

Effects of Dietary Betaine on the Laying Performance, Antioxidant Capacity, and Uterus and Ovary Function of Laying Hens at the Late Stage of Production
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds and Husbandry
2.2. Measurement of Antioxidant Enzyme Activity
2.3. Quantitation of mRNA Expression via Real-Time PCR
2.4. Methylated DNA Immunoprecipitation (MeDIP) Analysis
2.5. Statistical Analysis
3. Results
3.1. Betaine Increased the Egg-Laying Rate
3.2. Betaine Increased the Eggshell Thickness
3.3. The Effect of Betaine on the Antioxidant Content in the Uterus and Ovary
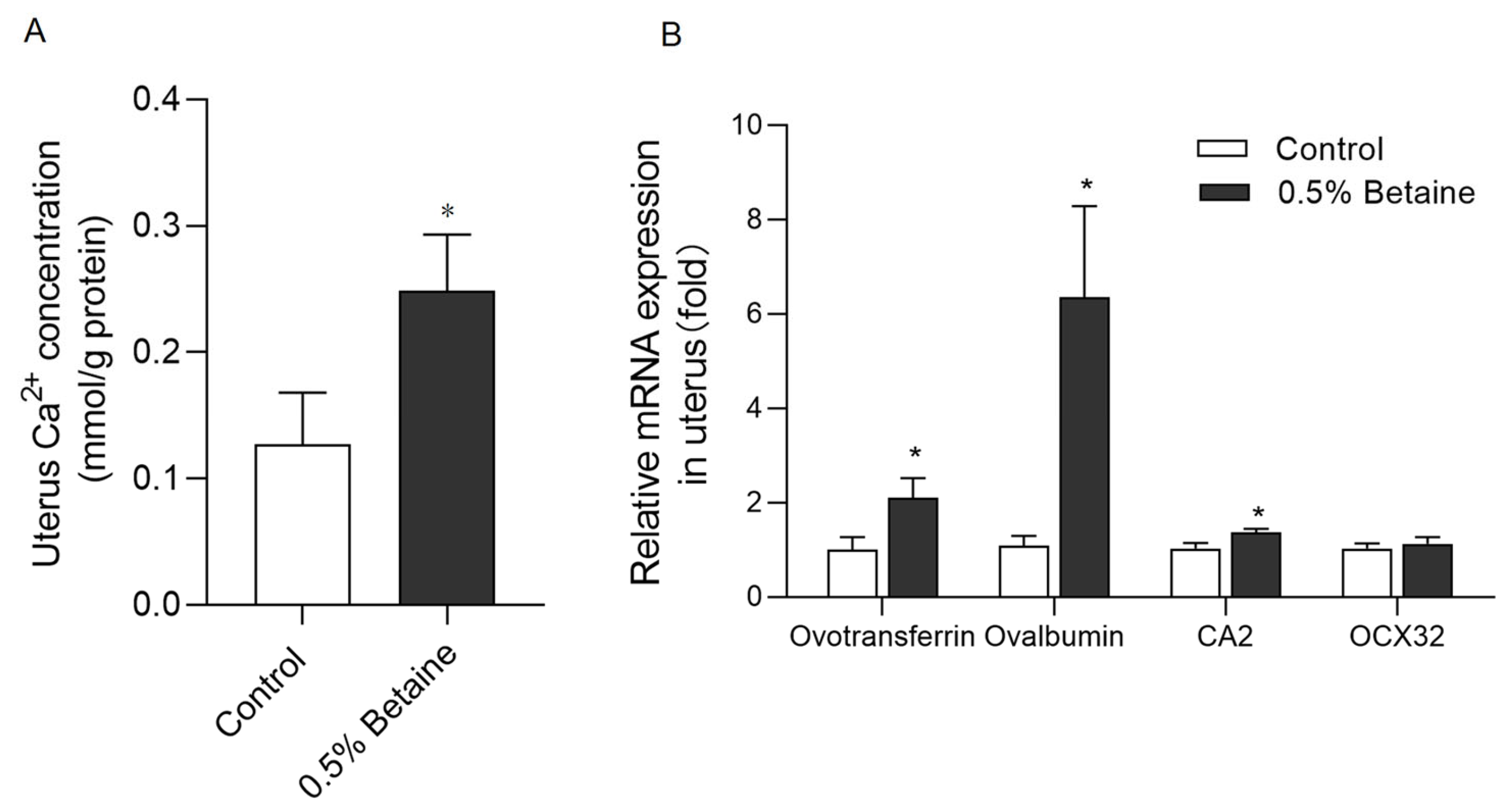
3.4. Dietary Betaine Consumption Increased the Ca2+ Concentration and Calcification-Associated Gene Expressions in the Uterus
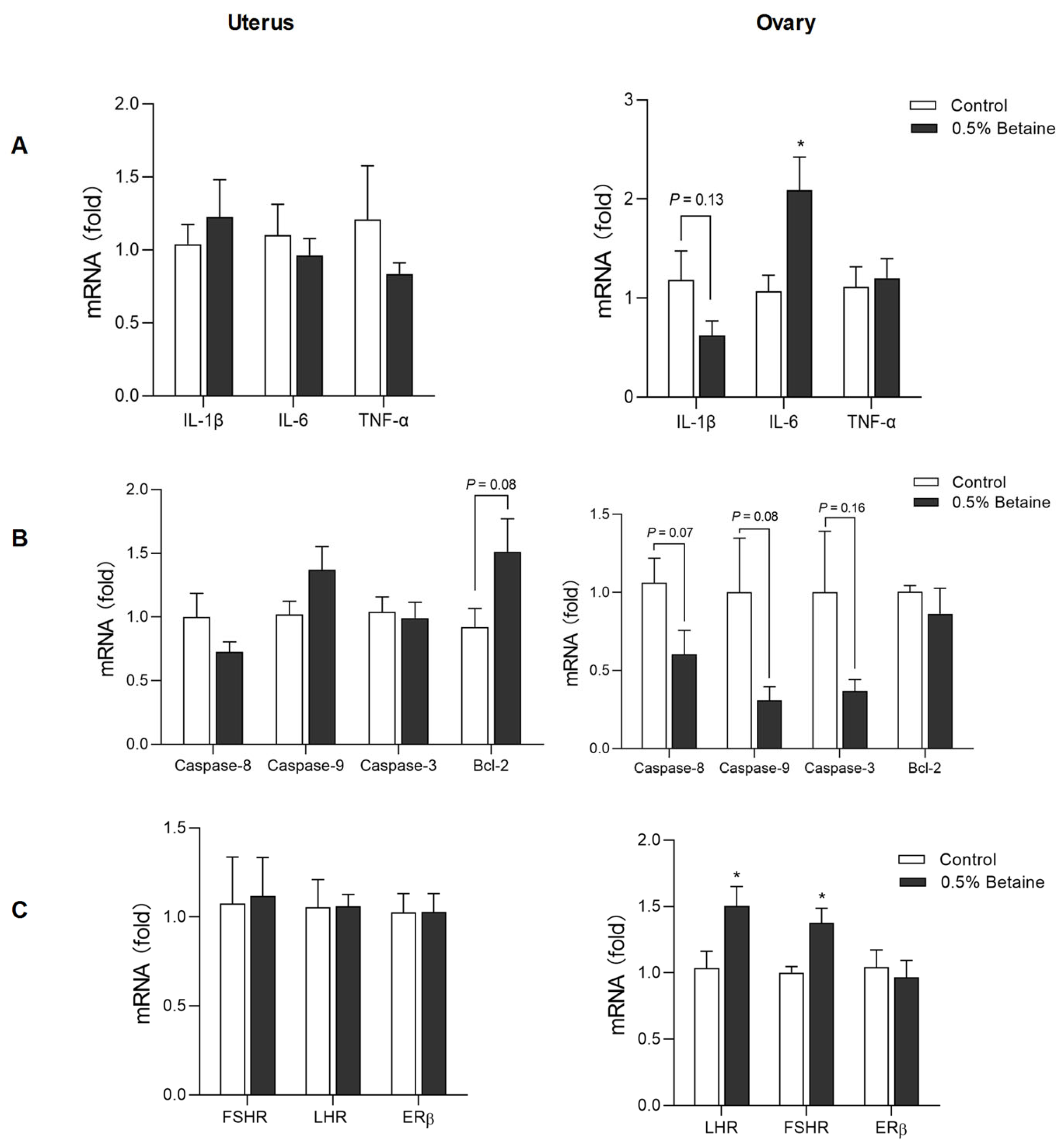
3.5. Effect of Dietary Betaine on the mRNA Expression of Pro-Inflammatory Cytokines, Apoptosis Factors, and Reproductive Hormone Receptors
3.6. Effect of Dietary Betaine on the mRNA Expression of DNA Methylation-Associated Enzymes
3.7. Effects of Dietary Betaine on DNA Methylation of FSHR, LHR, Caspase-3, and Caspase-8 Gene Promoters in the Ovary
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, F.; Zou, P.; Xu, S.; Wang, Q.; Zhou, Y.; Li, X.; Tang, L.; Wang, B.; Jin, Q.; Yu, D.; et al. Dietary supplementation of Macleaya cordata extract and Bacillus in combination improve laying performance by regulating reproductive hormones, intestinal microbiota and barrier function of laying hens. J. Anim. Sci. Biotechnol. 2022, 13, 118. [Google Scholar] [CrossRef]
- Matsuda-Minehata, F.; Inoue, N.; Goto, Y.; Manabe, N. The regulation of ovarian granulosa cell death by pro- and anti-apoptotic molecules. J. Reprod. Dev. 2006, 52, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Tardieu, A.; Chazelas, P.; Faye, P.A.; Favreau, F.; Nadal-Desbarats, L.; Sallée, C.; Margueritte, F.; Couquet, C.Y.; Marquet, P.; Guellec, C.B.; et al. Changes in the metabolic composition of storage solution with prolonged cold ischemia of the uterus. J. Assist. Reprod. Genet. 2019, 36, 1169–1178. [Google Scholar] [CrossRef]
- Yang, J.X.; Chaudhry, M.T.; Yao, J.Y.; Wang, S.N.; Zhou, B.; Wang, M.; Han, C.Y.; You, Y.; Li, Y. Effects of phyto-oestrogen quercetin on productive performance, hormones, reproductive organs and apoptotic genes in laying hens. J. Anim. Physiol. Anim. Nutr. 2018, 102, 505–513. [Google Scholar] [CrossRef]
- Wu, Y.; Li, M.; Zhang, J.; Wang, S. Unveiling uterine aging: Much more to learn. Ageing Res. Rev. 2023, 86, 101879. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Feng, Y.; Ding, Z.; Lv, L.; Sui, Y.; Sun, Q.; Abobaker, H.; Cai, D.; Zhao, R. Maternal betaine supplementation decreases hepatic cholesterol deposition in chicken offspring with epigenetic modulation of SREBP2 and CYP7A1 genes. Poult. Sci. 2020, 99, 3111–3120. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Chen, Y.; Leng, Z.; Ding, L.; Wang, T.; Zhou, Y. Dietary betaine improves meat quality and oxidative status of broilers under heat stress. J. Sci. Food Agric. 2019, 99, 620–623. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Y.; Ou, C.; Ma, J.; Wang, Q.; Du, S.; Xu, Z.; Li, R.; Guo, F. Alleviation of infectious-bursal-disease-virus-induced bursal injury by betaine is associated with DNA methylation in IL-6 and interferon regulatory factor 7 promoter. Poult. Sci. 2019, 98, 4457–4464. [Google Scholar] [CrossRef]
- Attia, Y.A.; Abd El-Hamid Ael, H.; Abedalla, A.A.; Berika, M.A.; Al-Harthi, M.A.; Kucuk, O.; Sahin, K.; Abou-Shehema, B.M. Laying performance, digestibility and plasma hormones in laying hens exposed to chronic heat stress as affected by betaine, vitamin C, and/or vitamin E supplementation. SpringerPlus 2016, 5, 1619. [Google Scholar] [CrossRef]
- Omer, N.A.; Hu, Y.; Idriss, A.A.; Abobaker, H.; Hou, Z.; Yang, S.; Ma, W.; Zhao, R. Dietary betaine improves egg-laying rate in hens through hypomethylation and glucocorticoid receptor-mediated activation of hepatic lipogenesis-related genes. Poult. Sci. 2020, 99, 3121–3132. [Google Scholar] [CrossRef]
- Bekdash, R.A. Methyl Donors, Epigenetic Alterations, and Brain Health: Understanding the Connection. Int. J. Mol. Sci. 2023, 24, 2346. [Google Scholar] [CrossRef]
- Omer, N.A.; Hu, Y.; Idriss, A.A.; Abobaker, H.; Hou, Z.; Dong, H.; Zhao, R. Dietary betaine activates hepatic VTGII expression in laying hens associated with hypomethylation of GR gene promoter and enhanced GR expression. J. Anim. Sci. Biotechnol. 2018, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Jing, M.; Zhang, A.; Yu, Y.; Gao, P.; Wang, Q.; Wang, L.; Xu, Z.; Ma, J.; Zhang, Y. Betaine Alleviates LPS-Induced Chicken Skeletal Muscle Inflammation with the Epigenetic Modulation of the TLR4 Gene. Animals 2022, 12, 1899. [Google Scholar] [CrossRef]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.B.; Gao, Y.; et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef]
- Cheng, X.; Liu, Y.; Zhang, J.; Ji, C.; Zhao, L.; Ma, Q. DNA methylome and transcriptome identified Key genes and pathways involved in Speckled Eggshell formation in aged laying hens. BMC Genom. 2023, 24, 31. [Google Scholar] [CrossRef]
- Zhang, T.; Li, C.; Deng, J.; Jia, Y.; Qu, L.; Ning, Z. Chicken Hypothalamic and Ovarian DNA Methylome Alteration in Response to Forced Molting. Animals 2023, 13, 1012. [Google Scholar] [CrossRef] [PubMed]
- Gudev, D.; Popova-Ralcheva, S.; Yanchev, I.; Moneva, P.; Petkov, E.; Ignatova, M. Effect of betaine on egg performance and some blood constituents in laying hens reared indoors under natural summer temperatures and varying levels of air ammonia. Bulg. J. Agric. Sci. 2011, 17, 8. [Google Scholar]
- Liu, S.; Zhao, L.; Ma, Q. Effects of Dietary Vitamin E Levels on the Productive Performance and Egg Quality During PeakProduction Period of Jing Hong Laying Hens. Chin. J. Anim. Sci. 2018, 54, 4. [Google Scholar] [CrossRef]
- Zhang, H.D.; Zhao, X.F.; Ren, Z.Z.; Tong, M.Q.; Chen, J.N.; Li, S.Y.; Chen, H.; Wang, D.H. Comparison between different breeds of laying hens in terms of eggshell translucency and its distribution in various ends of the eggshell. Poult. Sci. 2021, 100, 101510. [Google Scholar] [CrossRef]
- Ratriyanto, A.; Prastowo, S. Floor space and betaine supplementation alter the nutrient digestibility and performance of Japanese quail in a tropical environment. J. Therm. Biol. 2019, 83, 80–86. [Google Scholar] [CrossRef]
- Henny, C.J.; Bennett, J.K. Comparison of breaking strength and shell thickness as evaluators of white-faced ibis eggshell quality. Environ. Toxicol. Chem. 1990, 9, 797–805. [Google Scholar] [CrossRef]
- Riczu, C.M.; Saunders-Blades, J.L.; Yngvesson, A.K.; Robinson, F.E.; Korver, D.R. End-of-cycle bone quality in white- and brown-egg laying hens. Poult. Sci. 2004, 83, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Gautron, J.; Stapane, L.; Le Roy, N.; Nys, Y.; Rodriguez-Navarro, A.B.; Hincke, M.T. Avian eggshell biomineralization: An update on its structure, mineralogy and protein tool kit. BMC Mol. Cell Biol. 2021, 22, 11. [Google Scholar] [CrossRef]
- Jonchère, V.; Réhault-Godbert, S.; Hennequet-Antier, C.; Cabau, C.; Sibut, V.; Cogburn, L.A.; Nys, Y.; Gautron, J. Gene expression profiling to identify eggshell proteins involved in physical defense of the chicken egg. BMC Genom. 2010, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Zhao, Y.; Mahmood, T.; Guo, Y. Effects of dietary vitamins supplementation level on the production performance and intestinal microbiota of aged laying hens. Poult. Sci. 2020, 99, 3594–3605. [Google Scholar] [CrossRef]
- Marie, P.; Labas, V.; Brionne, A.; Harichaux, G.; Hennequet-Antier, C.; Rodriguez-Navarro, A.B.; Nys, Y.; Gautron, J. Quantitative proteomics provides new insights into chicken eggshell matrix protein functions during the primary events of mineralisation and the active calcification phase. J. Proteom. 2015, 126, 140–154. [Google Scholar] [CrossRef]
- Wang, X.; Wu, C.; Tao, K.; Zhao, K.; Wang, J.; Xu, H.; Xia, D.; Shan, H.; Lu, J.R. Influence of ovalbumin on CaCO3 precipitation during in vitro biomineralization. J. Phys. Chem. B 2010, 114, 5301–5308. [Google Scholar] [CrossRef]
- Gay, C.V.; Faleski, E.J.; Schraer, H.; Schraer, R. Localization of carbonic anhydrase in avian gastric mucosa, shell gland and bone by immunohistochemistry. J. Histochem. Cytochem. Off. J. Histochem. Soc. 1974, 22, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Fulton, J.E.; Soller, M.; Lund, A.R.; Arango, J.; Lipkin, E. Variation in the ovocalyxin-32 gene in commercial egg-laying chickens and its relationship with egg production and egg quality traits. Anim. Genet. 2012, 43 (Suppl. 1), 102–113. [Google Scholar] [CrossRef]
- Muhammad, A.I.; Dalia, A.M.; Loh, T.C.; Akit, H.; Samsudin, A.A. Effect of organic and inorganic dietary selenium supplementation on gene expression in oviduct tissues and Selenoproteins gene expression in Lohman Brown-classic laying hens. BMC Vet. Res. 2021, 17, 281. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Li, H.; Miao, L.; Li, L.; Dong, X.; Zou, X. Dietary cadmium chloride impairs shell biomineralization by disrupting the metabolism of the eggshell gland in laying hens. J. Anim. Sci. 2020, 98, 1–14. [Google Scholar] [CrossRef]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Ringuet, M.; Furness, J.B.; Dunshea, F.R. Betaine and Antioxidants Improve Growth Performance, Breast Muscle Development and Ameliorate Thermoregulatory Responses to Cyclic Heat Exposure in Broiler Chickens. Animals 2018, 8, 162. [Google Scholar] [CrossRef]
- Wen, C.; Leng, Z.; Chen, Y.; Ding, L.; Wang, T.; Zhou, Y. Betaine Alleviates Heat Stress-Induced Hepatic and Mitochondrial Oxidative Damage in Broilers. J. Poult. Sci. 2021, 58, 103–109. [Google Scholar] [CrossRef]
- Wang, H.; Cong, X.; Qin, K.; Yan, M.; Xu, X.; Liu, M.; Xu, X.; Zhang, Y.; Gao, Q.; Cheng, S.; et al. Se-Enriched Cardamine violifolia Improves Laying Performance and Regulates Ovarian Antioxidative Function in Aging Laying Hens. Antioxidants 2023, 12, 450. [Google Scholar] [CrossRef]
- He, W.; Wang, H.; Tang, C.; Zhao, Q.; Zhang, J. Dietary supplementation with astaxanthin alleviates ovarian aging in aged laying hens by enhancing antioxidant capacity and increasing reproductive hormones. Poult. Sci. 2023, 102, 102258. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Pors, S.E.; Christensen, J.P.; Bojesen, A.M.; Thøfner, I. Comparison and assessment of necropsy lesions in end-of-lay laying hens from different housing systems in Denmark. Poult. Sci. 2020, 99, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Grafl, B.; Polster, S.; Sulejmanovic, T.; Pürrer, B.; Guggenberger, B.; Hess, M. Assessment of health and welfare of Austrian laying hens at slaughter demonstrates influence of husbandry system and season. Br. Poult. Sci. 2017, 58, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, H.J.; Wu, S.G.; Qi, G.H.; Wang, J. Uterine transcriptome analysis reveals mRNA expression changes associated with the ultrastructure differences of eggshell in young and aged laying hens. BMC Genom. 2020, 21, 770. [Google Scholar] [CrossRef]
- Forcina, L.; Franceschi, C.; Musarò, A. The hormetic and hermetic role of IL-6. Ageing Res. Rev. 2022, 80, 101697. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2013, 38, 23–38. [Google Scholar] [CrossRef]
- Yuan, B.F. 5-methylcytosine and its derivatives. Adv. Clin. Chem. 2014, 67, 151–187. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content (%) |
|---|---|
| Cracked corn | 61.90 |
| Soybean meal | 24.20 |
| Wheat bran | 1.00 |
| Limestone | 8.00 |
| Dicalcium phosphate | 1.50 |
| Methionine | 0.10 |
| Sodium chloride | 0.30 |
| Premix 1 | 3.00 |
| Total | 100 |
| Nutrient level 2 | |
| ME (kcal/kg) | 2584 |
| CP | 15.67 |
| Lys | 0.81 |
| Digestible Lys | 0.72 |
| Thr | 0.61 |
| Digestible Thr | 0.53 |
| Met | 0.26 |
| Digestible Met | 0.24 |
| Calcium | 3.31 |
| Available phosphorus | 0.32 |
| Target Gene | Sequence (F: Forward, R: Reverse, 5′-3′) | GeneBank Access |
|---|---|---|
| CA2 | F: CTCCTCCGACAAGTCAGTGC R: TACGACGGCCAAACCATCAG | NM_205317.1 |
| Ovotransferrin | F: TGACTTCCACCTCTTTGGGC R: GAATCCATCAGCGAGGGGAC | NM_205304.1 |
| Ovalbumin | F: GAGTGGCATCAATGGCTTCTG R: RTCTAGGGCCATACCTGCTCAA | AH002466.2 |
| OCX-32 | F: AAGTCTCCGCCTGTAGTC R: CTTCCTTATCTGCTGCTTCA | NM-204534.5 |
| β-actin | F: TGCGTGACATCAAGGAGAAG R: TGCCAGGGTACATTGTGGTA | NM-205518 |
| TNF-α | F: TCACCCCTACCCTGTCCCA R: AGCCAAGTCAACGCTCCTG | NM-204267.2 |
| IL-1β | F: TTCCGCTACACCCGCTCACA R: TGCCGCTCATCACACACGAC | Y15006 |
| IL-6 | F: GAAATCCCTCCTCGCCAATCTG R: GCCCTCACGGTCTTCTCCATAAA | AJ309540 |
| LHR | F: GGTGTTCTCCATCCTGATAG R: TTGGCAATCTTGGTGTCTT | NM-205079 |
| FSHR | F: ACCTTCCAAGCCTCAGATA R: TTAGCCGTAGAATCACACTT | EF621308 |
| ERβ | F: ACCATTCAACGAAGCACTT R: ACTTCTAACCAGGCACATTC | Ab036415.1 |
| Caspase-9 | F: AGGGAGCAAGCACGACA R: GGTTGGACTGGGATGGAC | XM-424580.6 |
| Caspase-8 | F: AAGGAAGCGGGAAGAT R: GATACCTGAACGGAGACAC | NM-204184.1 |
| Caspase-3 | F: AAGGCTCCTGGTTTATTC R: CTGCCACTCTGCGATTTA | NM-204725.1 |
| Bcl-2 | F: CCGCATCCAGAGGGACT R: CGAAGAAGGCGACGAT | NM-205339.2 |
| DNMT1 | F: TTTTTTTACATAATCCTCCA R: AAAGTATCAATCCCCACTTG | NM_206952.1 |
| DNMT3a | F: ATCACCACTCGCTCCAACTC R: CCAAACACCCTCTCCATCTC | NM_001024832.3 |
| DNMT3b | F: TGATGAGAATCGCAGTAGA R: CACCAGGGAGAGTTAGAAA | NM_001024828.3 |
| Tet1 | F: AAAAGGAAGCGCTGTGAGAA F: CCACGCCAGTATGAGAATCA | XM_015278732.1 |
| Tet2 | F: CGGTCCTAATGTGGCAGCTA F: TGCCTTCTTTCCCAGTGTAGA | NM_001277794.1 |
| Target Gene/Fragment | Sequence (F: Forward, R: Reverse, 5′-3’) | Product (bp) |
|---|---|---|
| FSHR/Fragment 1 | F: CTCCATCCTACTGTAAGCCCATAA | 174 |
| R: GCAAGCAGGTGAGACCCAAG | ||
| FSHR/Fragment 2 | F: GATCTATGAAGGGGAGCAT | 153 |
| R: TCTACAGTGGAAAGGGAGC | ||
| FSHR/Fragment 3 | F: GAGATCATTTGGGTTGTGG | 261 |
| R: TTATGCTCCCCTTCATAGA | ||
| LHR/Fragment 1 | F: AGATCCACGCTTCCTCACAGTTTGGTG | 131 |
| R: CGCTGTCCCTTCTGGAGGTGTTCACT | ||
| Caspase-3/Fragment 1 | F: TTTGGTGAGGCAATGTTACG | 142 |
| R: TTTGGGCTCTGCGCTGTAAG | ||
| Caspase-8/Fragment 1 | F: CGGTAAGAGAAGAAAAGGAAAT | 193 |
| R: GAGACTGAAGAAGTAGAAAATC |
| Control | 0.1% Betaine | 0.5% Betaine | |
|---|---|---|---|
| Eggshell thickness, 0.01 mm | 32.0 ± 1.5 b | 32.3 ± 0.6 b | 41.6 ± 0.5 a |
| Shape index, % | 75.4 ± 0.49 | 76.4 ± 0.50 | 76.2 ± 0.47 |
| Yolk weight, g | 35.39 ± 0.59 | 36.8 ± 0.55 | 36.1 ± 0.49 |
| Egg white weight, g | 17.1 ± 0.27 | 17.4 ± 0.30 | 17.4 ± 0.28 |
| Yolk colour | 10.33 ± 0.17 | 10.7 ± 0.24 | 10.5 ± 0.18 |
| Item | Control | 0.5% Betaine | p |
|---|---|---|---|
| Uterus | |||
| MDA (nmol/mg protein) | 0.18 ± 0.07 | 0.25 ± 0.06 | 0.49 |
| T-SOD (U/mg protein) | 10.4 ± 0.71 | 8.7 ± 0.62 | 0.11 |
| GSH (μmol/g protein) | 65.17 ± 1.38 | 84.46 ± 3.03 | 0.001 |
| GSH-PX (μmol/g protein) | 61.96 ± 5.1 | 89.24 ± 11.01 | 0.05 |
| Ovary | |||
| MDA (nmol/mg protein) | 0.95 ± 0.18 | 1.06 ± 0.11 | 0.63 |
| T-SOD (U/mg protein) | 14.69 ± 0.54 | 17.37 ± 1.06 | 0.04 |
| GSH (μmol/g protein) | 55.70 ± 7.88 | 53.10 ± 9.96 | 0.84 |
| GSH-PX (μmol/g protein) | 46.54 ± 1.13 | 45.90 ± 3.34 | 0.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, F.; Jing, M.; Zhang, A.; Yi, J.; Zhang, Y. Effects of Dietary Betaine on the Laying Performance, Antioxidant Capacity, and Uterus and Ovary Function of Laying Hens at the Late Stage of Production. Animals 2023, 13, 3283. https://doi.org/10.3390/ani13203283
Guo F, Jing M, Zhang A, Yi J, Zhang Y. Effects of Dietary Betaine on the Laying Performance, Antioxidant Capacity, and Uterus and Ovary Function of Laying Hens at the Late Stage of Production. Animals. 2023; 13(20):3283. https://doi.org/10.3390/ani13203283
Chicago/Turabian StyleGuo, Feng, Mengna Jing, Aaoyu Zhang, Jinfan Yi, and Yanhong Zhang. 2023. "Effects of Dietary Betaine on the Laying Performance, Antioxidant Capacity, and Uterus and Ovary Function of Laying Hens at the Late Stage of Production" Animals 13, no. 20: 3283. https://doi.org/10.3390/ani13203283