
Thermal Balance in Male Water Buffaloes Transported by Long and Short Journeys

 ,
,  ,
,  , ,
, , 
 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Location
2.2. Animals
2.3. Road Characteristics and Vehicle Type
2.4. Experimental Phases
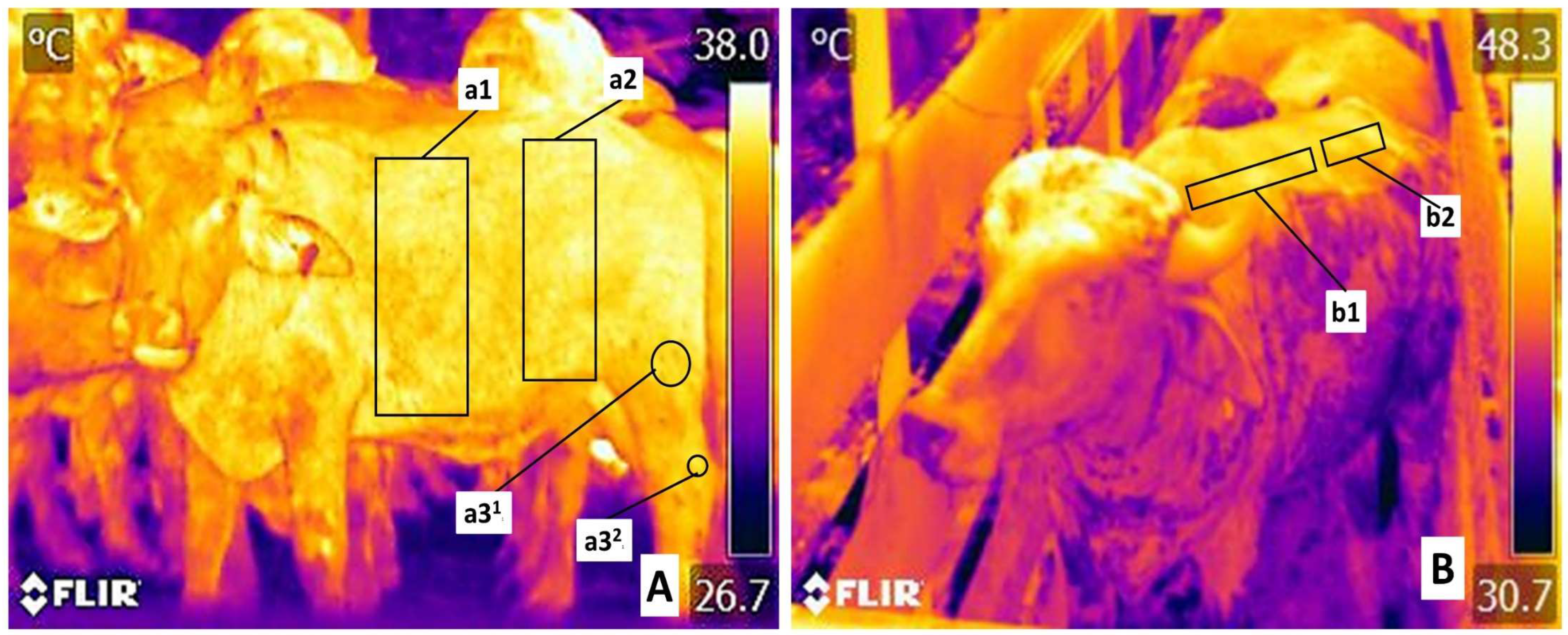
2.5. Monitoring by Infrared Thermography
2.6. Statistical Analysis
- Y = response variable (surface temperature)
- τa = effect of the transport
- τi = effect of the thermal window
- τj = effect of the event
- τk = effect of transport duration
- τiτjτk = interaction effect
- β = random effect (animal)
- µ = population mean
- e = error
2.7. Ethics Statement
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pascual-Alonso, M.; Lama, G.C.M.l.; Aguayo-Ulloa, L.; Villarroel, M.; Mitchell, M.; María, A.G. Thermophysiological, haematological, biochemical and behavioural stress responses of sheep transported on road. J. Anim. Physiol. Anim. Nutr. 2016, 101, 541–551. [Google Scholar]
- Chandra, B.S.; Das, N. Behaviour of indian river buffaloes (Bubalus bubalis) during short- ultrasonographic findings of urinary retention caused by a vaginal stricture following dystocia in a ewe. Vet. Rec. 2001, 148, 314. [Google Scholar]
- Gupta, S.; Earley, B.; Crowe, M.A. Effect of 12-hour road transportation on physiological, immunological and haematological parameters in bulls housed at different space allowances. Vet. J. 2007, 173, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, F.S.; Vaz, R.Z.; Cardoso, F.F.; Restle, J.; Vaz, F.N.; Pascoal, L.L.; Reimann, F.A.; Boligon, A.A. Pre-slaughtering factors related to bruises on cattle carcasses. Anim. Prod. Sci. 2018, 58, 385. [Google Scholar] [CrossRef]
- Guerrero-Legarreta, I.; Napolitano, F.; Cruz-Monterrosa, R.; Mota-Rojas, D.; Mora-Medina, P.; Ramírez-Bribiesca, E.; Bertoni, A.; Berdugo-Gutiérrez, J.; Braghieri, A. River buffalo meat production and quality: Sustainability, productivity, nutritional and sensory properties. J. Buffalo Sci. 2020, 9, 159–169. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Alvarez, J.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortázar Schmidt, C.; Michel, V.; et al. Welfare of cattle during transport. EFSA J. 2022, 20, e07442. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Martínez-Burnes, J.; Casas-Alvarado, A.; Gómez-Prado, J.; Hernández-Ávalos, I.; Domínguez-Oliva, A.; Lezama-García, K.; Jacome-Romero, J.; Rodríguez-González, D.; Pereira, A.M.F. Clinical usefulness of infrared thermography to detect sick animals: Frequent and current cases. CABI Rev. 2022, 17, 1–17. [Google Scholar] [CrossRef]
- Idris, M.; Uddin, J.; Sullivan, M.; McNeill, D.M.; Phillips, C.J.C. Non-invasive physiological indicators of heat stress in cattle. Animals 2021, 11, 71. [Google Scholar] [CrossRef]
- Rodríguez-González, D.; Guerrero-Legarreta, I.; Cruz Monterrosa, R.G.; Napolitano, F.; Gonçalves-Titto, C.; El-Aziz, A.H.A.; Hernández-Ávalos, I.; Casas-Alvarado, A.; Oliva-Domínguez, A.; Mota-Rojas, D. Assessment of thermal changes in water buffalo mobilized from the paddock and transported by short journeys. Front. Vet. Sci. 2023, 10, 1184577. [Google Scholar] [CrossRef]
- Cruz-Monterrosa, R.; Mota-Rojas, D.; Ramírez-Bibriesca, E.; Mora-Medina, P.; Guerrero-Legarreta, I. Scientific findings on the quality of river buffalo meat and prospects for future studies. J. Buffalo Sci. 2020, 9, 170–180. [Google Scholar] [CrossRef]
- Chulayo, A.Y.; Bradley, G.; Muchenje, V. Effects of transport distance, lairage time and stunning efficiency on cortisol, glucose, hspa1a and how they relate with meat quality in cattle. Meat Sci. 2016, 117, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Liotta, L.; Costa, L.N.; Chiofalo, B.; Ravarotto, L.; Chiofalo, V. Effect of lairage duration on some blood constituents and beef quality in bulls after long journey. Ital. J. Anim. Sci. 2007, 6, 375–384. [Google Scholar] [CrossRef]
- Alam, M.R.; Gregory, N.G.; Jabbar, M.A.; Uddin, M.S.; Kibria, A.S.M.G.; Silva-Fletcher, A. Skin injuries identified in cattle and water buffaloes at livestock markets in Bangladesh. Vet. Rec. 2010, 167, 415–419. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.R.; Gregory, N.G.; Uddin, M.S.; Jabbar, M.A.; Chowdhury, S.; Debnath, N.C. Frequency of nose and tail injuries in cattle and water buffalo at livestock markets in Bangladesh. Anim. Welf. 2010, 19, 295–300. [Google Scholar]
- Mota-Rojas, D.; Napolitano, F.; Braghieri, A.; Guerrero-Legarreta, I.; Bertoni, A.; Martínez-Burnes, J.; Cruz-Monterrosa, R.; Gómez, J.; Ramírez-Bribiesca, E.; Barrios-García, H.; et al. Thermal biology in river buffalo in the humid tropics: Neurophysiological and behavioral responses assessed by infrared thermography. J. Anim. Behav. Biometeorol. 2021, 9, 2103. [Google Scholar] [CrossRef]
- Robertson, J.K.; Mastromonaco, G.; Burness, G. Evidence that stress-induced changes in surface temperature serve a thermoregulatory function. J. Exp. Biol. 2020, 223, jeb213421. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Titto, C.G.; de Mira Geraldo, A.; Martínez-Burnes, J.; Gómez, J.; Hernández-Ávalos, I.; Casas, A.; Domínguez, A.; José, N.; Bertoni, A.; et al. Efficacy and function of feathers, hair, and glabrous skin in the thermoregulation strategies of domestic animals. Animals 2021, 11, 3472. [Google Scholar] [CrossRef]
- Damtew, A.; Erega, Y.; Hussen, E.; Tsegaye, S.; Desiye, M. The effect of long distance transportation stress on cattle: A review. Biomed. J. Sci. Tech. Res. 2018, 3, 1–5. [Google Scholar] [CrossRef]
- INEGI Clima Veracruz de Ignacio de La Llave. Available online: https://cuentame.inegi.org.mx/monografias/informacion/ver/territorio/clima.aspx?tema=me&e=30 (accessed on 5 January 2023).
- SEGOB Norma Oficial Mexicana. Señalamiento Horizontal y Vertical de Carreteras y Vialidades Urbanas; PROY-NOM-034-SCT2-2003; Diario Oficial de la Federación: Mexico City, Mexico, 2004; pp. 2–7. [Google Scholar]
- Rodríguez, A.; Romero, E. Evaluación del efecto de diferentes dosis de la fertilización orgánica en la calidad nutricional del pasto camelote (Paspalum fasciculatum willd). Tlatemoani Rev. Académica Investig. 2016, 1, 1–14. [Google Scholar]
- DOF. Trato Humanitario en la Movilización de Animales; NOM-051-ZOO-1995; Diario Oficial de la Federación: Mexico City, Mexico, 1995; pp. 1–23. [Google Scholar]
- Peeters, E.; Deprez, K.; Beckers, F.; De Baerdemaeker, J.; Auben, A.E.; Geers, R. Effect of driver and driving style on the stress responses of pigs during a short journey by trailer. Anim. Welf. 2008, 17, 189–196. [Google Scholar]
- Bozzo, G.; Barrasso, R.; Marchetti, P.; Roma, R.; Samoilis, G.; Tantillo, G.; Ceci, E. Analysis of stress indicators for evaluation of animal welfare and meat quality in traditional and jewish slaughtering. Animals 2018, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Mounier, L.; Dubroeucq, H.; Andanson, S.; Veissier, I. Variations in meat pH of beef bulls in relation to conditions of transfer to slaughter and previous history of the animals. J. Anim. Sci. 2006, 84, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, F.; Luo, Y.; Zhu, L.; Li, G. Effect of acute heat stress on adrenocorticotropic hormone, cortisol, interleukin-2, interleukin-12 and apoptosis gene expression in rats. Biomed. Rep. 2015, 3, 425–429. [Google Scholar] [CrossRef]
- Rebez, E.B.; Sejian, V.; Silpa, M.V.; Dunshea, F.R. Heat stress and histopathological changes of vital organs: A novel approach to assess climate resilience in farm animals. Sustainability 2023, 15, 1242. [Google Scholar] [CrossRef]
- Oka, T. Stress-Induced Hyperthermia and Hypothermia. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 157, pp. 599–621. ISBN 9780444640741. [Google Scholar]
- Olivier, B.; van Bogaert, M.; van Oorschot, R.; Oosting, R.; Groenink, L. Stress-Induced Hyperthermia; Elsevier B.V.: Amsterdam, The Netherlands, 2005; Volume 15, ISBN 0444518231. [Google Scholar]
- Peaston, R.T.; Weinkove, C. Measurement of catecholamines and their metabolites. Ann. Clin. Biochem. 2004, 41, 17–38. [Google Scholar] [CrossRef]
- Ghassemi Nejad, J.; Kim, B.-W.; Lee, B.-H.; Sung, K.-I. Coat and hair color: Hair cortisol and serotonin levels in lactating holstein cows under heat stress conditions. Anim. Sci. J. 2017, 88, 190–194. [Google Scholar] [CrossRef]
- Archana, P.R.; Sejian, V.; Ruban, W.; Bagath, M.; Krishnan, G.; Aleena, J.; Manjunathareddy, G.B.; Beena, V.; Bhatta, R. Comparative assessment of heat stress induced changes in carcass traits, plasma leptin profile and skeletal muscle myostatin and HSP70 gene expression patterns between indigenous osmanabadi and salem black goat breeds. Meat Sci. 2018, 141, 66–80. [Google Scholar] [CrossRef]
- Hagenmaier, J.A.; Reinhardt, C.D.; Bartle, S.J.; Henningson, J.N.; Ritter, M.J.; Calvo-Lorenzo, M.S.; Vogel, G.J.; Guthrie, C.A.; Siemens, M.G.; Thomson, D.U. Effect of handling intensity at the time of transport for slaughter on physiological response and carcass characteristics in beef cattle fed ractopamine hydrochloride. J. Anim. Sci. 2017, 95, 1963. [Google Scholar] [CrossRef]
- Chulayo, A.Y.; Muchenje, V. The effects of pre-slaughter stress and season on the activity of plasma creatine kinase and mutton quality from different sheep breeds slaughtered at a smallholder abattoir. Asian-Australas. J. Anim. Sci. 2013, 26, 1762–1772. [Google Scholar] [CrossRef]
- Hultgren, J.; Segerkvist, K.A.; Berg, C.; Karlsson, A.H.; Öhgren, C.; Algers, B. Preslaughter stress and beef quality in relation to slaughter transport of cattle. Livest. Sci. 2022, 264, 105073. [Google Scholar] [CrossRef]
- Burdick, N.C.; Carroll, J.A.; Hulbert, L.E.; Dailey, J.W.; Willard, S.T.; Vann, R.C.; Welsh, T.H.; Randel, R.D. Relationships between temperament and transportation with rectal temperature and serum concentrations of cortisol and epinephrine in bulls. Livest. Sci. 2010, 129, 166–172. [Google Scholar] [CrossRef]
- Gallo, C.; Lizondo, G.; Knowles, T.G. Effects of journey and lairage time on steers transported to slaughter in Chile. Vet. Rec. 2003, 152, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Valadez Noriega, M.; de la Lama, G.C. Implications, trends, and prospects for long-distance transport in cattle. Review. Rev. Mex. Cienc. Pecu. 2020, 11, 517–538. [Google Scholar] [CrossRef]
- Lei, M.C.; Félix, L.; Cardoso, R.; Monteiro, S.M.; Silva, S.; Venâncio, C. Non-invasive biomarkers in saliva and eye infrared thermography to assess the stress response of calves during transport. Animals 2023, 13, 2311. [Google Scholar] [CrossRef] [PubMed]
- Borell, E.H. The biology of stress and its application to livestock housing and transportation assessment. J. Anim. Sci. 2001, 79, E260. [Google Scholar] [CrossRef]
- Stewart, M.; Schaefer, A.; Haley, D.; Colyn, J.; Cook, N.; Stafford, K.; Webster, J. Infrared thermography as a non-invasive method for detecting fear-related responses of cattle to handling procedures. Anim. Welf. 2008, 17, 387–393. [Google Scholar] [CrossRef]
- Stewart, M.; Stookey, J.M.; Stafford, K.J.; Tucker, C.B.; Rogers, A.R.; Dowling, S.K.; Verkerk, G.A.; Schaefer, A.L.; Webster, J.R. Effects of local anesthetic and a nonsteroidal antiinflammatory drug on pain responses of dairy calves to hot-iron dehorning. J. Dairy Sci. 2009, 92, 1512–1519. [Google Scholar] [CrossRef]
- Stewart, M.; Verkerk, G.A.; Stafford, K.J.; Schaefer, A.L.; Webster, J.R. Noninvasive assessment of autonomic activity for evaluation of pain in calves, using surgical castration as a model. J. Dairy Sci. 2010, 93, 3602–3609. [Google Scholar] [CrossRef]
- Knizkova, I.; Kunc, P.; Gurdil, G.; Pinar, Y.; Selvi, K. Applications of infrared thermography in animal production. Anadolu Tarım Bilim. Derg. 2007, 22, 329–336. [Google Scholar]
- Schaefer, A.L.; Jones, S.D.M.; Tong, A.K.W.; Vincent, B.C. Infrared thermography of pigs with known genotypes for stress susceptibility in relation to pork quality. Can. J. Anim. Sci. 1989, 69, 491–495. [Google Scholar] [CrossRef]
- Schmitz, L.; Ebinghaus, A.; Ivemeyer, S.; Domas, L.; Knierim, U. Validity aspects of behavioural measures to assess cows’ responsiveness towards humans. Appl. Anim. Behav. Sci. 2020, 228, 105011. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Broom, D.M.; Orihuela, A.; Velarde, A.; Napolitano, F.; Alonso-Spilsbury, M. Effects of human-animal relationship on animal productivity and welfare. J. Anim. Behav. Biometeorol. 2020, 8, 196–205. [Google Scholar] [CrossRef]
- Hemsworth, P. Human–animal interactions in livestock production. Appl. Anim. Behav. Sci. 2003, 81, 185–198. [Google Scholar] [CrossRef]
- Lees, A.M.; Sejian, V.; Wallage, A.L.; Steel, C.C.; Mader, T.L.; Lees, J.C.; Gaughan, J.B. The impact of heat load on cattle. Animals 2019, 9, 322. [Google Scholar] [CrossRef]
- Sapolsky, R.M. How do glucocorticoids influence stress responses? integrating permissive, suppressive, stimulatory, and preparative actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef]
- Hernández-Avalos, I.; Mota-Rojas, D.; Mendoza-Flores, J.E.; Casas-Alvarado, A.; Flores-Padilla, K.; Miranda-Cortes, A.E.; Torres-Bernal, F.; Gómez-Prado, J.; Mora-Medina, P. Nociceptive pain and anxiety in equines: Physiological and behavioral alterations. Vet. World 2021, 14, 2984–2995. [Google Scholar] [CrossRef]
- Nakamura, K. Neural circuit for psychological stress-induced hyperthermia. Temperature 2015, 2, 352–361. [Google Scholar] [CrossRef]
- Grandin, T.; Shivley, C. How farm animals react and perceive stressful situations such as handling, restraint, and transport. Animals 2015, 5, 1233–1251. [Google Scholar] [CrossRef]
- Lowe, G.; Sutherland, M.; Waas, J.; Schaefer, A.; Cox, N.; Stewart, M. Infrared thermography—A non-invasive method of measuring respiration rate in calves. Animals 2019, 9, 535. [Google Scholar] [CrossRef]
- Stewart, M.; Wilson, M.T.; Schaefer, A.L.; Huddart, F.; Sutherland, M.A. The Use of Infrared Thermography and accelerometers for remote monitoring of dairy cow health and welfare. J. Dairy Sci. 2017, 100, 3893–3901. [Google Scholar] [CrossRef]
- Thomson, D.U.; Loneragan, G.H.; Henningson, J.N.; Ensley, S.; Bawa, B. Special report of finished feedlot cattle following transportation. J. Am. Vet. Med. Assoc. 2015, 247, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Tattersall, G.J. Infrared thermography: A non-invasive window into thermal physiology. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 202, 78–98. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Pereira, A.M.F.; Wang, D.; Martínez-Burnes, J.; Ghezzi, M.; Hernández-Avalos, I.; Lendez, P.; Mora-Medina, P.; Casas, A.; Olmos-Hernández, A.; et al. Clinical applications and factors involved in validating thermal windows used in infrared thermography in cattle and river buffalo to assess health and productivity. Animals 2021, 11, 2247. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Piedra, C.; Gomez-Milan, E.; Di Stasi, L.L. Nasal skin temperature reveals changes in arousal levels due to time on task: An experimental thermal infrared imaging study. Appl. Ergon. 2019, 81, 102870. [Google Scholar] [CrossRef] [PubMed]
- Matsuno, S.; Mizuno, T.; Asano, H.; Mito, K.; Itakura, N. Estimating autonomic nerve activity using variance of thermal face images. Artif. Life Robot. 2018, 23, 367–372. [Google Scholar] [CrossRef]
- International Committee Veterinary Gross Anatomical Nomenclature Nomina Anatómica Veterinaria. 2017, 160. Available online: https://www.wava-amav.org/index.html (accessed on 2 February 2023).
- Badakhshan, Y.; Mohammadabadi, M.R. Thermoregulatory mechanisms of jersey adult cattle and calves based on different body sites temperature. Iran. J. Appl. Anim. Sci. 2015, 5, 793–798. [Google Scholar]
- Picard, K.; Thomas, D.W.; Festa-Bianchet, M.; Belleville, F.; Laneville, A. Differences in the thermal conductance of tropical and temperate bovid horns. Ecoscience 1999, 6, 148–158. [Google Scholar] [CrossRef]
- Cain, J.W.; Krausman, P.R.; Rosenstock, S.S.; Turner, J.C. Mechanisms of thermoregulation and water balance in desert ungulates. Wildl. Soc. Bull. 2006, 34, 570–581. [Google Scholar] [CrossRef]
- Algra, M.; de Keijzer, L.; Arndt, S.S.; van Eerdenburg, F.J.C.M.; Goerlich, V.C. Evaluation of the thermal response of the horns in dairy cattle. Animals 2023, 13, 500. [Google Scholar] [CrossRef]
- Upadhyay, R.C.; Singh, A.K.; Devi, R.; Kumar, Y.; Kumar, P. Physiological changes and blood flow in murrah buffaloes during summer and winter season. J. Buffalo Sci. 2014, 3, 63–69. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Pereira, A.M.F.; Martínez-Burnes, J.; Domínguez-Oliva, A.; Mora-Medina, P.; Casas-Alvarado, A.; Rios-Sandoval, J.; de Mira Geraldo, A.; Wang, D. Thermal imaging to assess the health status in wildlife animals under human care: Limitations and perspectives. Animals 2022, 12, 3558. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, F.; Bragaglio, A.; Braghieri, A.; El-Aziz, A.H.A.; Titto, C.G.; Villanueva-García, D.; Mora-Medina, P.; Pereira, A.M.F.; Hernández-Avalos, I.; José-Pérez, N.; et al. The effect of birth weight and time of day on the thermal response of newborn water buffalo calves. Front. Vet. Sci. 2023, 10, 1084092. [Google Scholar] [CrossRef] [PubMed]
- Ewart, S. Termorregulación. In Cunningham Fisiología Veterinaria; Klein, B.G., Ed.; Elsevier: Barcelona, Spain, 2020; pp. 596–607. [Google Scholar]
- Brcko, C.C.; da Silva, J.A.R.; Martorano, L.G.; Vilela, R.A.; de Nahúm, B.S.; Silva, A.G.M.; Barbosa, A.V.C.; Bezerra, A.S.; Lourenço Júnior, J. de B. Infrared thermography to assess thermoregulatory reactions of female buffaloes in a humid tropical environment. Front. Vet. Sci. 2020, 7, 180. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rivas, P.A.; Chauhan, S.S.; Ha, M.; Fegan, N.; Dunshea, F.R.; Warner, R.D. Effects of heat stress on animal physiology, metabolism, and meat quality: A review. Meat Sci. 2020, 162, 108025. [Google Scholar] [CrossRef] [PubMed]
- Soerensen, D.D.; Pedersen, L.J. Infrared skin temperature measurements for monitoring health in pigs: A review. Acta Vet. Scand. 2015, 57, 5. [Google Scholar] [CrossRef]
- Arduini, A.; Redaelli, V.; Luzi, F.; Dall’Olio, S.; Pace, V.; Nanni Costa, L. Relationship between deck level, body surface temperature and carcass damages in italian heavy pigs after short journeys at different unloading environmental conditions. Animals 2017, 7, 10. [Google Scholar] [CrossRef]
- Carnovale, F.; Xiao, J.; Shi, B.; Kaart, T.; Arney, D.; Phillips, C.J.C. The effects of vehicle type, transport duration and pre-transport feeding on the welfare of sheep transported in low temperatures. Animals 2021, 11, 1659. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phases | Description | Characteristics |
|---|---|---|
| P1 | Paddock | Considered as basal monitoring. The animals were housed in paddocks whose feeding system is based on pastures, providing 50 g of mineral supplements per animal. In each paddock, the presence of native grasses (Paspalum fasciculatum) (dry matter = 14.7%, 56.7% neutral detergent fiber, 38.8% acid detergent fiber and 6.2% raw protein [21]. |
| P2 | Herding on horseback | Animals with and appropriate body weight were selected. Handlers moved the buffaloes with the help of horses and without the use of physical tools that could cause injuries. For both SJs and LJs, the average duration of this phase was between 25 and 35 min. |
| P3 | Corral system | Once the animals entered the pen, they remained for 12 consecutive hours (including the night before weighing, handling, and transport). The pen surface area was 630 m2 with a dirt floor. It was delimited by metal structures with 1.6 m height and two doors, of which one connected to the handling chute (P4). The animal did not receive solid food during the last 8 h but had water ad libitum. |
| P4 | Handling chute | The handling chute hallway dimensions were 1.6 m 1 m wide × 1.6 m height × 3.5 m long. It was connected to an individual weight scale and a loading ramp. The individual passing of animals through the ramp had an average duration of 50 min. |
| P5 | Loading | Buffaloes were monitored for 30–50 min during their passage on a loading ramp with anti-slip grooving with a slope of 20°. |
| P6 | Pre transport | Monitorization was performed after the animal was loaded into the truck, inside the vehicle. |
| P7 | Post transport | Thermal monitoring was carried out when the animals were still inside the vehicle, in the shade, after arriving at the reception site. |
| Phases | Description | Hours | Ambient Temperature (AT) and Relative Humidity (RH) |
|---|---|---|---|
| P1 Paddock | Prior to the buffaloes being herded by men on horseback, while they were at rest for one hour under shade | Between 17:00 and 18:00 | AT-23–25 °C RH-81–90% |
| P2 Herding | Inside the corral on the day prior to transport | Between 18:00 and 18:30 | AT-21–25 °C RH-81–88% |
| P3 Corral | Before entering the handling chute to be weighed and loaded | 08:00 | AT-20–23 °C RH-81–86% |
| P4 Chute | While the buffaloes were inside the handling chute | 09:00 | AT-22–25 °C RH-81–88% |
| P5 Loading | During loading, with the buffaloes walking up the ramp | 10:00 | AT-22–27 °C RH-81–88% |
| P6 Pre transport | After loading the truck, with the motor shut off | 11:00 | AT-22–28 °C RH-81–89% |
| P7 Post transport | Immediately after reaching the destination for both short and long trips, with the truck’s motor still turned on | SJ-11:50 ± 5.48 | AT-23–30 °C RH-83–94% |
| LJ-00:31 ± 47.32 | AT-10.5–15.7 °C RH-84–46% |
| Group | Lacrimal Caruncle | Periocular Region | Lower Eyelid | Auricular Pavilion | Nostrils | Frontal-Parietal Region | Thoracic Region | Abdominal Region | Limbs Region | Lumbar Region | Thoracic Vertebral Region | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lacrimal caruncle | SJ (n = 7383) | 0.935 * | ||||||||||
| LJ (n = 733) | 0.935 * | |||||||||||
| Periocular region | SJ (n = 7383) | 0.963 * | 1 * | |||||||||
| LJ (n = 733) | 0.989 * | 0.999 * | ||||||||||
| Lower eyelid | SJ (n = 7383) | 0.971 * | 0.998 * | 1 * | ||||||||
| LJ (n = 733) | 0.976 * | 0.974 * | 0.999 * | |||||||||
| Auricular pavilion | SJ (n = 7383) | 0.941 * | 0.997 * | 0.974 * | 1 * | |||||||
| LJ (n = 733) | 0.811 * | 0.839 * | 0.801 * | 0.999 * | ||||||||
| Nostrils | SJ (n = 7383) | 0.962 * | 0.999 * | 0.992 * | 0.999 * | 1 * | ||||||
| LJ (n = 733) | 0.989 * | 0.989 * | 0.999 * | 0.984 * | 0.999 * | |||||||
| Frontal-parietal region | SJ (n = 7383) | 0.957 * | 0.999 * | 0.991 * | 0.981 * | 0.999 * | 1 * | |||||
| LJ (n = 733) | 0.976 * | 0.977 * | 0.990 * | 0.964 * | 0.971 * | 0.999 * | ||||||
| Thoracic region | SJ (n = 7383) | 0.954 * | 0.988 * | 0.995 * | 0.987 * | 0.999 * | 0.999 * | 1 * | ||||
| LJ (n = 733) | 0.981 * | 0.977 * | 0.993 * | 0.965 * | 0.974 * | 0.999 * | 0.999 * | |||||
| Abdominal region | SJ (n = 7383) | 0.952 * | 0.989 * | 0.987 * | 0.963 * | 0.986 * | 0.999 * | 1 * | 1 * | |||
| LJ (n = 733) | 0.967 * | 0.953 * | 0.989 * | 0.948 * | 0.986 * | 0.988 * | 0.999 * | 0.999 * | ||||
| Limbs region | SJ (n = 7383) | 0.952 * | 0.999 * | 0.988 * | 0.999 * | 0.999 * | 0.987 * | 0.988 * | 0.975 * | 1 * | ||
| LJ (n = 733) | 0.996 * | 0.999 * | 0.999 * | 0.999 * | 0.976 * | 0.999 * | 0.999 * | 0.982 * | 0.999 * | |||
| Lumbar region | SJ (n = 7383) | 0.957 * | 0.999 * | 0.988 * | 0.986 * | 0.999 * | 0.999 * | 0.999 * | 0.999 * | 0.999 * | 1 * | |
| LJ (n = 733) | 0.982 * | 0.975 * | 0.999 * | 0.978 * | 0.987 * | 1 * | 0.987 * | 0.994 * | 0.976 * | 0.999 * | ||
| Thoracic vertebral region | SJ (n = 7383) | 0.945 * | 0.982 * | 0.999 * | 0.975 * | 0.983 * | 0.999 * | 1 * | 0.999 * | 0.985 * | 1 * | 1 * |
| LJ (n = 733) | 0.975 * | 0.974 * | 0.999 * | 0.963 * | 0.978 * | 0.999 * | 0.999 * | 0.999 * | 0.978 * | 0.999 * | 0.999 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-González, D.; Guerrero Legarreta, I.; Chay-Canul, A.; Hernández-Avalos, I.; Napolitano, F.; García-Herrera, R.; Pereira, A.M.F.; Domínguez-Oliva, A.; Casas-Alvarado, A.; Reyes-Sotelo, B.; et al. Thermal Balance in Male Water Buffaloes Transported by Long and Short Journeys. Animals 2023, 13, 3274. https://doi.org/10.3390/ani13203274
Rodríguez-González D, Guerrero Legarreta I, Chay-Canul A, Hernández-Avalos I, Napolitano F, García-Herrera R, Pereira AMF, Domínguez-Oliva A, Casas-Alvarado A, Reyes-Sotelo B, et al. Thermal Balance in Male Water Buffaloes Transported by Long and Short Journeys. Animals. 2023; 13(20):3274. https://doi.org/10.3390/ani13203274
Chicago/Turabian StyleRodríguez-González, Daniela, Isabel Guerrero Legarreta, Alfonso Chay-Canul, Ismael Hernández-Avalos, Fabio Napolitano, Ricardo García-Herrera, Alfredo M. F. Pereira, Adriana Domínguez-Oliva, Alejandro Casas-Alvarado, Brenda Reyes-Sotelo, and et al. 2023. "Thermal Balance in Male Water Buffaloes Transported by Long and Short Journeys" Animals 13, no. 20: 3274. https://doi.org/10.3390/ani13203274