
Phylogenetic Analyses of Lizards from the Chilean Humboldt Archipelago Reveal a New Species for the Chañaral Island (Squamata: Liolaemidae) †

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Areas of Lizard Collections
2.2. Lizard Sampling
2.3. Obtaining Mitochondrial DNA Sequences
2.4. Phylogenetic Relationships and Divergence Dates Analyses
2.5. Morphological Characters
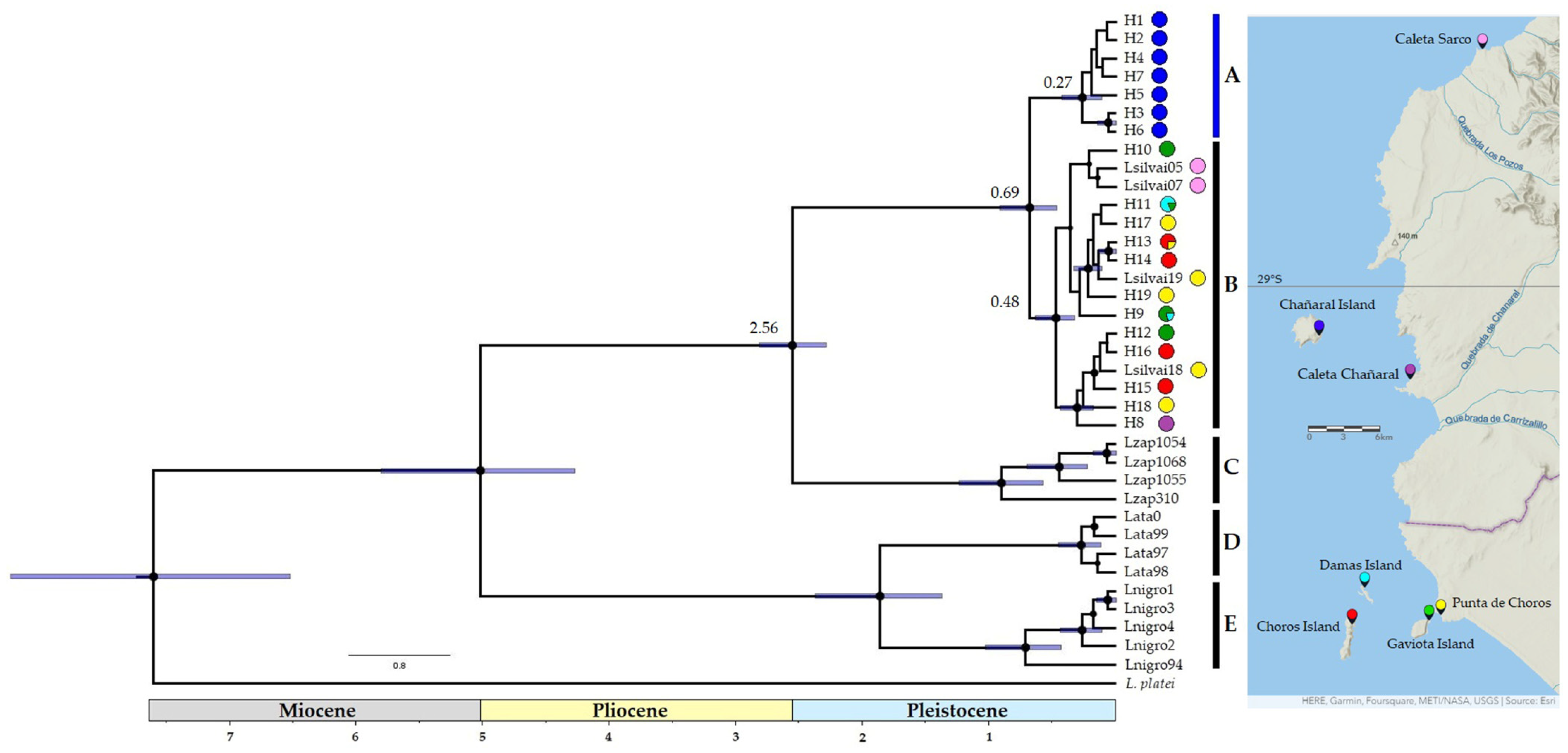
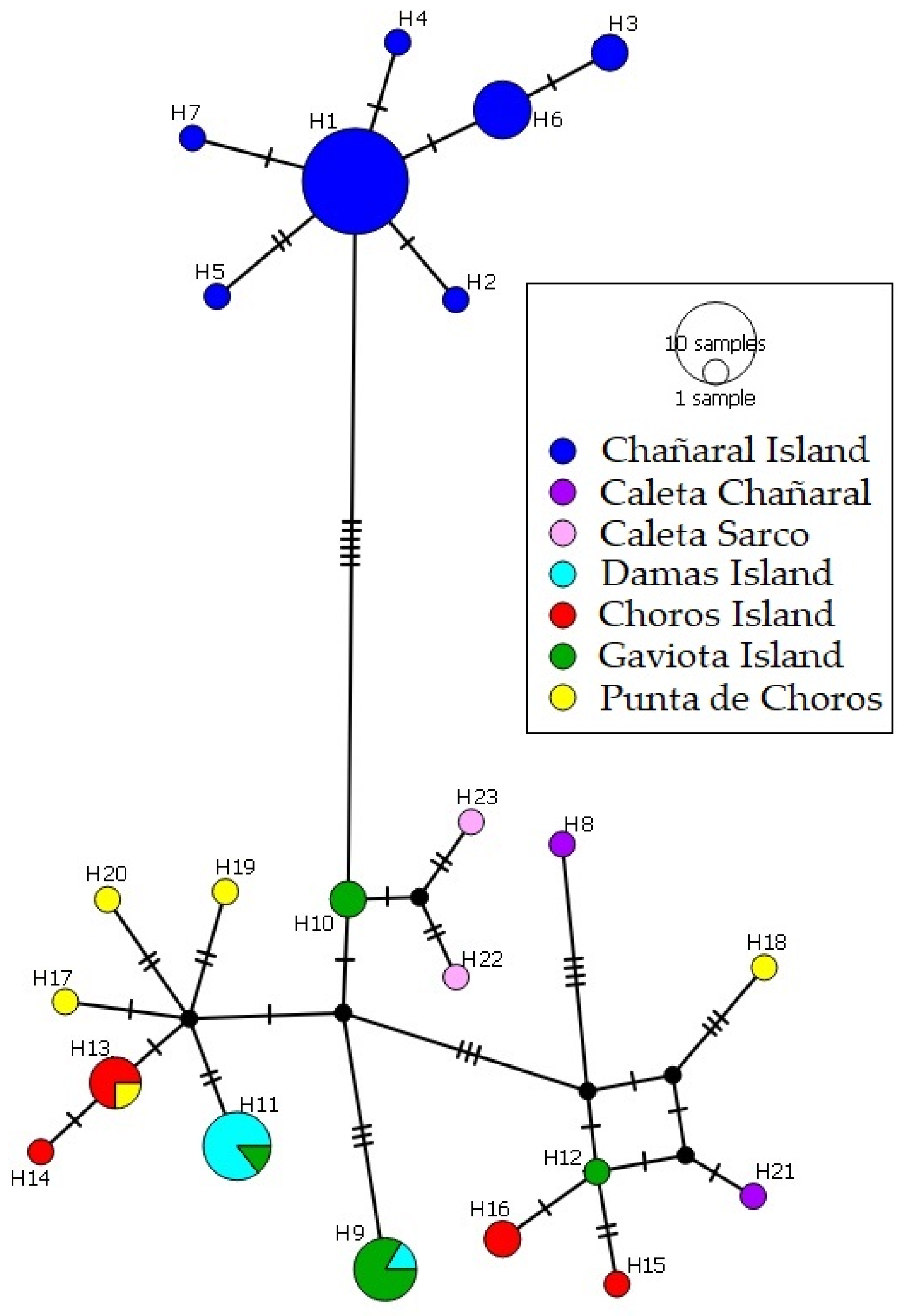
3. Results
3.1. Phylogenetic, Divergence Dates and Phylogeographic Analyses
3.2. Morphological Characteristics
3.3. Description of the New Species
Liolaemus carezzae sp. nov. CAMPOS-SOTO, CIANFERONI and TORRES-PÉREZ
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeler, R.A.; Mittermeler, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Cárcamo, P.F.; Cortés, M.; Ortega, L.; Squeo, F.A.; Gaymer, C.F. Crónica de un conflicto anunciado: Tres centrales termoeléctricas a carbón en un hotspot de biodiversidad de importancia mundial. Rev. Chil. Hist. Nat. 2011, 84, 171–180. [Google Scholar] [CrossRef]
- Luna-Jorquera, G.; Fernández, C.E.; Rivadeneira, M.M. Determinants of the diversity of plants, birds and mammals of coastal islands of the Humboldt current systems: Implications for conservation. Biodivers. Conserv. 2012, 21, 13–32. [Google Scholar] [CrossRef]
- Quispe, R.; Lerma, M.; Luna, N.; Portflitt-Toro, M.; Serratosa, J.; Luna-Jorquera, G. Foraging ranges of humboldt penguins Spheniscus humboldti from tilgo island: The critical need for protecting a unique marine habitat. Mar. Ornithol. 2020, 48, 205–208. [Google Scholar]
- Esquerré, D.; Brennan, I.G.; Catullo, R.A.; Torres-Pérez, F.; Keogh, J.S. How mountains shape biodiversity: The role of the Andes in biogeography, diversification, and reproductive biology in South America’s most species-rich lizard radiation (Squamata: Liolaemidae). Evolution 2019, 73, 214–230. [Google Scholar] [CrossRef] [PubMed]
- Díaz Gómez, J.M.; Lobo, F. Historical Biogeography of a Clade of Liolaemus (Iguania: Liolaemidae) based on ancestral areas and dispersal-vicariance analysis (DIVA). Pap. Avulsos Zool. 2006, 46, 261–274. [Google Scholar] [CrossRef]
- Schulte, J.A.; Macey, J.R.; Espinoza, R.E.; Larson, A. Phylogenetic relationships in the iguanid lizard genus Liolaemus: Multiple origins of viviparous reproduction and evidence for recurring Andean vicariance and dispersal. Biol. J. Linn. Soc. 2000, 69, 75–102. [Google Scholar] [CrossRef]
- Drummond, C.S.; Eastwood, R.J.; Miotto, S.T.S.; Hughes, C.E. Multiple Continental Radiations and Correlates of Diversification in Lupinus (Leguminosae): Testing for Key Innovation with Incomplete Taxon Sampling. Syst. Biol. 2012, 61, 443–460. [Google Scholar] [CrossRef]
- Villamil, J.; Avila, L.J.; Morando, M.; Sites, J.W.; Leaché, A.D.; Maneyro, R.; Camargo, A. Coalescent-based species delimitation in the sand lizards of the Liolaemus wiegmannii complex (Squamata: Liolaemidae). Mol. Phylogenet. Evol. 2019, 138, 89–101. [Google Scholar] [CrossRef]
- Pincheira-Donoso, D.; Núñez, H. Las especies chilenas del género Liolaemus Wiegmann, 1834 (Iguania: Tropiduridae: Liolamidae). Taxonomía, sistemática y evolución. Mus. Nac. Hist. Nat. Chile 2005, 59, 1–489. [Google Scholar]
- Vidal, M.A.; Soto, E.R.; Veloso, A. Biogeography of Chilean herpetofauna: Distributional patterns of species richness and endemism. Amphib. Reptil. 2009, 30, 151–171. [Google Scholar] [CrossRef]
- Laurent, R.F. Segunda contribución al conocimiento de la estructura taxonómica del género Liolaemus Wiegmann (Iguanidae). Cuad. Herpetol. 1985, 1, 1–37. [Google Scholar]
- Troncoso-Palacios, J.; Schulte, J.A.; Marambio-Alfaro, Y.; Hiriart, D. Phenotypic Variation, Phylogenetic Position and New Distributional Records for the Poorly Known Liolaemus silvai Ortiz, 1989 (Iguania: Iguanidae: Liolaemini). S. Am. J. Herpetol. 2015, 10, 71. [Google Scholar] [CrossRef]
- Esquerré, D.; Keogh, J.S.; Demangel, D.; Morando, M.; Avila, L.J.; Sites, J.W.; Ferri-Yáñez, F.; Leaché, A.D. Rapid Radiation and Rampant Reticulation: Phylogenomics of South American Liolaemus Lizards. Syst. Biol. 2021, 71, 286–300. [Google Scholar] [CrossRef]
- Demangel, D. Reptiles en Chile; Fauna Nativa Ediciones: Santiago, Chile, 2016; ISBN 978-956-9801-00-6. [Google Scholar]
- ArcGIS Online. Available online: https://www.arcgis.com/index.html (accessed on 17 April 2023).
- Beaupre, S.; Jacobson, E.; Lillywhite, H.; Zamudio, K. Guidelines for Use of Live Amphibians and Reptiles in Field and Laboratory Research, 2nd ed.; American Society of Ichthyologists and Herpetologists: Lawrence, KS, USA, 2004. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Fontanella, F.M.; Olave, M.; Avila, L.J.; Sites, J.W.; Morando, M. Molecular dating and diversification of the South American lizard genus Liolaemus (subgenus Eulaemus) based on nuclear and mitochondrial DNA sequences. Zool. J. Linn. Soc. 2012, 164, 825–835. [Google Scholar] [CrossRef]
- Maturana, P.; Brewer, B.J.; Klaere, S.; Bouckaert, R. Model selection and parameter inference in phylogenetics using Nested Sampling. Syst. Biol. 2019, 68, 219–233. [Google Scholar] [CrossRef]
- Kass, R.E.; Raftery, A.E. Bayes Factors. J. Am. Stat. Assoc. 1995, 90, 773–795. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Ortiz, J.C. Description de Liolaemus silvai sp. nov. (Sauria, Iguanidae) du «Norte Chico» du Chili. Bull. Mus. Natn. Hist. Nat. Paris 1989, 11, 247–252. [Google Scholar]
- Etheridge, R. A review of lizards of the Liolaemus wiegmannii group (Squamata, Iguania, Tropiduridae), and a history of morphological change in the sand-dwelling species. Herpetol. Monogr. 2000, 14, 293–352. [Google Scholar] [CrossRef]
- Avila, L.J.; Perez, C.H.F.; Morando, M.; Sites, J.W. A new species of Liolaemus (Reptilia: Squamata) from southwestern Rio Negro province, northern Patagonia, Argentina. Zootaxa 2010, 2434, 47–59. [Google Scholar] [CrossRef]
- Troncoso-Palacios, J.; Contreras-Piderit, F. Una nueva especie del grupo de Liolaemus nigroviridis de los Andes de Chile central (Iguania: Liolaemidae). Acta Zoológica Lilloana 2023, 67, 233–259. [Google Scholar] [CrossRef]
- Etheridge, R. Redescription of Ctenoblepharys adspersa Tschudi, 1845, and the Taxonomy of Liolaeminae (Reptilia: Squamata: Tropiduridae). Am. Mus. Novit. 1995, 3142, 1–34. [Google Scholar]
- Botto-Mahan, C.; Correa, J.P.; Araya-Donoso, R.; Farías, F.; Juan, E.S.; Quiroga, N.; Campos-Soto, R.; Reyes-Olivares, C.; González-Acuña, D. Lizards as Silent Hosts of Trypanosoma cruzi. Emerg. Infect. Dis. 2022, 28, 1250–1253. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, K.; Staley, C.; Eifler, M.A.; Núñez, H.; Eifler, D.A. Color Variation and Habitat Use in Liolaemus silvai. S. Am. J. Herpetol. 2021, 21, 80–84. [Google Scholar] [CrossRef]
- Grismado, C.J.; Pizarro-Araya, J. The spider genus Cyrioctea Simon on Chañaral Island (Pingüino de Humboldt National Reserve, Atacama, Chile): Description of a new species, and the male of Cyrioctea cruz Platnick (Araneae, Zodariidae). Zootaxa 2016, 4107, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Morrone, J.J. Biogeographical regionalisation of the Andean region. Zootaxa 2015, 3936, 207–236. [Google Scholar] [CrossRef]
- Ceccarelli, F.S.; Pizarro-Araya, J.; Ojanguren-Affilastro, A.A. Phylogeography and population structure of two Brachistosternus species (Scorpiones: Bothriuridae) from the Chilean coastal desert—The perils of coastal living. Biol. J. Linn. Soc. 2017, 120, 75–89. [Google Scholar] [CrossRef]
- Quezada, J.; González, G.; Dunai, T.; Jensen, A.; Juez-Larré, J. Alzamiento litoral Pleistoceno del norte de Chile: Edades 21Ne de la terraza costera más alta del área de Caldera-Bahía Inglesa. Rev. Geológica Chile 2007, 34, 81–96. [Google Scholar] [CrossRef]
- Pedoja, K.; Husson, L.; Johnson, M.E.; Melnick, D.; Witt, C.; Pochat, S.; Nexer, M.; Delcaillau, B.; Pinegina, T.; Poprawski, Y.; et al. Earth-Science Reviews Coastal staircase sequences reflecting sea-level oscillations and tectonic uplift during the Quaternary and Neogene. Earth Sci. Rev. 2014, 132, 13–38. [Google Scholar] [CrossRef]
- Ortlieb, L.; Zazo, C.; Goy, J.L.; Hillaire-Marcel, C.; Ghaleb, B.; Cournoyer, L. Coastal deformation and sea-level changes in the northern Chile subduction area (23 degrees S) during the last 330 ky. Quat. Sci. Rev. 1996, 15, 819–831. [Google Scholar] [CrossRef]
- Campos-Soto, R.; Díaz-Campusano, G.; Cianferoni, F.; Torres-Pérez, F. Biogeographic origin and phylogenetic relationships of Mepraia (Hemiptera, Reduviidae) on islands of northern Chile. PLoS ONE 2020, 15, e0234056. [Google Scholar] [CrossRef]
- Campos-Soto, R.; Rodríguez-Valenzuela, E.; Díaz-Campusano, G.; Boric-Bargetto, D.; Zúñiga-Reinoso, Á.; Cianferoni, F.; Torres-Pérez, F. Testing Phylogeographic Hypotheses in Mepraia (Hemiptera: Reduviidae) Suggests a Complex Spatio-Temporal Colonization in the Coastal Atacama Desert. Insects 2022, 13, 419. [Google Scholar] [CrossRef] [PubMed]
- Lobo, F.; Espinoza, R.E.; Quinteros, S. A critical review and systematic discussion of recent classification proposals for Liolaemid lizards. Zootaxa 2010, 2549, 1–30. [Google Scholar] [CrossRef]
- Troncoso-Palacios, J.; Garin, C.F. On the identity of Liolaemus nigromaculatus Wiegmann, 1834 (Iguania, Liolaemidae) and correction of its type locality. Zookeys 2013, 294, 37–56. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Latitude/Longitude | Species | N | H |
|---|---|---|---|---|
| Chañaral Island | 29°2′17″ S/71°34′8″ W | L. carezzae sp. nov. | 28 | 7 |
| Caleta Chañaral | 29°4′14″ S/71°29′16″ W | L. silvai | 1 | 1 |
| Damas Island | 29°13′48″ S/71°31′41″ W | L. silvai | 7 | 2 |
| Choros Island | 29°15′32″ S/71°32′19″ W | L. silvai | 7 | 4 |
| Gaviota Island | 29°15′24″ S/71°28′16″ W | L. silvai | 9 | 4 |
| Punta de Choros | 29°15′4″ S/71°27′40″ W | L. silvai | 4 | 4 |
| Los Molles | 32°14′29″ S/71°31′7″ W | L. zapallarensis | 3 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos-Soto, R.; Rodríguez-Valenzuela, E.; Bruna, Y.; Díaz-Campusano, G.; Cianferoni, F.; Boric-Bargetto, D.; Torres-Pérez, F. Phylogenetic Analyses of Lizards from the Chilean Humboldt Archipelago Reveal a New Species for the Chañaral Island (Squamata: Liolaemidae). Animals 2023, 13, 3576. https://doi.org/10.3390/ani13223576
Campos-Soto R, Rodríguez-Valenzuela E, Bruna Y, Díaz-Campusano G, Cianferoni F, Boric-Bargetto D, Torres-Pérez F. Phylogenetic Analyses of Lizards from the Chilean Humboldt Archipelago Reveal a New Species for the Chañaral Island (Squamata: Liolaemidae). Animals. 2023; 13(22):3576. https://doi.org/10.3390/ani13223576
Chicago/Turabian StyleCampos-Soto, Ricardo, Evelyn Rodríguez-Valenzuela, Yareta Bruna, Gabriel Díaz-Campusano, Franco Cianferoni, Dusan Boric-Bargetto, and Fernando Torres-Pérez. 2023. "Phylogenetic Analyses of Lizards from the Chilean Humboldt Archipelago Reveal a New Species for the Chañaral Island (Squamata: Liolaemidae)" Animals 13, no. 22: 3576. https://doi.org/10.3390/ani13223576