
Isolation and Characterization of Cetacean Cell-Derived Extracellular Vesicles
 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell lines
2.2. Isolation of Extracellular Vesicles with Ultracentrifugation (UC) and Size Exclusion Chromatography (SEC)
2.3. Nanoparticle Tracking Analysis (NTA)
2.4. Protein Extraction and Western Blotting (WB) Analysis
2.5. Scanning Transmission Electron Microscopy
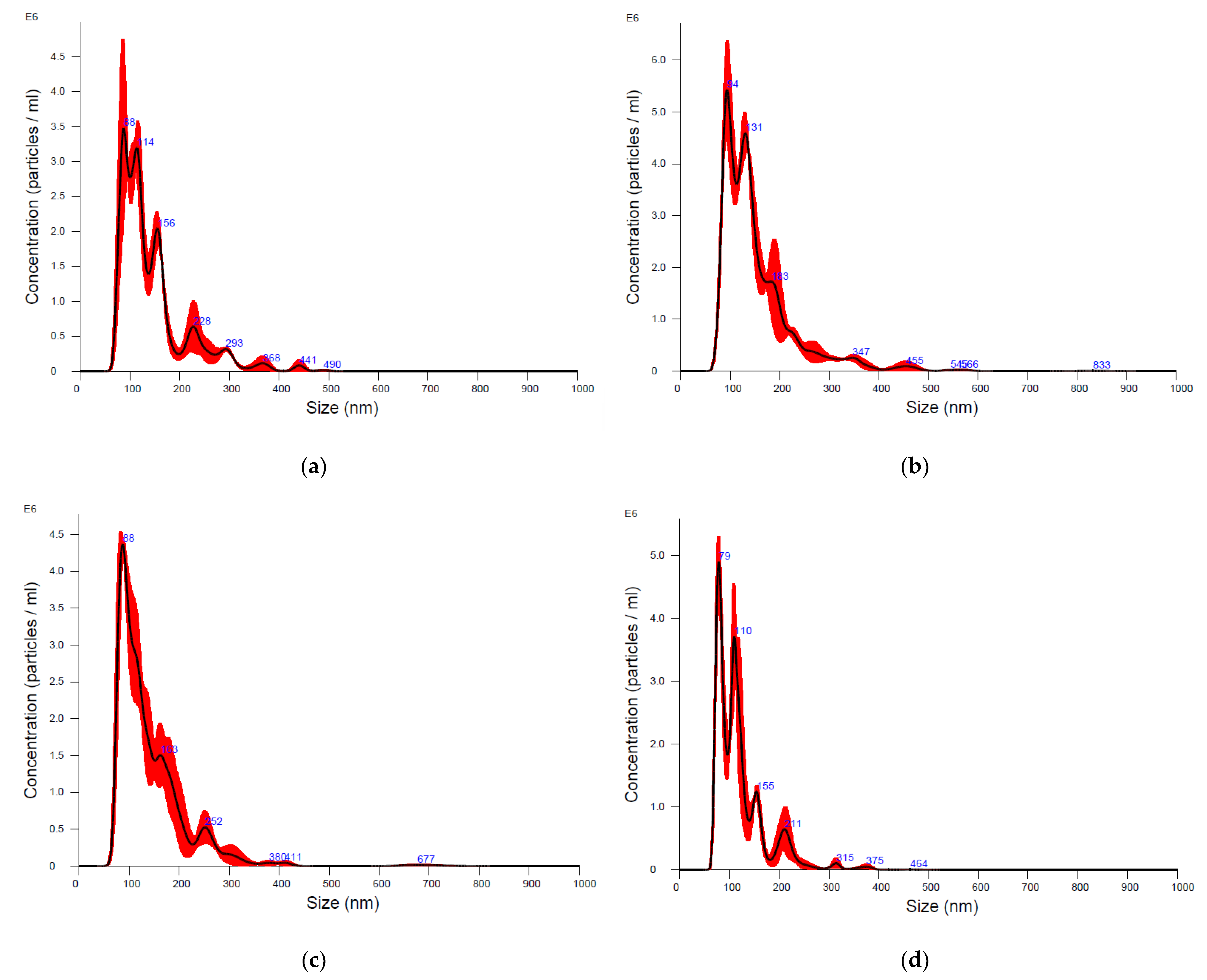
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; Angelo, G.D.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Crescitelli, R.; Lässer, C.; Lötvall, J. Isolation and Characterization of Extracellular Vesicle Subpopulations from Tissues. Nat. Protoc. 2021, 16, 1548–1580. [Google Scholar] [CrossRef]
- Monguió-Tortajada, M.; Gálvez-Montón, C.; Bayes-Genis, A.; Roura, S.; Borràs, F.E. Extracellular Vesicle Isolation Methods: Rising Impact of Size-Exclusion Chromatography. Cell. Mol. Life Sci. 2019, 76, 2369–2382. [Google Scholar] [CrossRef]
- Coumans, F.A.W.; Brisson, A.R.; Buzas, E.I.; Dignat-george, F.; Drees, E.E.E.; El-andaloussi, S.; Emanueli, C.; Gasecka, A.; Hendrix, A.; Hill, A.F.; et al. Review Extracellular Vesicles Methodological Guidelines to Study Extracellular Vesicles. Circ. Res. 2017, 120, 1632–1648. [Google Scholar] [CrossRef]
- Xu, R.; Greening, D.W.; Zhu, H.-J.; Takahashi, N.; Simpson, R.J. Extracellular Vesicle Isolation and Characterization: Toward Clinical Application. J. Clin. Investig. 2016, 126, 1152–1162. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Hill, A.F. Therapeutically Harnessing Extracellular Vesicles. Nat. Rev. Drug Discov. 2022, 21, 379–399. [Google Scholar] [CrossRef]
- Moccia, V.; Sammarco, A.; Cavicchioli, L.; Castagnaro, M.; Bongiovanni, L.; Zappulli, V. Extracellular Vesicles in Veterinary Medicine. Animals 2022, 12, 2716. [Google Scholar] [CrossRef] [PubMed]
- Urzì, O.; Raimondo, S.; Alessandro, R. Extracellular Vesicles from Plants: Current Knowledge and Open Questions. Int. J. Mol. Sci. 2021, 22, 5366. [Google Scholar] [CrossRef] [PubMed]
- Sartorio, M.G.; Pardue, E.J.; Feldman, M.F.; Haurat, M.F. Bacterial Outer Membrane Vesicles: From Discovery to Applications. Annu. Rev. Microbiol. 2021, 75, 609–630. [Google Scholar] [CrossRef]
- Magnadóttir, B.; Uysal-Onganer, P.; Kraev, I.; Svansson, V.; Skírnisson, K.; Lange, S. Deiminated Proteins and Extracellular Vesicles as Novel Biomarkers in Pinnipeds: Grey Seal (Halichoerus gryptus) and Harbour Seal (Phoca vitulina). Biochimie 2020, 171–172, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Magnadóttir, B.; Uysal-Onganer, P.; Kraev, I.; Svansson, V.; Hayes, P.; Lange, S. Deiminated Proteins and Extracellular Vesicles-Novel Serum Biomarkers in Whales and Orca. Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 34, 100676. [Google Scholar] [CrossRef]
- Bossart, G.D. Marine Mammals as Sentinel Species for Oceans and Human Health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A. Deciphering the Whale’s Secrets to Have a Long Life. Exp. Gerontol. 2021, 151, 111425. [Google Scholar] [CrossRef]
- Finlayson, K.A.; Van de Merwe, J.P. Differences in Marine Megafauna in Vitro Sensitivity Highlights the Need for Species-Specific Chemical Risk Assessments. Aquat. Toxicol. 2021, 239, 105939. [Google Scholar] [CrossRef] [PubMed]
- Otero-Sabio, C.; Giacomello, M.; Centelleghe, C.; Caicci, F.; Bonato, M.; Venerando, A.; Graïc, J.M.; Mazzariol, S.; Finos, L.; Corain, L.; et al. Cell Cycle Alterations Due to Perfluoroalkyl Substances PFOS, PFOA, PFBS, PFBA and the New PFAS C6O4 on Bottlenose Dolphin (Tursiops truncatus) Skin Cell. Ecotoxicol. Environ. Saf. 2022, 244, 113980. [Google Scholar] [CrossRef]
- Jia, K.; Ding, L.; Zhang, L.; Zhang, M.; Yi, M.; Wu, Y. In Vitro Assessment of Environmental Stress of Persistent Organic Pollutants on the Indo-Pacific Humpback Dolphin. Toxicol. Vitr. 2015, 30, 529–535. [Google Scholar] [CrossRef]
- Goodwin, T.J.; Coate-li, L.; Linnehan, R.M.; Hammond, T.G. Cellular Responses to Mechanical Stress Selected Contribution: A Three-Dimensional Model for Assessment of in Vitro Toxicity in Balaena Mysticetus Renal Tissue. J. Appl. Physiol. 2000, 89, 2508–2517. [Google Scholar] [CrossRef]
- Fossi, M.C.; Casini, S.; Bucalossi, D.; Marsili, L. First Detection of CYP1A1 and CYP2B Induction in Mediterranean Cetacean Skin Biopsies and Cultured Fibroblasts by Western Blot Analysis. Mar. Environ. Res. 2008, 66, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, M.; Kurihara, N.; Hirano, M.; Nakata, A.; Iwata, H. In Vitro Cytotoxicity and Risk Assessments of Environmental Pollutants Using Fibroblasts of a Stranded Finless Porpoise (Neophocaena asiaeorientalis). Environ. Sci. Technol. 2020, 54, 6832–6841. [Google Scholar] [CrossRef]
- Huang, Y.; Rajput, I.R.; Sanganyado, E.; Yajing, S.; Yu, F.; Liang, B.; Liu, W. Immune Stimulation Effect of PBDEs via Prostaglandin Pathway in Pantropical Spotted Dolphin: An in Vitro Study. Chemosphere 2020, 254, 126717. [Google Scholar] [CrossRef]
- Gauthier, J.M.; Dubeau, H.; Rassart, E. Induction of Micronuclei in Vitro by Organochlorine Compounds in Beluga Whale Skin Fibroblasts. Mutat. Res.-Genet. Toxicol. Environ. Mutagen. 1999, 439, 87–95. [Google Scholar] [CrossRef]
- Moccia, V.; Sammarco, A.; Ferro, S.; Cavicchioli, L.; Zappulli, V. Characterization and Function of Extracellular Vesicles in a Canine Mammary Tumour Cell Line: Ultracentrifugation versus Size Exclusion Chromatography. Vet. Comp. Oncol. 2023, 21, 36–44. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Gámez-Valero, A.; Monguió-Tortajada, M.; Carreras-Planella, L.; Franquesa, M.; Beyer, K.; Borràs, F.E. Size-Exclusion Chromatography-Based Isolation Minimally Alters Extracellular Vesicles’ Characteristics Compared to Precipitating Agents. Sci. Rep. 2016, 6, 33641. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Li, Y.J.; Wu, J.Y.; Cai, J.X.; Wen, J.; Xiang, D.X.; Hu, X.B.; Li, W.Q. Comparative Evaluation of Methods for Isolating Small Extracellular Vesicles Derived from Pancreatic Cancer Cells. Cell Biosci. 2021, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Hu, X.M.; Su, Z.H.; Zeng, X.B.; Ma, H.Y.; Xiong, K. Tissue-Derived Extracellular Vesicles: Research Progress from Isolation to Application. Pathol. Res. Pract. 2021, 226, 153604. [Google Scholar] [CrossRef]
- Barreiro, K.; Lay, A.C.; Leparc, G.; Tran, V.D.T.; Rosler, M.; Dayalan, L.; Burdet, F.; Ibberson, M.; Coward, R.J.M.; Huber, T.B.; et al. An in Vitro Approach to Understand Contribution of Kidney Cells to Human Urinary Extracellular Vesicles. J. Extracell. Vesicles 2023, 12, e12304. [Google Scholar] [CrossRef]
- Bilandžić, N.; Sedak, M.; Dokić, M.; Gomerčić, M.D.; Gomerčić, T.; Zadravec, M.; Benić, M.; Crnić, A.P. Toxic Element Concentrations in the Bottlenose (Tursiops truncatus), Striped (Stenella coeruleoalba) and Risso’s (Grampus griseus) Dolphins Stranded in Eastern Adriatic Sea. Bull. Environ. Contam. Toxicol. 2012, 89, 467–473. [Google Scholar] [CrossRef]
- Reif, J.S.; Schaefer, A.M.; Bossart, G.D. Atlantic Bottlenose Dolphins (Tursiops truncatus) as a Sentinel for Exposure to Mercury in Humans: Closing the Loop. Vet. Sci. 2015, 2, 407–422. [Google Scholar] [CrossRef]
- Ruiz-Hernández, I.M.; Nouri, M.Z.; Kozuch, M.; Denslow, N.D.; Díaz-Gamboa, R.E.; Rodríguez-Canul, R.; Collí-Dulá, R.C. Trace Element and Lipidomic Analysis of Bottlenose Dolphin Blubber from the Yucatan Coast: Lipid Composition Relationships. Chemosphere 2022, 299, 134353. [Google Scholar] [CrossRef]
- Grattarola, C.; Giorda, F.; Iulini, B.; Pintore, M.D.; Pautasso, A.; Zoppi, S.; Goria, M.; Romano, A.; Peletto, S.; Varello, K.; et al. Meningoencephalitis and Listeria Monocytogenes, Toxoplasma Gondii and Brucella spp. Coinfection in a Dolphin in Italy. Dis. Aquat. Organ. 2016, 118, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Grattarola, C.; Gallina, S.; Giorda, F.; Pautasso, A.; Ballardini, M.; Iulini, B.; Varello, K.; Goria, M.; Peletto, S.; Masoero, L.; et al. First Report of Salmonella 1,4,[5],12:I:- In Free-Ranging Striped Dolphins (Stenella coeruleoalba), Italy. Sci. Rep. 2019, 9, 6061. [Google Scholar] [CrossRef]
- Sévellec, Y.; Torresi, M.; Félix, B.; Palma, F.; Centorotola, G.; Bilei, S.; Senese, M.; Terracciano, G.; Leblanc, J.C.; Pomilio, F.; et al. First Report on the Finding of Listeria Monocytogenes St121 Strain in a Dolphin Brain. Pathogens 2020, 9, 802. [Google Scholar] [CrossRef]
- Marangi, M.; Carlucci, R.; Carlino, P.; Fanizza, C.; Cirelli, G.; Maglietta, R.; Beneduce, L. Dolphins and Sea Turtles May Host Zoonotic Parasites and Pathogenic Bacteria as Indicators of Anthropic Pressure in the Gulf of Taranto (Northern Ionian Sea, Central-Eastern Mediterranean Sea). Vet. Res. Commun. 2022, 46, 1157–1166. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; Terregino, C.; Aznar, I.; Guajardo, I.M.; et al. Avian Influenza Overview September–December 2022. EFSA J. 2023, 21, e07786. [Google Scholar] [CrossRef]
- Di Guardo, G.; Mazzariol, S. Cetacean Morbillivirus: A Land-to-Sea Journey and Back? Virol. Sin. 2019, 34, 240–242. [Google Scholar] [CrossRef]
- Tian, R.; Wang, Z.; Niu, X.; Zhou, K.; Xu, S.; Yang, G. Evolutionary Genetics of Hypoxia Tolerance in Cetaceans during Diving. Genome Biol. Evol. 2016, 8, 827–839. [Google Scholar] [CrossRef]
- Noren, S.R. Postnatal Development of Diving Physiology: Implications of Anthropogenic Disturbance for Immature Marine Mammals. J. Exp. Biol. 2020, 223, jeb227736. [Google Scholar] [CrossRef]
- Hindle, A.G. Diving Deep: Understanding the Genetic Components of Hypoxia Tolerance in Marine Mammals. J. Appl. Physiol. 2020, 128, 1439–1446. [Google Scholar] [CrossRef]
- Schorr, G.S.; Falcone, E.A.; Moretti, D.J.; Andrews, R.D. First Long-Term Behavioral Records from Cuvier’s Beaked Whales (Ziphius Cavirostris) Reveal Record-Breaking Dives. PLoS ONE 2014, 9, e92633. [Google Scholar] [CrossRef]
- Bernaldo De Quirós, Y.; Fernandez, A.; Baird, R.W.; Brownell, R.L.; Aguilar De Soto, N.; Allen, D.; Arbelo, M.; Arregui, M.; Costidis, A.; Fahlman, A.; et al. Advances in Research on the Impacts of Anti-Submarine Sonar on Beaked Whales. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182533. [Google Scholar] [CrossRef] [PubMed]
- Beineke, A.; Siebert, U.; Wohlsein, P.; Baumgärtner, W. Immunology of Whales and Dolphins. Vet. Immunol. Immunopathol. 2010, 133, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Seluanov, A.; Gladyshev, V.N.; Vijg, J.; Gorbunova, V. Mechanisms of Cancer Resistance in Long-Lived Mammals. Nat. Rev. Cancer 2018, 18, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Chai, S.; Huang, X.; Wang, Y.; Xiao, L.; Xu, S.; Yang, G. Novel Genomic Insights into Body Size Evolution in Cetaceans and a Resolution of Peto’s Paradox. Am. Nat. 2021, 199, E28–E42. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample | Particle Concentration/mL +/− SD * | Particle Mean Size (nm) +/− SD |
|---|---|---|
| EV-UC bottlenose dolphin | 2.9 × 1011 +/− 2.7 × 1010 | 140.8 +/− 3.1 |
| EV-UC Cuvier’s beaked whale | 4.7 × 1011 +/− 2.8 × 109 | 151.9 +/− 7.3 |
| EV-SEC bottlenose dolphin | 2.7 × 109 +/− 1.9 × 108 | 144.1 +/− 9.9 |
| EV-SEC Cuvier’s beaked whale | 2 × 109 +/− 1.4 × 108 | 120.7 +/− 2.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moccia, V.; Centelleghe, C.; Giusti, I.; Peruffo, A.; Dolo, V.; Mazzariol, S.; Zappulli, V. Isolation and Characterization of Cetacean Cell-Derived Extracellular Vesicles. Animals 2023, 13, 3304. https://doi.org/10.3390/ani13213304
Moccia V, Centelleghe C, Giusti I, Peruffo A, Dolo V, Mazzariol S, Zappulli V. Isolation and Characterization of Cetacean Cell-Derived Extracellular Vesicles. Animals. 2023; 13(21):3304. https://doi.org/10.3390/ani13213304
Chicago/Turabian StyleMoccia, Valentina, Cinzia Centelleghe, Ilaria Giusti, Antonella Peruffo, Vincenza Dolo, Sandro Mazzariol, and Valentina Zappulli. 2023. "Isolation and Characterization of Cetacean Cell-Derived Extracellular Vesicles" Animals 13, no. 21: 3304. https://doi.org/10.3390/ani13213304