
Changes in Growth and Feeding Characteristics during Early Ontogenesis in Threadsail Filefish, Stephanolepis cirrhifer
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Measurement of Growth Indices
2.3. Yolk Sac and Oil Globule Volume
2.4. Satiety, Satiety Rate, Feeding Rate, and PNR
2.5. Diel Feeding Rhythm
2.6. Image Processing and Statistical Analysis
3. Results
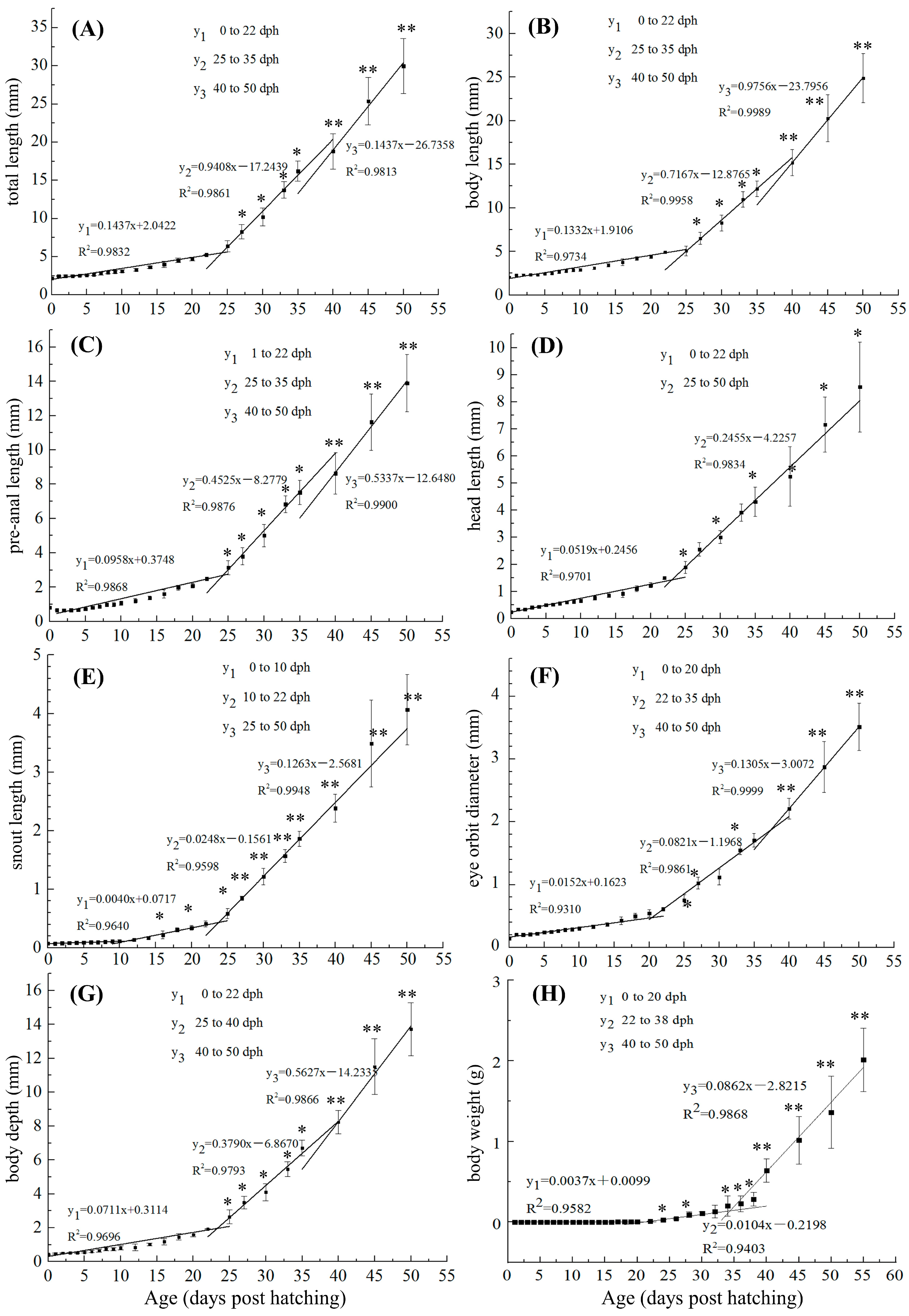
3.1. Changes in Growth Indicators from 0 to 50 dph in S. cirrhifer
3.2. Feeding Rate, Satiety, and Satiety Rate
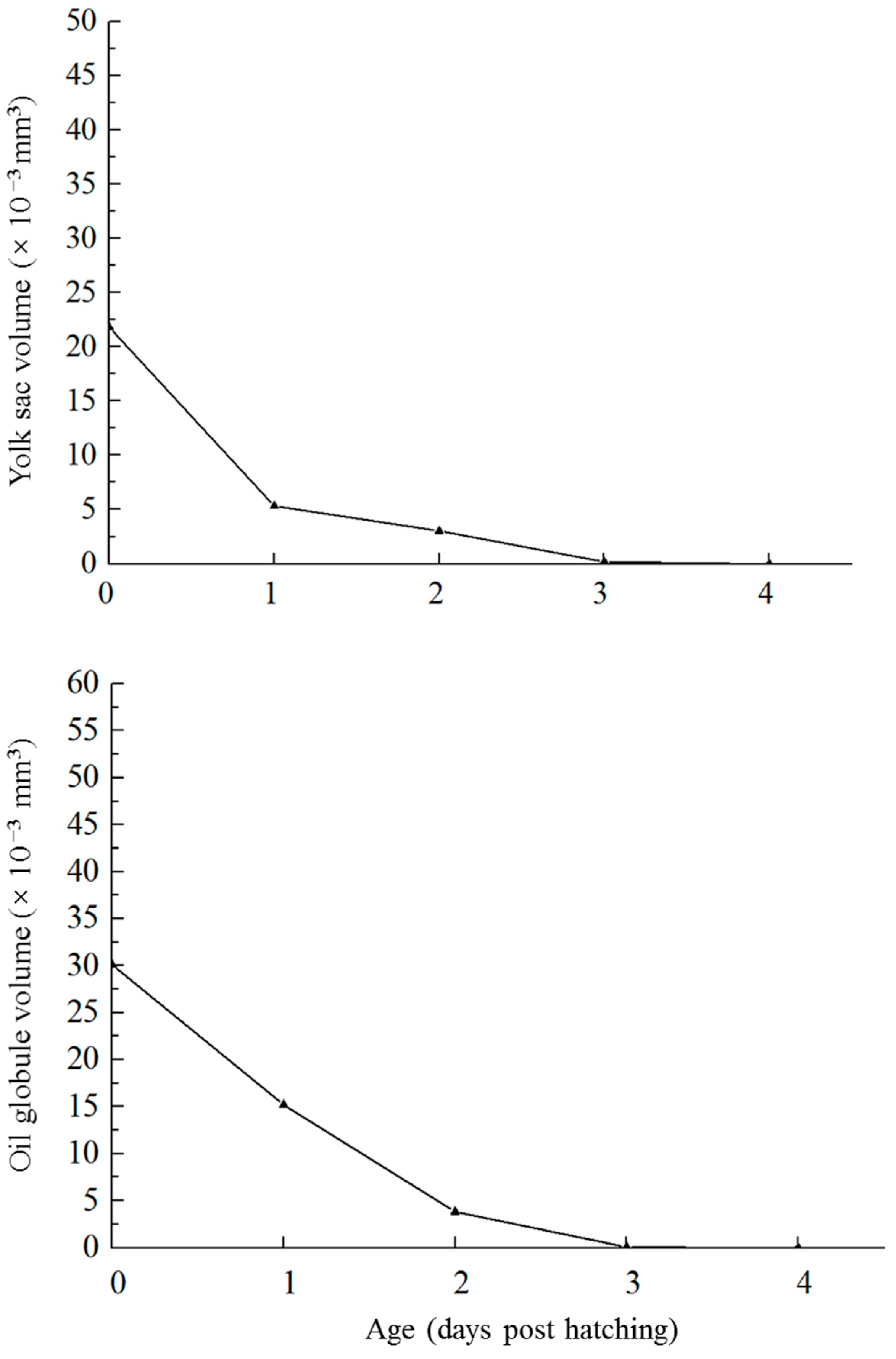
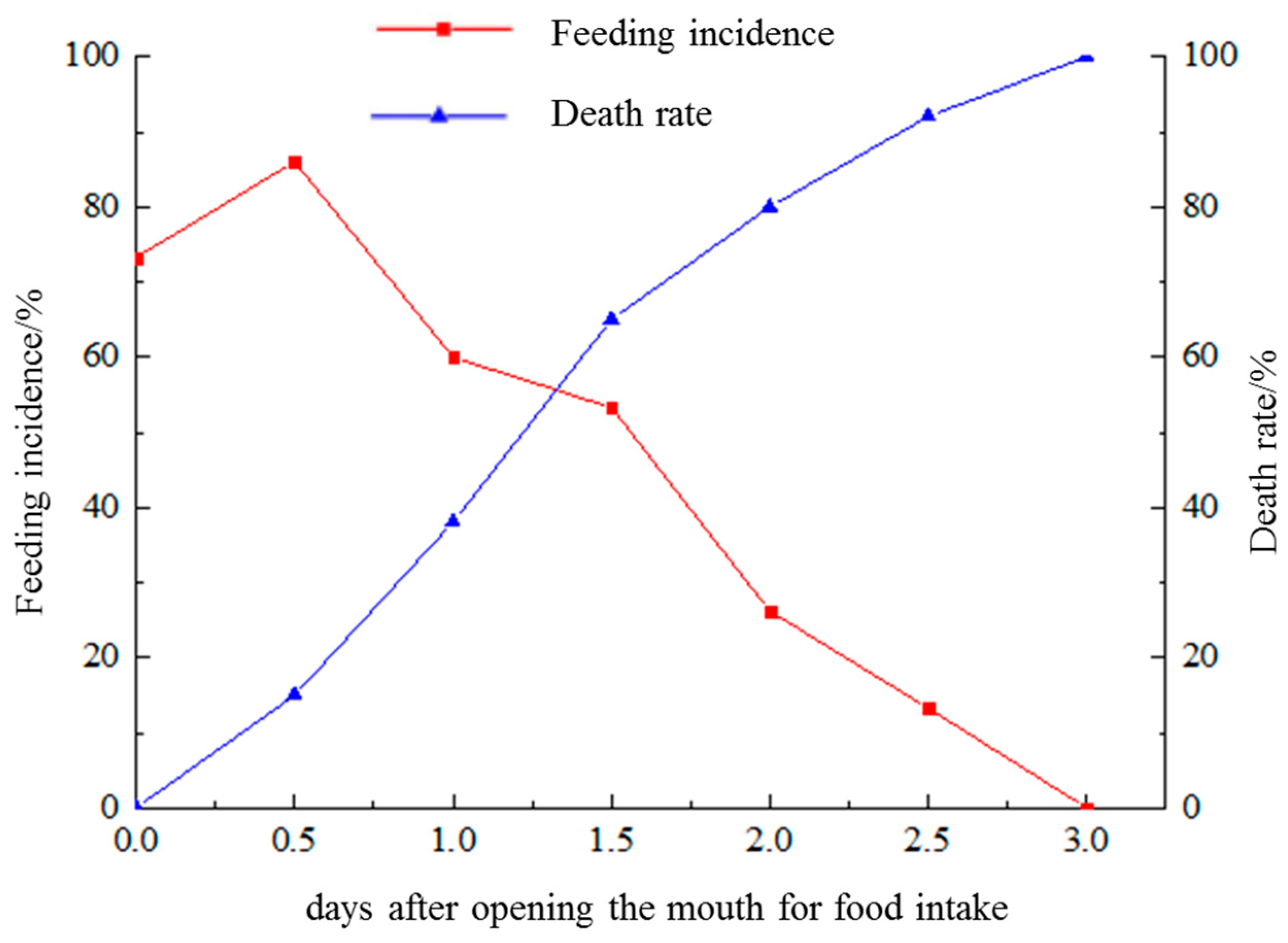
3.3. Absorption of Endogenous Nutrients and PNR
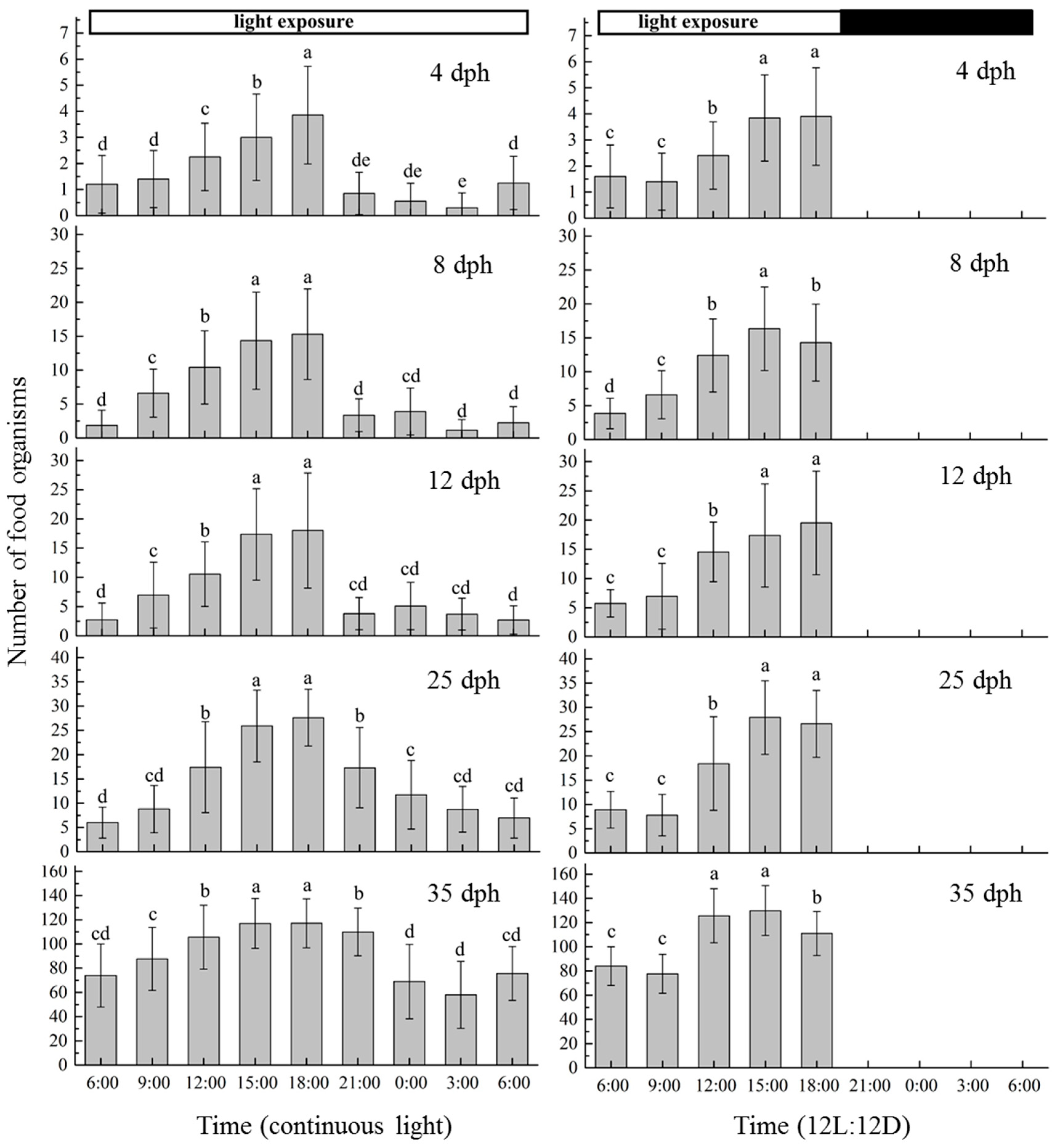
3.4. Diel Feeding Rhythm
4. Discussion
4.1. Growth Indices of the Early Developmental Phase
4.2. Mixed Feeding Stage, Yolk-Sac Absorption, and PNR during Early Development
4.3. Feeding Rhythms of Early Development Stage
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mizuno, K.; Miura, C.; Miura, T. Relationships between temperature and growth of Thread-sail Filefish Stephanolepis cirrhifer and Black Scraper Thamnaconus modestus. Aquacult. Sci. 2014, 62, 23–30. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, N.Y.; Kim, S.H.; Cho, J.I.; Lee, H.J.; Ha, S.D. Effect of ultraviolet radiation on the reduction of major food spoilage molds and sensory quality of the surface of dried filefish (Stephanolepis cirrhifer) fillets. Food Res. Int. 2014, 62, 1108–1112. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, P.M.; Chen, G.B.; Fang, L.C.; Tang, Y. Acoustic target strength measurement of banded grouper (Epinephelus awoara (Temming and Schlegel, 1842)) and threadsial filefish (Stephanolepis cirrhifer (Temming & Schlegel, 1850)) in the South China Sea. J. Appl. Ichthyol. 2014, 29, 1453–1455. [Google Scholar] [CrossRef]
- Seoul Ministry for Food, Agriculture, Forestry and Fisheries. Annual Statistics of Fisheries Production in 1990–2009; Seoul Ministry for Food, Agriculture, Forestry and Fisheries: Seoul, Republic of Korea, 2009. [Google Scholar]
- IUCN 2017. The IUCN Red List of Threatened Species. Version 18 July 2017. Available online: https://www.iucnredlist.org (accessed on 15 May 2023).
- Markevich, A.I.; Balanov, A.A. Finding of threadsail filefish Stephanolepis cirrhifer (Temminck et Schlegel, 1850) rare for Peter the Great Bay, Sea of Japan. J. Ichthyol. 2011, 51, 678–682. [Google Scholar] [CrossRef]
- Allen, J.J.; Akkaynak, D.; Sugden, A.U.; Hanlon, R.T. Adaptive body patterning, three-dimensional skin morphology, and camouflage measures of the slender filefish Monacanthus tuckeri on a Caribbean coral reef. Biol. J. Linn. Soc. 2015, 116, 377–396. [Google Scholar] [CrossRef]
- Hossain, M.A.; Furuichi, M. Necessity of dietary calcium supplement in file fish (Monacanthus cirrhifer). Bangladesh J. Fish. Res. 2008, 12, 157–162. Available online: http://hdl.handle.net/1834/34223 (accessed on 7 August 2023).
- Khosravi, S.; Lee, S.M. Optimum dietary protein and lipid levels in juvenile filefish, Stephanolepis cirrhifer, feed. J. World Aquacult. Soc. 2017, 48, 867–876. [Google Scholar] [CrossRef]
- Miyajima, Y.; Masuda, R.; Kurihara, A.; Kamata, R.; Yamashita, Y.; Takeuchi, T. Juveniles of threadsail filefish, Stephanolepis cirrhifer, can survive and grow by feeding on moon jellyfish Aurelia aurita. Fish. Sci. 2011, 77, 41–48. [Google Scholar] [CrossRef]
- Løkkeborg, S.; Siikavuopio, S.I.; Humborstad, O.B.; Utne-Palm, A.C.; Ferter, K. Towards more efficient longline fisheries: Fish feeding behaviour, bait characteristics and development of alternative baits. Rev. Fish Biol. Fish. 2014, 24, 985–1003. [Google Scholar] [CrossRef]
- Moteki, M.; Ishikawa, T.; Teraoka, N.; Fushimi, H. Transition from endogenous to exogenous nutritional sources in larval red sea bream, Pagrus major. Aquacult. Sci. 2001, 49, 323–328. [Google Scholar] [CrossRef]
- Wittenrich, M.L.; Rhody, N.R.; Turingan, R.G.; Main, K.L. Coupling osteological development of the feeding apparatus with feeding performance in common snook, Centropomus undecimalis, larvae: Identifying morphological constraints to feeding. Aquaculture 2009, 294, 221–227. [Google Scholar] [CrossRef]
- López-Olmeda, J.F.; Noble, C.; Sánchez-Vázquez, F.J. Does feeding time affect fish welfare? Fish Physiol. Biochem. 2012, 38, 143–152. [Google Scholar] [CrossRef]
- Ma, Z. Food ingestion, prey selectivity, feeding incidence, and performance of yellowtail kingfish Seriola lalandi larvae under constant and varying temperatures. Aquacult. Int. 2014, 22, 1317–1330. [Google Scholar] [CrossRef]
- de Mattos, B.O.; Sousa, E.D.S.; Saboia, W.A.; Santana, T.M.; Corrêa, M.A.D.A.; Sánchez-Vázquez, F.J.; Fortes-Silva, R. Effects of three feeding strategies (self-feeders, automatic feeding and apparent satiety) on growth performance, haematological parameters, waste excretion and feed cost in Amazonian fish (Colossoma macropomum). Aquac. Res. 2022, 53, 4531–4539. [Google Scholar] [CrossRef]
- Blaxter, J.H.; Hempel, G. The influence of egg size on herring larvae (Clupea harengus L.). ICES J. Mar. Sci. 1963, 28, 211–240. [Google Scholar] [CrossRef]
- Chai, Y.; Xie, C.; Wei, Q.W. Yolk-sac absorption and point of no return in Chinese sturgeon Acipenser sinensis larvae. J. Appl. Ichthyol. 2011, 27, 687–689. [Google Scholar] [CrossRef]
- Xu, Z.; Li, C.; Ling, Q.; Gaughan, S.; Wang, G.; Han, X. Early development and the point of no return in pikeperch (Sander lucioperca L.) larvae. Chin. J. Oceanol. Limn. 2017, 35, 1493–1500. [Google Scholar] [CrossRef]
- Garcia, L.M.B.; Sayco, M.J.P.; Aya, F.A. Point-of-no-return and delayed feeding mortalities in first-feeding larvae of the silver therapon Leiopotherapon plumbeus (Kner) (Teleostei: Terapontidae). Aquacult. Rep. 2020, 16, 100288. [Google Scholar] [CrossRef]
- Ahmed, I.; Reshi, Q.M.; Fazio, F. The influence of the endogenous and exogenous factors on hematological parameters in different fish species: A review. Aquacult. Int. 2020, 28, 869–899. [Google Scholar] [CrossRef]
- May, R.C. Larval mortality in marine fishes and the critical period concept. In The Early Life History of Fish: The Proceedings of an International Symposium Held at the Dunstaffnage Marine Research Laboratory of the Scottish Marine Biological Association at Oban, Scotland, from May 17–23, 1973; Springer: Berlin/Heidelberg, Germany, 1974; pp. 3–19. [Google Scholar] [CrossRef]
- Blaxter, J.H.S. 1 Pattern and variety in development. In Fish Physiology; Academic Press: Cambridge, MA, USA, 1988; pp. 1–58. [Google Scholar] [CrossRef]
- Otterlei, E.; Nyhammer, G.; Folkvord, A.; Stefansson, S.O. Temperature- and size-dependent growth of larval and early juvenile Atlantic cod (Gadus morhua): A comparative study of Norwegian coastal cod and northeast Arctic cod. Can. J. Fish. Aquat. Sci. 1999, 56, 2099–2111. [Google Scholar] [CrossRef]
- Li, S.F.; Mathias, J.A. The critical period of high mortality of larvae fish-a discussion based on current research. Chin. J. Oceanol. Limn. 1987, 5, 80–96. [Google Scholar] [CrossRef]
- Leggett, W.C.; Deblois, E. Recruitment in marine fishes: Is it regulated by starvation and predation in the egg and larval stages? Neth. J. Fish. Res. 1994, 32, 119–134. [Google Scholar] [CrossRef]
- Sirois, P.; Dodson, J.J. Critical periods and growth-dependent survival of larvae of an estuarine fish, the rainbow smelt Osmerus mordax. Mar. Ecol. Prog. Ser. 2000, 203, 233–245. [Google Scholar] [CrossRef]
- Weber, R.A.; Peleteiro, J.B.; García Martín, L.O.; Aldegunde, M. The efficacy of 2-phenoxyethanol, metomidate, clove oil and MS-222 as anaesthetic agents in the Senegalese sole (Solea senegalensis Kaup 1858). Aquaculture 2009, 288, 147–150. [Google Scholar] [CrossRef]
- Srithongthum, S.; Au, H.L.; Amornsakun, T.; Chesoh, S.; Jantarat, S.; Suzuki, N.; Takeuchi, Y.; Hassan, A.; Kawamura, G.; Lim, L. Yolk-sac absorption, mouth size development, and first exogenous feeding of sultan fish, Leptobarbus hoevenii. AACL Bioflux 2020, 13, 1320–1327. Available online: http://www.bioflux.com.ro/aacl (accessed on 7 May 2023).
- Soler, P.; Faria, M.; Barata, C.; García-Galea, E.; Lorente, B.; Vinyoles, D. Improving water quality does not guarantee fish health: Effects of ammonia pollution on the behaviour of wild-caught pre-exposed fish. PLoS ONE 2021, 16, e0243404. [Google Scholar] [CrossRef] [PubMed]
- Montoya, A.; López-Olmeda, J.F.; Yúfera, M.; Sánchez-Muros, M.J.; Sánchez-Vázquez, F.J. Feeding time synchronises daily rhythms of behaviour and digestive physiology in gilthead seabream (Sparus aurata). Aquaculture 2010, 306, 315–321. [Google Scholar] [CrossRef]
- Fast, A.W.; Qin, T.; Szyper, J.P. A new method for assessing fish feeding rhythms using demand feeders and automated data acquisition. Aquacult. Eng. 1997, 16, 213–220. [Google Scholar] [CrossRef]
- Petit, G.; Beauchaud, M.; Attia, J.; Buisson, B. Food intake and growth of largemouth bass (Micropterus salmoides) held under alternated light/dark cycle (12L:12D) or exposed to continuous light. Aquaculture 2003, 228, 397–401. [Google Scholar] [CrossRef]
- Fukuhara, O. Morphological and functional development of larval and juvenile Limanda yokohamae (Pisces: Pleuronectidae) reared in the laboratory. Mar. Biol. 1988, 99, 271–281. [Google Scholar] [CrossRef]
- Khemis, I.B.; Gisbert, E.; Alcaraz, C.; Zouiten, D.; Besbes, R.; Zouiten, A.; Masmoudi, A.S.; Cahu, C. Allometric growth patterns and development in larvae and juveniles of thick-lipped grey mullet Chelon labrosus reared in mesocosm conditions. Aquac. Res. 2013, 44, 1872–1888. [Google Scholar] [CrossRef]
- Zhang, Z.; Jiang, L.L.; Wang, Z.; Du, R.B.; Wu, Y.Q.; Liu, L.M. Growth, development, and feeding characteristics of Thamnaconus modestus during early stages. Mar. Sci. 2021, 45, 1–13, (In Chinese with English Abstract). Available online: http://qdhys.ijournal.cn/html/hykx/2021/1/20210101.html (accessed on 10 May 2023).
- Balon, E.K. Types of feeding in the ontogeny of fishes and the life-history model. In Contemporary Studies on Fish Feeding: The Proceedings of GUTSHOP’84: Papers from the Fourth Workshop on Fish Food Habits Held at the Asilomar Conference Center, Pacific Grove: California, USA, 2–6 December 1984; Springer: Dordrecht, The Netherlands, 1986; pp. 11–24. [Google Scholar] [CrossRef]
- Moteki, M.; Yoseda, K.; Sahin, T.; Üstündağ, C.; Kohno, H. Transition from endogenous to exogenous nutritional sources in larval Black Sea turbot Psetta maxima. Fish. Sci. 2001, 67, 571–578. [Google Scholar] [CrossRef]
- Hansen, T.J.; Møller, D. Yolk absorption, yolk sac constrictions, mortality, and growth during first feeding of Atlantic salmon (Salmo salar) incubated on astro-turf. Can. J. Fish. Aquat. Sci. 1985, 42, 1073–1078. [Google Scholar] [CrossRef]
- Fukuhara, O. Development and growth of laboratory reared Engraulis japonica (Houttuyn) larvae. J. Fish Biol. 1983, 23, 641–652. [Google Scholar] [CrossRef]
- Williams, K.; Papanikos, N.; Phelps, R.P.; Shardo, J.D. Development, growth, and yolk utilization of hatchery-reared red snapper Lutjanus campechanus larvae. Mar. Ecol. Prog. Ser. 2004, 275, 231–239. [Google Scholar] [CrossRef]
- Dhaneesh, K.V.; Kumar, T.A.; Divya, S.P.; Kumaresan, S.; Balasubramanian, T. Influence of prompt first feeding on growth and survival of Clownfish Amphiprion percula larvae. Emir. J. Food Agric. 2012, 24, 92–97. [Google Scholar] [CrossRef]
- Zamal, H.; Ollevier, F. Effect of feeding and lack of food on the growth, gross biochemical and fatty acid composition of juvenile catfish. J. Fish Biol. 1995, 46, 404–414. [Google Scholar] [CrossRef]
- Diaz, M.V.; Do Souto, M.; Betti, P.; Temperoni, B.; Schiariti, A.; Machinandiarena, L.; Brown, D.R.; Macchi, G.J. Evaluating the role of endogenous and exogenous features on larval hake nutritional condition. Fish. Oceanogr. 2020, 29, 584–596. [Google Scholar] [CrossRef]
- Krylov, V.V.; Izvekov, E.I.; Pavlova, V.V.; Pankova, N.A.; Osipova, E.A. Circadian rhythms in zebrafish (Danio rerio) behaviour and the sources of their variability. Biol. Rev. 2021, 96, 785–797. [Google Scholar] [CrossRef]
- Assan, D.; Huang, Y.; Mustapha, U.F.; Addah, M.N.; Li, G.; Chen, H. Fish feed intake, feeding behavior, and the physiological response of apelin to fasting and refeeding. Front. Endocrinol. 2021, 12, 798903. [Google Scholar] [CrossRef] [PubMed]
- Kulczykowska, E.; Sánchez Vázquez, F.J. Neurohormonal regulation of feed intake and response to nutrients in fish: Aspects of feeding rhythm and stress. Aquac. Res. 2010, 41, 654–667. [Google Scholar] [CrossRef]
- Heilman, M.J.; Spieler, R.E. The daily feeding rhythm to demand feeders and the effects of timed meal-feeding on the growth of juvenile Florida pompano, Trachinotus carolinus. Aquaculture 1999, 180, 53–64. [Google Scholar] [CrossRef]
- del Pozo, A.; Sánchez-Férez, J.A.; Sánchez-Vázquez, F.J. Circadian rhythms of self-feeding and locomotor activity in zebrafish (Danio rerio). Chronobiol. Int. 2011, 28, 39–47. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| dph | Sampling Numbers | Feeding Rate (%) | Satiety Prey Item | Satiety Rate (%) |
|---|---|---|---|---|
| 3 | 30 | 73.3 | 5 (S-Rotifer) | 33.33 |
| 4 | 30 | 90 | 7 (S-Rotifer) | 36.67 |
| 5 | 30 | 96.7 | 13 (S-Rotifer) | 43.33 |
| 8 | 30 | 100 | 24 (L-Rotifer) | 76.67 |
| 12 | 30 | 100 | 32 (L-Rotifer) | 83.33 |
| 25 | 30 | 100 | 37 (Artemia salina nauplii) | 83.33 |
| 35 | 30 | 100 | 143 (A. salina nauplii) | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Zeng, J.; Mei, W.; Jiang, L.; Manabe, S.; Wu, Y.; Liu, L. Changes in Growth and Feeding Characteristics during Early Ontogenesis in Threadsail Filefish, Stephanolepis cirrhifer. Animals 2023, 13, 3420. https://doi.org/10.3390/ani13213420
Xu W, Zeng J, Mei W, Jiang L, Manabe S, Wu Y, Liu L. Changes in Growth and Feeding Characteristics during Early Ontogenesis in Threadsail Filefish, Stephanolepis cirrhifer. Animals. 2023; 13(21):3420. https://doi.org/10.3390/ani13213420
Chicago/Turabian StyleXu, Wengang, Jun Zeng, Weiping Mei, Lianglong Jiang, Soichiro Manabe, Yanqin Wu, and Liming Liu. 2023. "Changes in Growth and Feeding Characteristics during Early Ontogenesis in Threadsail Filefish, Stephanolepis cirrhifer" Animals 13, no. 21: 3420. https://doi.org/10.3390/ani13213420