
Effect of Food Restriction on Food Grinding in Brandt’s Voles

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Experimental Design
2.2.1. 16S rRNA Gene Sequencing and Bioinformatic Analysis of Fecal Microbiota
2.2.2. Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Prediction of Fecal Microbiota
2.2.3. SCFA Assay
2.2.4. Statistical Analysis
3. Results
3.1. Differences in Food Intake, Food Ground, and Body Mass Growth Rate
3.2. OTUs Analysis
3.3. Differences in Alpha and Beta Diversities of Fecal Microbial Community
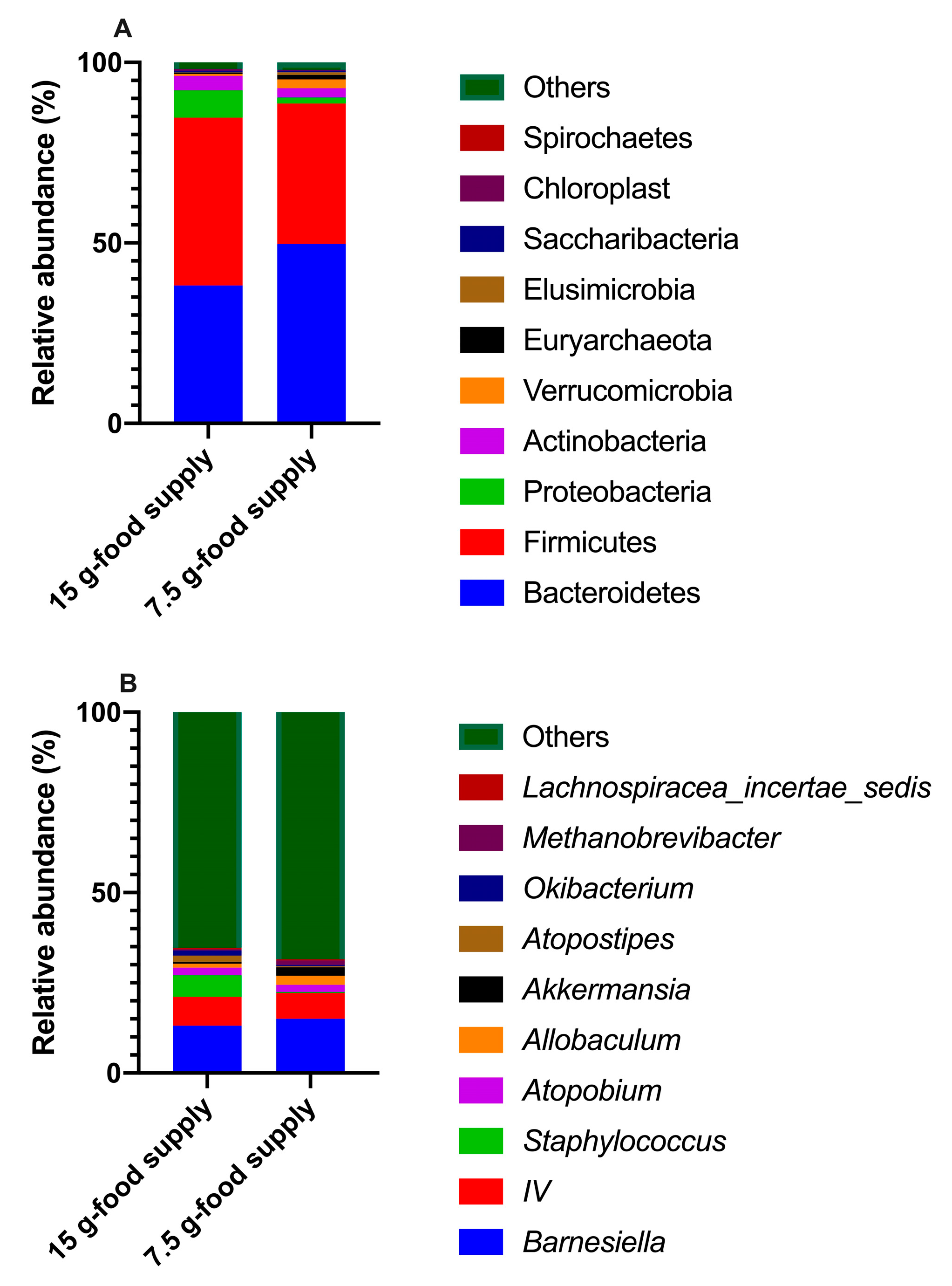
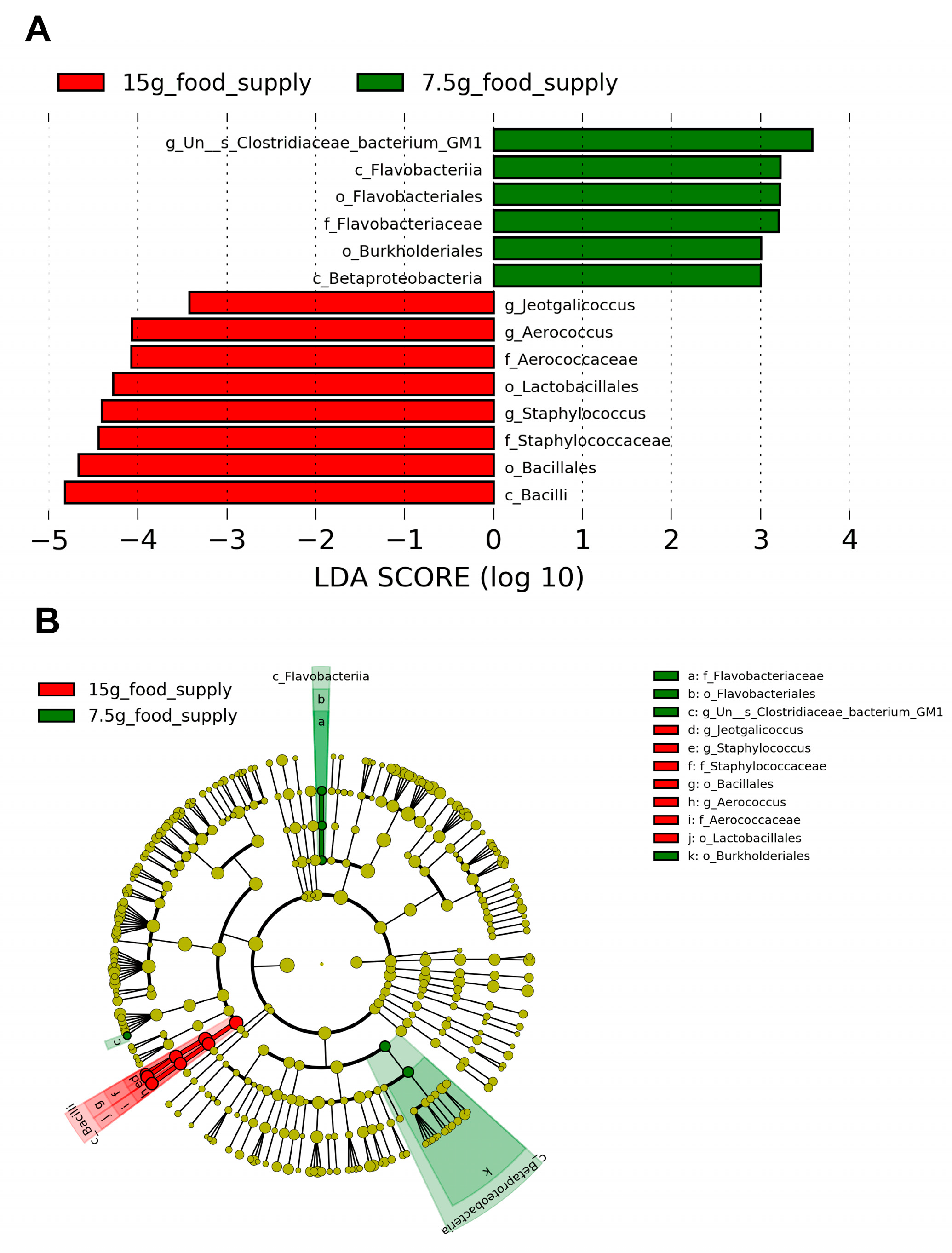
3.4. Differences in the Abundances of the Enriched Biomarkers of Fecal Microbial Community
3.5. The Enriched KEGG Pathways of Fecal Microbial Community and Differences in the Content of Fecal SCFAs
3.6. Spearman’s Rank Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron, K.M.; Speakman, J.R. The extent and function of ‘food grinding’ in the laboratory mouse (Mus musculus). Lab. Anim. 2010, 44, 298–304. [Google Scholar] [CrossRef]
- Starr, M.E.; Saito, H. Age-related increase in food spilling by laboratory mice may lead to significant overestimation of actual food consumption: Implications for studies on dietary restriction, metabolism, and dose calculations. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Tertil, R. Impact of the Common Vole, Microtus arvalis (Pallas) on Winter Wheat and Alfalfa Crops. EPPO Bull. 1977, 7, 317–339. [Google Scholar] [CrossRef]
- Koteja, P.; Carter, P.A.; Swallow, J.G.; Garland, T., Jr. Food wasting by house mice: Variation among individuals, families, and genetic lines. Physiol. Behav. 2003, 80, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.; Wunder, B.A. Food Sorting by Collared Lemmings (Dicrostonyx groenlandicus) and Prairie Voles (Microtus ochrogaster): A Cautionary Note for Digestibility Studies. Comp. Biochem. Physiol. A Physiol. 1997, 116, 119–124. [Google Scholar] [CrossRef]
- Ford, D.J. Influence of diet pellet hardness and particle size on food utilization by mice, rats and hamsters. Lab. Anim. 1977, 11, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Martins, P.J.F.; Nobrega, J.N.; Tufik, S.; D’Almeida, V. Sleep deprivation-induced gnawing—Relationship to changes in feeding behavior in rats. Physiol. Behav. 2008, 93, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Esumi, L.A.; Palma, B.D.; Gomes, A.C.; Tufik, S.; Hipólide, D.C. Inflammatory markers are associated with inhibitory avoidance memory deficit induced by sleep deprivation in rats. Behav. Brain Res. 2011, 221, 7–12. [Google Scholar] [CrossRef]
- Zhao, Z.J.; Chi, Q.S.; Zhao, L.; Zhu, Q.X.; Cao, J.; Wang, D.H. Effect of food restriction on energy budget in warm-acclimated striped hamsters. Physiol. Behav. 2015, 147, 220–226. [Google Scholar] [CrossRef]
- Xie, X.M.; Sun, R.Y.; Fang, J.M. The mating system and reproduction of Brandt’s voles (Microtus brandtii). Acta Zool. Sin. 1994, 40, 262–265. [Google Scholar]
- Shi, D.Z.; Hai, S.Z.; Lu, D.; Liu, X.F. The structure and order in colony of Brandt’s vole. Acta Theriol. Sin. 1999, 19, 48–55. [Google Scholar]
- Wan, X.R.; Wang, M.J.; Wang, G.H.; Liu, W.; Zhong, W.Q. The reproductive parameters in the marked populations of Brandt’s vole. Acta Theriol. Sin. 2002, 22, 116–122. [Google Scholar]
- Li, G.; Yin, B.; Li, J.; Wang, J.; Wei, W.; Bolnick, D.I.; Wan, X.; Zhu, B.; Zhang, Z. Host-microbiota interaction helps to explain the bottom-up effects of climate change on a small rodent species. ISME J. 2020, 14, 1795–1808. [Google Scholar] [CrossRef]
- Li, G.; Li, J.; Kohl, K.D.; Yin, B.; Wei, W.; Wan, X.; Zhu, B.; Zhang, Z. Dietary shifts influenced by livestock grazing shape the gut microbiota composition and co-occurrence networks in a local rodent species. J. Anim. Ecol. 2019, 88, 302–314. [Google Scholar] [CrossRef]
- Wang, G.M.; Zhou, Q.Q.; Zhong, W.Q.; Wang, G.H. Food habits of Brandt’s vole (Microtus brandtii). Acta Theriol. Sin. 1992, 12, 57–64. [Google Scholar]
- Wan, X.R.; Wang, G.H.; Zhong, W.Q. Estimating the Non-Feeding Grass Consumption of Brandts Vole. Chin. J. Ecol. 2001, 20, 77–78. [Google Scholar]
- Shen, Q.; Shi, J.; Gu, K.; Wei, W.; Yang, S.; Dai, X. Relationship between food grinding and gut microbiota in Brandt’s voles. Can. J. Zool. 2023, 101, 623–634. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, L.; Li, Z.; Wang, D.; Li, N.; Song, Y.; Guo, C.; Liu, X. Molecular cloning and characterization of kiss1 in Brandt’s voles (Lasiopodomys brandtii). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2017, 208–209, 68–74. [Google Scholar] [CrossRef]
- Kelley, T.L. The selection of upper and lower groups for the validation of test items. J. Educ. Psychol. 1939, 30, 17–24. [Google Scholar] [CrossRef]
- Dai, X.; Chen, L.; Liu, M.; Liu, Y.; Jiang, S.; Xu, T.; Wang, A.; Yang, S.; Wei, W. Effect of 6-Methoxybenzoxazolinone on the Cecal Microbiota of Adult Male Brandt’s Vole. Front. Microbiol. 2022, 13, 847073. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Campos, P.M.; Darwish, N.; Shao, J.; Proszkowiec-Weglarz, M. Research Note: Choice of microbiota database affects data analysis and interpretation in chicken cecal microbiota. Poult. Sci. 2022, 101, 101971. [Google Scholar] [CrossRef]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Dovrou, A.; Bei, E.; Sfakianakis, S.; Marias, K.; Papanikolaou, N.; Zervakis, M. Synergies of Radiomics and Transcriptomics in Lung Cancer Diagnosis: A Pilot Study. Diagnostics 2023, 13, 738. [Google Scholar] [CrossRef]
- Bo, T.B.; Zhang, X.Y.; Wen, J.; Deng, K.; Qin, X.W.; Wang, D.H. The microbiota-gut-brain interaction in regulating host metabolic adaptation to cold in male Brandt’s voles (Lasiopodomys brandtii). ISME J. 2019, 13, 3037–3053. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, Z. Sex- and age-specific variation of gut microbiota in Brandt’s voles. PeerJ 2021, 9, e11434. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Kumar, M.; Baker, M.T.; Singh, V.; Vijay-Kumar, M. Mammalian gut immunity. Biomed. J. 2014, 37, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Lavrinienko, A.; Tukalenko, E.; Kesaniemi, J.; Kivisaari, K.; Masiuk, S.; Boratynski, Z.; Mousseau, T.A.; Milinevsky, G.; Mappes, T.; Watts, P.C. Applying the Anna Karenina principle for wild animal gut microbiota: Temporal stability of the bank vole gut microbiota in a disturbed environment. J. Anim. Ecol. 2020, 89, 2617–2630. [Google Scholar] [CrossRef]
- Steketee, J.K.; Rocha, A.V.; Gough, L.; Griffin, K.L.; Klupar, I.; An, R.; Williamson, N.; Rowe, R.J. Small herbivores with big impacts: Tundra voles (Microtus oeconomus) alter post-fire ecosystem dynamics. Ecology 2022, 103, e3689. [Google Scholar] [CrossRef]
- Pritchett-Corning, K.R.; Keefe, R.; Garner, J.P.; Gaskill, B.N. Can seeds help mice with the daily grind? Lab. Anim. 2013, 47, 312–315. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Mao, H.; Ruan, Y.; Wu, C.; Xu, Z.; Hu, G.; Guo, X.; Zhang, C.; Cao, H.; Liu, P. Copper Changes Intestinal Microbiota of the Cecum and Rectum in Female Mice by 16S rRNA Gene Sequencing. Biol. Trace Elem. Res. 2020, 193, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Ruzauskas, M.; Armalyte, J.; Lastauskiene, E.; Siugzdiniene, R.; Klimiene, I.; Mockeliunas, R.; Bartkiene, E. Microbial and Antimicrobial Resistance Profiles of Microbiota in Common Carps (Cyprinus carpio) from Aquacultured and Wild Fish Populations. Animals 2021, 11, 929. [Google Scholar] [CrossRef]
- Narita, K.; Asano, K.; Nakane, A. IL-17A plays an important role in protection induced by vaccination with fibronectin-binding domain of fibronectin-binding protein A against Staphylococcus aureus infection. Med. Microbiol. Immunol. 2017, 206, 225–234. [Google Scholar] [CrossRef]
- Stokes, W.; Parkins, M.D.; Parfitt, E.C.T.; Ruiz, J.C.; Mugford, G.; Gregson, D.B. Incidence and Outcomes of Staphylococcus aureus Bacteriuria: A Population-based Study. Clin. Infect. Dis. 2019, 69, 963–969. [Google Scholar] [CrossRef]
- Sethi, S.; Chakraborty, T. Role of TLR-/NLR-signaling and the associated cytokines involved in recruitment of neutrophils in murine models of Staphylococcus aureus infection. Virulence 2011, 2, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.A.; Fernandes, A.; Dias, M.I.; Pereira, C.; Calhelha, R.C.; Chrysargyris, A.; Tzortzakis, N.; Ivanov, M.; Sokovic, M.D.; Barros, L.; et al. Chemical Composition and Plant Growth of Centaurea raphanina subsp. mixta Plants Cultivated under Saline Conditions. Molecules 2020, 25, 2204. [Google Scholar] [CrossRef]
- Sado-Kamdem, S.L.; Vannini, L.; Guerzoni, M.E. Effect of alpha-linolenic, capric and lauric acid on the fatty acid biosynthesis in Staphylococcus aureus. Int. J. Food Microbiol. 2009, 129, 288–294. [Google Scholar] [CrossRef]
- Munoz-Sanchez, S.; van der Vaart, M.; Meijer, A.H. Autophagy and Lc3-Associated Phagocytosis in Zebrafish Models of Bacterial Infections. Cells 2020, 9, 2372. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Han, S.; Wang, P.; Wen, B.; Jian, W.; Guo, W.; Yu, Z.; Du, D.; Fu, X.; Kong, F.; et al. The bacterial communities associated with fecal types and body weight of rex rabbits. Sci. Rep. 2015, 5, 9342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Guo, Z.; Xue, Z.; Sun, Z.; Zhang, M.; Wang, L.; Wang, G.; Wang, F.; Xu, J.; Cao, H.; et al. A phylo-functional core of gut microbiota in healthy young Chinese cohorts across lifestyles, geography and ethnicities. ISME J. 2015, 9, 1979–1990. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Total Tags | Taxon Tags | OTU Numbers | Goods Coverage |
|---|---|---|---|---|
| 15 g-food supply | 63,501 ± 1070 | 57,234 ± 1017 | 877 ± 47 | 99.77 ± 0.02 |
| 7.5 g-food supply | 66,282 ± 1567 | 57,704 ± 2233 | 981 ± 22 | 99.75 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, X.; Han, Y.-X.; Shen, Q.-Y.; Tang, H.; Cheng, L.-Z.; Yang, F.-P.; Wei, W.-H.; Yang, S.-M. Effect of Food Restriction on Food Grinding in Brandt’s Voles. Animals 2023, 13, 3424. https://doi.org/10.3390/ani13213424
Dai X, Han Y-X, Shen Q-Y, Tang H, Cheng L-Z, Yang F-P, Wei W-H, Yang S-M. Effect of Food Restriction on Food Grinding in Brandt’s Voles. Animals. 2023; 13(21):3424. https://doi.org/10.3390/ani13213424
Chicago/Turabian StyleDai, Xin, Yu-Xuan Han, Qiu-Yi Shen, Hao Tang, Li-Zhi Cheng, Feng-Ping Yang, Wan-Hong Wei, and Sheng-Mei Yang. 2023. "Effect of Food Restriction on Food Grinding in Brandt’s Voles" Animals 13, no. 21: 3424. https://doi.org/10.3390/ani13213424