Scholar] [CrossRef]
- Juhasz, A.; Molnar-Nagy, V.; Bata, Z.; Tso, K.H.; Mayer, Z.; Posta, K. Alternative to ZnO to establish balanced intestinal microbiota for weaning piglets. PLoS ONE 2022, 17, e0265573. [Google Scholar] [CrossRef]
- Sarri, L.; Costa-Roura, S.; Balcells, J.; Seradj, A.R.; de la Fuente, G. The Impact of Genetics on Gut Microbiota of Growing and Fattening Pigs under Moderate N Restriction. Animals 2021, 11, 2846. [Google Scholar] [CrossRef]
- ISO-14502-1:2005; Determination of Substances Characteristic of Green and Black Tea. Part 1: Content of Total Polyphenols in Tea—Colorimetric Method Using Folin-Ciocalteu Reagent. International Organization for Standardization: Geneva, Switzerland, 2005.
- Union, E. Council Directive 2008/120/EC of 18 December 2008 laying down minimum standards for the protection of pigs (Codified version). Off. J. Eur. Union 2009, 47, 5–13. [Google Scholar]
- İnaç, S.; Gücükoğlu, A. The S-Europ System in Classification the Quality of Cattle Carcasses and Monitoring Methods in Carcass Grading. Turk. J. Agric. Food Sci. Technol. 2020, 8, 27–34. [Google Scholar]
- Kvapilík, J.; Přibyl, J.; Růžička, Z.; Řehák, D. Results of pig carcass classification according to SEUROP in the Czech Republic. Czech J. Anim. Sci. 2009, 54, 217–228. [Google Scholar] [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2017, 34, 1287–1294. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Mahe, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and fast clustering method for amplicon-based studies. PeerJ 2014, 2, e593. [Google Scholar] [CrossRef]
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, W20–W25. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Kemény, S.; Deák, A. Kísérletek Tervezése és Értékelése; Műszaki Könyvkiadó: Budapest, Hungary, 2000. [Google Scholar]
- Fox, J.; Leanage, A. R and the Journal of Statistical Software. J. Stat. Softw. 2016, 73, 1–13. [Google Scholar] [CrossRef]
- Chen, S.; Luo, S.; Yan, C. Gut Microbiota Implications for Health and Welfare in Farm Animals: A Review. Animals 2021, 12, 93. [Google Scholar] [CrossRef]
- Kesavelu, D.; Jog, P. Current understanding of antibiotic-associated dysbiosis and approaches for its management. Ther. Adv. Infect. Dis. 2023, 10, 20499361231154443. [Google Scholar] [CrossRef]
- Francino, M.P. Antibiotics and the Human Gut Microbiome: Dysbioses and Accumulation of Resistances. Front. Microbiol. 2015, 6, 1543. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, D.; Nigam, P.S. Antibiotic-Therapy-Induced Gut Dysbiosis Affecting Gut Microbiota-Brain Axis and Cognition: Restoration by Intake of Probiotics and Synbiotics. Int. J. Mol. Sci. 2023, 24, 3074. [Google Scholar] [CrossRef]
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef]
- Millet, S.; Maertens, L. The European ban on antibiotic growth promoters in animal feed: From challenges to opportunities. Vet. J. 2011, 187, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Alzawqari, M.; Saleh, A.; Al-Zaaim, M.; Hamoda, M.; Abu Khalifa, M.; Fouad, M.; Abu Khalifa, M. Dietary Effect of Antibiotic Growth Promoter and Essential Oil on Growth Performance, Carcass Traits and Some Physiological Indicators in Broilers. New Val. J. Agric. Sci. 2021, 1, 1–9. [Google Scholar] [CrossRef]
- Selim, S.; Seleiman, M.F.; Hassan, M.M.; Saleh, A.A.; Mousa, M.A. Impact of Dietary Supplementation with Moringa oleifera Leaves on Performance, Meat Characteristics, Oxidative Stability, and Fatty Acid Profile in Growing Rabbits. Animals 2021, 11, 248. [Google Scholar] [CrossRef]
- Grela, E.R.; Swiatkiewicz, M.; Florek, M.; Bakowski, M.; Skiba, G. Effect of Inulin Source and a Probiotic Supplement in Pig Diets on Carcass Traits, Meat Quality and Fatty Acid Composition in Finishing Pigs. Animals 2021, 11, 2438. [Google Scholar] [CrossRef]
- Wang, W.; Chen, D.; Yu, B.; Huang, Z.; Mao, X.; Zheng, P.; Luo, Y.; Yu, J.; Luo, J.; Yan, H.; et al. Effects of dietary inulin supplementation on growth performance, intestinal barrier integrity and microbial populations in weaned pigs. Br. J. Nutr. 2020, 124, 296–305. [Google Scholar] [CrossRef]
- Kjos, N.P.; Øverland, M.; Fauske, A.K.; Sørum, H. Feeding chicory inulin to entire male pigs during the last period before slaughter reduces skatole in digesta and backfat. Livest. Sci. 2010, 134, 143–145. [Google Scholar] [CrossRef]
- Molbak, L.; Thomsen, L.E.; Jensen, T.K.; Bach Knudsen, K.E.; Boye, M. Increased amount of Bifidobacterium thermacidophilum and Megasphaera elsdenii in the colonic microbiota of pigs fed a swine dysentery preventive diet containing chicory roots and sweet lupine. J. Appl. Microbiol. 2007, 103, 1853–1867. [Google Scholar] [CrossRef] [PubMed]
- Begum, M.; Hossain, M.M.; Kim, I.H. Effects of fenugreek seed extract supplementation on growth performance, nutrient digestibility, diarrhoea scores, blood profiles, faecal microflora and faecal noxious gas emission in weanling piglets. J. Anim. Physiol. Anim. Nutr. 2016, 100. [Google Scholar] [CrossRef]
- Graham, A.; Knopf, B.; Sandberg, F.B.; Soltwedel, K.T.; Bible, M.R.; Greiner, L. 349 Evaluation of a Fenugreek Extract Fed to Sows from 90 Days of Gestation through Lactation on Sow and Litter Performance. J. Anim. Sci. 2018, 96, 188. [Google Scholar] [CrossRef]
- Hossain, M.M.; Begum, M.; Nyachoti, C.M.; Hancock, J.D.; Kim, I.H. Dietary fenugreek seed extract improves performance and reduces fecal E. coli counts and fecal gas emission in lactating sows and suckling piglets. Can. J. Anim. Sci. 2015, 95, 561–568. [Google Scholar] [CrossRef]
- He, M.; Fang, S.; Huang, X.; Zhao, Y.; Ke, S.; Yang, H.; Li, Z.; Gao, J.; Chen, C.; Huang, L. Evaluating the Contribution of Gut Microbiota to the Variation of Porcine Fatness with the Cecum and Fecal Samples. Front. Microbiol. 2016, 7, 2108. [Google Scholar] [CrossRef]
- Gardiner, G.E.; Metzler-Zebeli, B.U.; Lawlor, P.G. Impact of Intestinal Microbiota on Growth and Feed Efficiency in Pigs: A Review. Microorganisms 2020, 8, 1886. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.K.; Chae, J.P.; Pajarillo, E.A.B.; Kim, S.H.; Kwak, M.J.; Eun, J.S.; Chee, S.W.; Whang, K.Y.; Kim, S.H.; Kang, D.K. Association between the body weight of growing pigs and the functional capacity of their gut microbiota. Anim. Sci. J. 2020, 91, e13418. [Google Scholar] [CrossRef]
- Navarro, A.; Rodea, G.E.; Castelán-Sánchez, H.G.; Saucedo-Pastrana, H.A.; Licona-Moreno, D.; Eslava-Campos, C.; Tirado-Gómez, L.L.; Vilchis-Reyes, A.; García de la Torre, G.; Cruz-Licea, V. Importance of Microbiome of Fecal Samples Obtained from Adolescents with Different Weight Conditions on Resistance Gene Transfer. Microorganisms 2022, 10, 1995. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ren, W.; Smidt, H.; Wright, A.G.; Yu, B.; Schyns, G.; McCormack, U.M.; Cowieson, A.J.; Yu, J.; He, J.; et al. Dynamic Distribution of Gut Microbiota in Pigs at Different Growth Stages: Composition and Contribution. Microbiol. Spectr. 2022, 10, e0068821. [Google Scholar] [CrossRef]
- Tan, Z.; Wang, Y.; Yang, T.; Ao, H.; Chen, S.; Xing, K.; Zhang, F.; Zhao, X.; Liu, J.; Wang, C. Differences in gut microbiota composition in finishing Landrace pigs with low and high feed conversion ratios. Antonie Van Leeuwenhoek 2018, 111, 1673–1685. [Google Scholar] [CrossRef]
- Jensen, A.N.; Mejer, H.; Mølbak, L.; Langkjær, M.; Jensen, T.K.; Angen, Ø.; Martinussen, T.; Klitgaard, K.; Baggesen, D.L.; Thamsborg, S.M.; et al. The effect of a diet with fructan-rich chicory roots on intestinal helminths and microbiota with special focus on Bifidobacteria and Campylobacter in piglets around weaning. Animal 2011, 5, 851–860. [Google Scholar] [CrossRef]
- Jensen, A.N.; Hansen, L.L.; Baggesen, D.L.; Mølbak, L. Effects of feeding finisher pigs with chicory or lupine feed for one week or two weeks before slaughter with respect to levels of Bifidobacteria and Campylobacter. Animal 2013, 7, 66–74. [Google Scholar] [CrossRef]
- Wei, S.; Wang, J.; Wang, C.; Wang, Y.; Jin, M. Inulin mitigates high fructose-induced gut dysbiosis and metabolic dysfunction in mice. J. Funct. Foods 2022, 97, 105236. [Google Scholar] [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Wang, J.; Sailer, M.; Theis, S.; Verbeke, K.; Raes, J. Prebiotic inulin-type fructans induce specific changes in the human gut microbiota. Gut 2017, 66, 1968–1974. [Google Scholar] [CrossRef]
- Pouille, C.L.; Ouaza, S.; Roels, E.; Behra, J.; Tourret, M.; Molinié, R.; Fontaine, J.-X.; Mathiron, D.; Gagneul, D.; Taminiau, B.; et al. Chicory: Understanding the Effects and Effectors of This Functional Food. Nutrients 2022, 14, 957. [Google Scholar] [CrossRef]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenterol. 2013, 6, 39–51. [Google Scholar] [CrossRef]
- Khan, I.; Bai, Y.; Zha, L.; Ullah, N.; Ullah, H.; Shah, S.R.H.; Sun, H.; Zhang, C. Mechanism of the Gut Microbiota Colonization Resistance and Enteric Pathogen Infection. Front. Cell. Infect. Microbiol. 2021, 11, 716299. [Google Scholar] [CrossRef]
- Szabo, C.; Kachungwa Lugata, J.; Ortega, A. Gut Health and Influencing Factors in Pigs. Animals 2023, 13, 1350. [Google Scholar] [CrossRef]
- Wang, X.; Tsai, T.; Deng, F.; Wei, X.; Chai, J.; Knapp, J.; Apple, J.; Maxwell, C.V.; Lee, J.A.; Li, Y.; et al. Longitudinal investigation of the swine gut microbiome from birth to market reveals stage and growth performance associated bacteria. Microbiome 2019, 7, 109. [Google Scholar] [CrossRef]
- Yang, H.; Huang, X.; Fang, S.; He, M.; Zhao, Y.; Wu, Z.; Yang, M.; Zhang, Z.; Chen, C.; Huang, L. Unraveling the Fecal Microbiota and Metagenomic Functional Capacity Associated with Feed Efficiency in Pigs. Front. Microbiol. 2017, 8, 1555. [Google Scholar] [CrossRef]
- Nishiyama, K.; Yokoi, T.; Sugiyama, M.; Osawa, R.; Mukai, T.; Okada, N. Roles of the Cell Surface Architecture of Bacteroides and Bifidobacterium in the Gut Colonization. Front. Microbiol. 2021, 12, 754819. [Google Scholar] [CrossRef]
- Tardiolo, G.; Romeo, O.; Zumbo, A.; Di Marsico, M.; Sutera, A.M.; Cigliano, R.A.; Paytuvi, A.; D’Alessandro, E. Characterization of the Nero Siciliano Pig Fecal Microbiota after a Liquid Whey-Supplemented Diet. Animals 2023, 13, 642. [Google Scholar] [CrossRef]
- Wang, C.; Wei, S.; Chen, N.; Xiang, Y.; Wang, Y.; Jin, M. Characteristics of gut microbiota in pigs with different breeds, growth periods and genders. Microb. Biotechnol. 2022, 15, 793–804. [Google Scholar] [CrossRef]
- Oliveira, A.; Oliveira, L.C.; Aburjaile, F.; Benevides, L.; Tiwari, S.; Jamal, S.B.; Silva, A.; Figueiredo, H.C.P.; Ghosh, P.; Portela, R.W.; et al. Insight of Genus Corynebacterium: Ascertaining the Role of Pathogenic and Non-pathogenic Species. Front. Microbiol. 2017, 8, 1937. [Google Scholar] [CrossRef]
- Kim, H.; Cho, J.H.; Song, M.; Cho, J.H.; Kim, S.; Kim, E.S.; Keum, G.B.; Kim, H.B.; Lee, J.H. Evaluating the Prevalence of Foodborne Pathogens in Livestock Using Metagenomics Approach. J. Microbiol. Biotechnol. 2021, 31, 1701–1708. [Google Scholar] [CrossRef]
- Su, H.; Jiang, J.; Lu, Q.; Zhao, Z.; Xie, T.; Zhao, H.; Wang, M. Engineering Corynebacterium crenatum to produce higher alcohols for biofuel using hydrolysates of duckweed (Landoltia punctata) as feedstock. Microb. Cell Factories 2015, 14, 16. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient 1 | |
|---|---|
| Chicory (Cichorium intybus L.) roots 1, % | 50.0 |
| Carob (Ceratonia siliqua) seed, % | 30.0 |
| Fenugreek (Trigonella foenum-graecum) seed, % | 20.0 |
| Total, % | 100.0 |
| Calculated nutrient composition | |
| Dry matter, % | 88.0 |
| Crude protein, % | 10.0 |
| Crude fiber, % | 20.0 |
| Main active compound composition | |
| Inulin 2, % | 2.5 |
| Total polyphenolic compounds, GAE 3/g | 40.0 |
| Ingredient | % | |||
|---|---|---|---|---|
| Grower I (Day 0–42) | Grower II (Day 42–70) | |||
| Barley | 10.5 | 8.5 | ||
| Triticale | 25.0 | 32.0 | ||
| Corn | 36.5 | 35.0 | ||
| Malt germ, 26% | 4.0 | 6.0 | ||
| Soybean meal, 46% | 9.0 | 4.5 | ||
| Extracted rapeseed meal | 0.0 | 5.0 | ||
| Pressed linseed | 3.0 | 0.0 | ||
| Full fat soy | 9.0 | 6.0 | ||
| 1 Premix | 3.0 | 3.0 | ||
| Total | 100.0 | 100.0 | ||
| Calculated nutrient composition | 2 C | 3 T | C | T |
| Dry matter, % | 89.9 | 89.4 | 89.2 | 89.6 |
| Crude protein, % | 15.0 | 15.1 | 14.2 | 14.4 |
| Crude fiber, % | 5.27 | 4.82 | 5.41 | 5.36 |
| Crude fat, % | 3.45 | 3.61 | 2.77 | 2.93 |
| Item | C Group | T Group | p Value |
|---|---|---|---|
| Initial body weight, kg | 32.8 ± 0.5 | 32.7 ± 0.5 | 0.6220 |
| Final body weight, kg | 104.9 ± 1.6 b | 106.7 ± 1.7 a | 0.0010 |
| Body weight gain, kg | 72.1 ± 1.5 b | 74.0 ± 1.7 a | 0.0001 |
| ADFI, g/pig/day | 2780.3 ± 90.9 | 2835.4 ± 90.3 | 0.1900 |
| ADG, g/pig/day | 1030.3 ± 16.0 b | 1057.8 ± 10.8 a | 0.0001 |
| FCR | 2.70 ± 0.07 | 2.68 ± 0.09 | 0.6350 |
| Variable | C Group | T Group |
|---|---|---|
| Lean meat, % | 57.2 ± 0.7 b | 59.5 ± 0.7 a |
| % of cases in S class | 20.5 | 50.9 |
| % of cases in E class | 58.4 | 44.0 |
| % of cases in U class | 17.5 | 5.0 |
| % of cases in R class | 2.4 | 0.0 |
| % of cases in O class | 1.2 | 0.0 |
| % of cases in P class | 0.0 | 0.0 |
| Item, log10 CFU/g | S Group | C Group | T Group |
|---|---|---|---|
| Aerobic bacteria | 6.72 ± 0.73 a | 8.06 ± 0.46 b | 7.96 ± 0.75 b |
| Anaerobic bacteria | 9.33 ± 0.44 a | 9.39 ± 0.26 a | 9.39 ± 0.29 a |
| Lactic acid bacteria | 8.97 ± 0.32 a | 8.92 ± 0.31 a | 9.27 ± 0.18 b |
| Coliform bacteria | 5.93 ± 0.75 a | 5.39 ± 0.49 a | 5.41 ± 0.54 a |
| Clostridium perfringens | 4.05 ± 0.61 a | 4.26 ± 0.48 a | 4.27 ± 0.51 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juhász, Á.; Molnár-Nagy, V.; Bata, Z.; Tso, K.-H.; Posta, K. Phytobiotic-Prebiotic Feed Additive Containing a Combination of Carob Pulp, Chicory, and Fenugreek Improve Growth Performance, Carcass Traits, and Fecal Microbiota of Fattening Pigs. Animals 2023, 13, 3621. https://doi.org/10.3390/ani13233621
Juhász Á, Molnár-Nagy V, Bata Z, Tso K-H, Posta K. Phytobiotic-Prebiotic Feed Additive Containing a Combination of Carob Pulp, Chicory, and Fenugreek Improve Growth Performance, Carcass Traits, and Fecal Microbiota of Fattening Pigs. Animals. 2023; 13(23):3621. https://doi.org/10.3390/ani13233621
Chicago/Turabian StyleJuhász, Ákos, Viviána Molnár-Nagy, Zsófia Bata, Ko-Hua Tso, and Katalin Posta. 2023. "Phytobiotic-Prebiotic Feed Additive Containing a Combination of Carob Pulp, Chicory, and Fenugreek Improve Growth Performance, Carcass Traits, and Fecal Microbiota of Fattening Pigs" Animals 13, no. 23: 3621. https://doi.org/10.3390/ani13233621