


Transcriptomic Analysis on Pectoral Muscle of European Meat Pigeons and Shiqi Pigeons during Embryonic Development



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Tissue Collection
2.2. Hematoxylin–Eosin (H&E) Staining of Pectoral Muscle
2.3. RNA Extraction, Library Construction and Sequencing
2.4. Quality Control and Comparative Analysis
2.5. Quantification of Gene Abundance
2.6. Identification of Differentially Expressed Genes
2.7. Analysis of GO and KEGG Pathway Enrichment
2.8. Construct the Gene Interaction Network
2.9. Real-Time Quantitative PCR (RT-qPCR) Confirmation of DEGs
3. Results
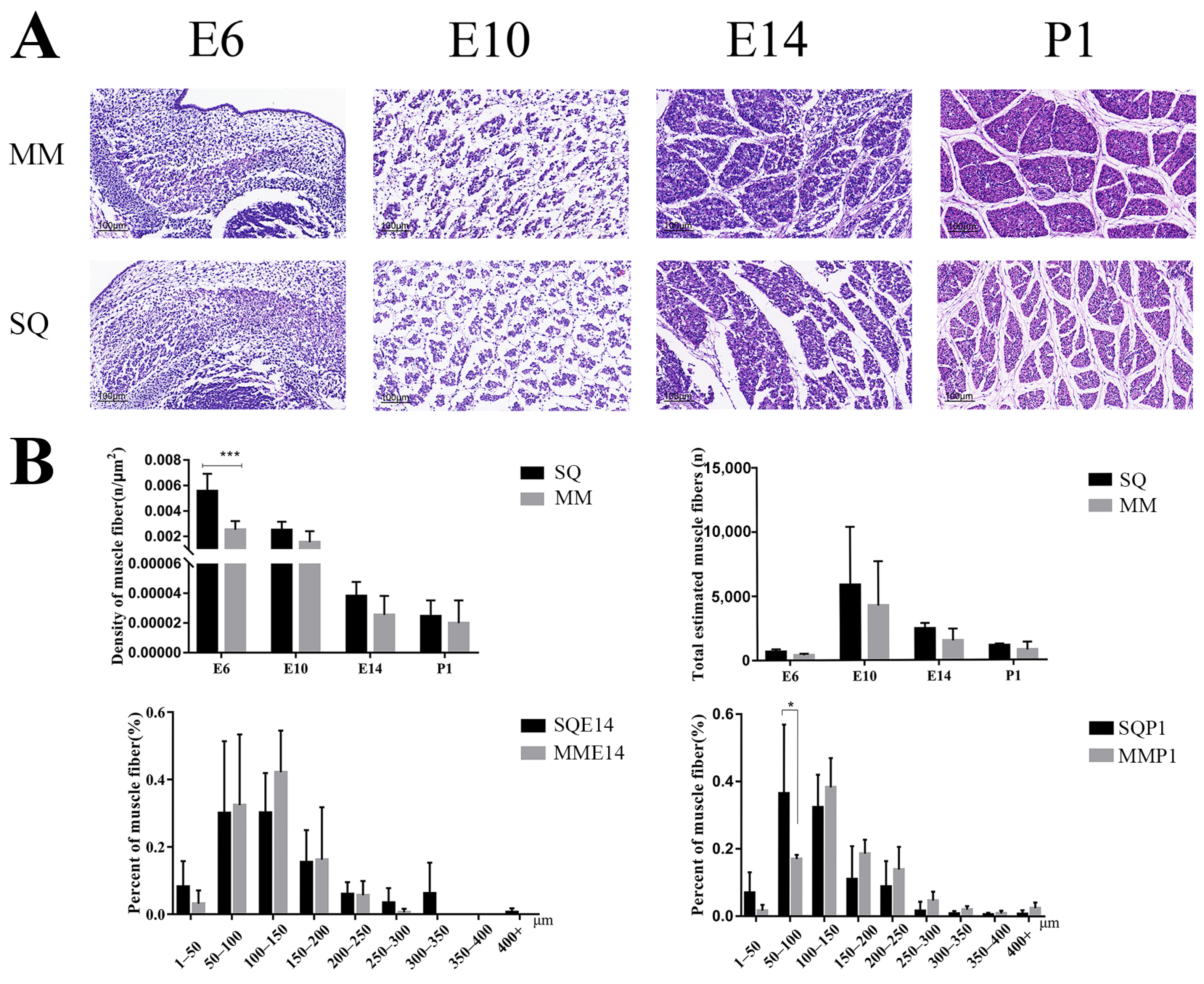
3.1. Analysis of Pectoral Muscle H&E Staining Results of Two Pigeon Breeds
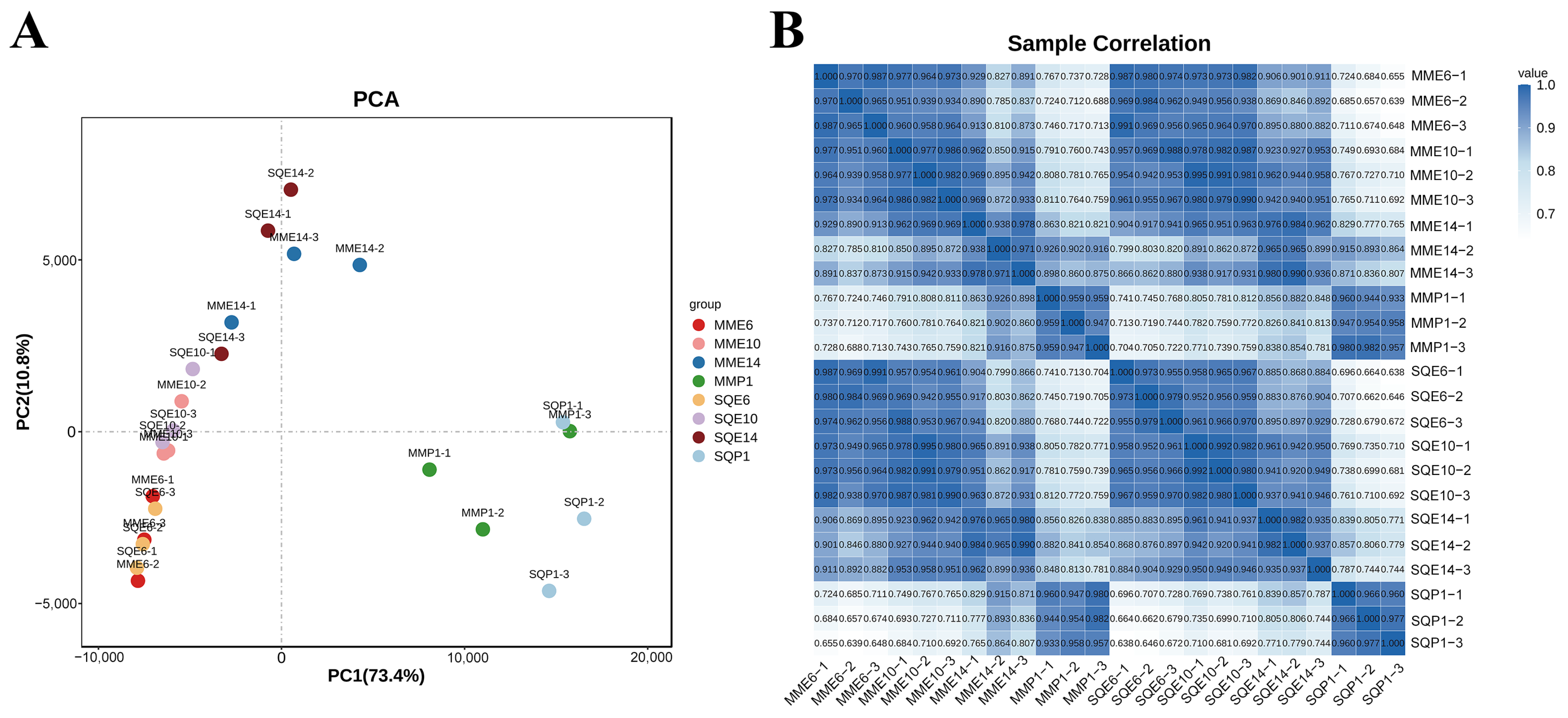
3.2. Overview of RNA-Seq
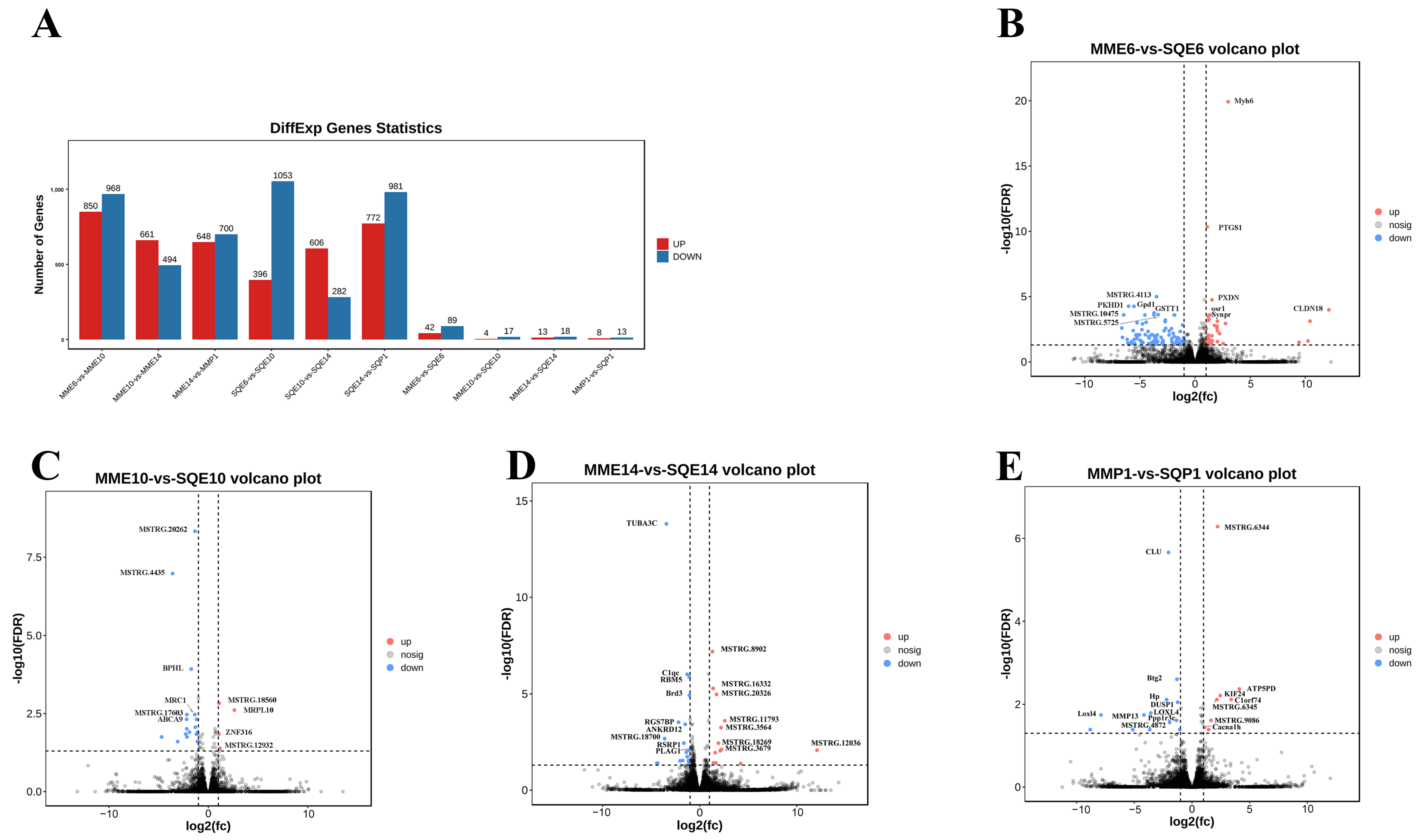
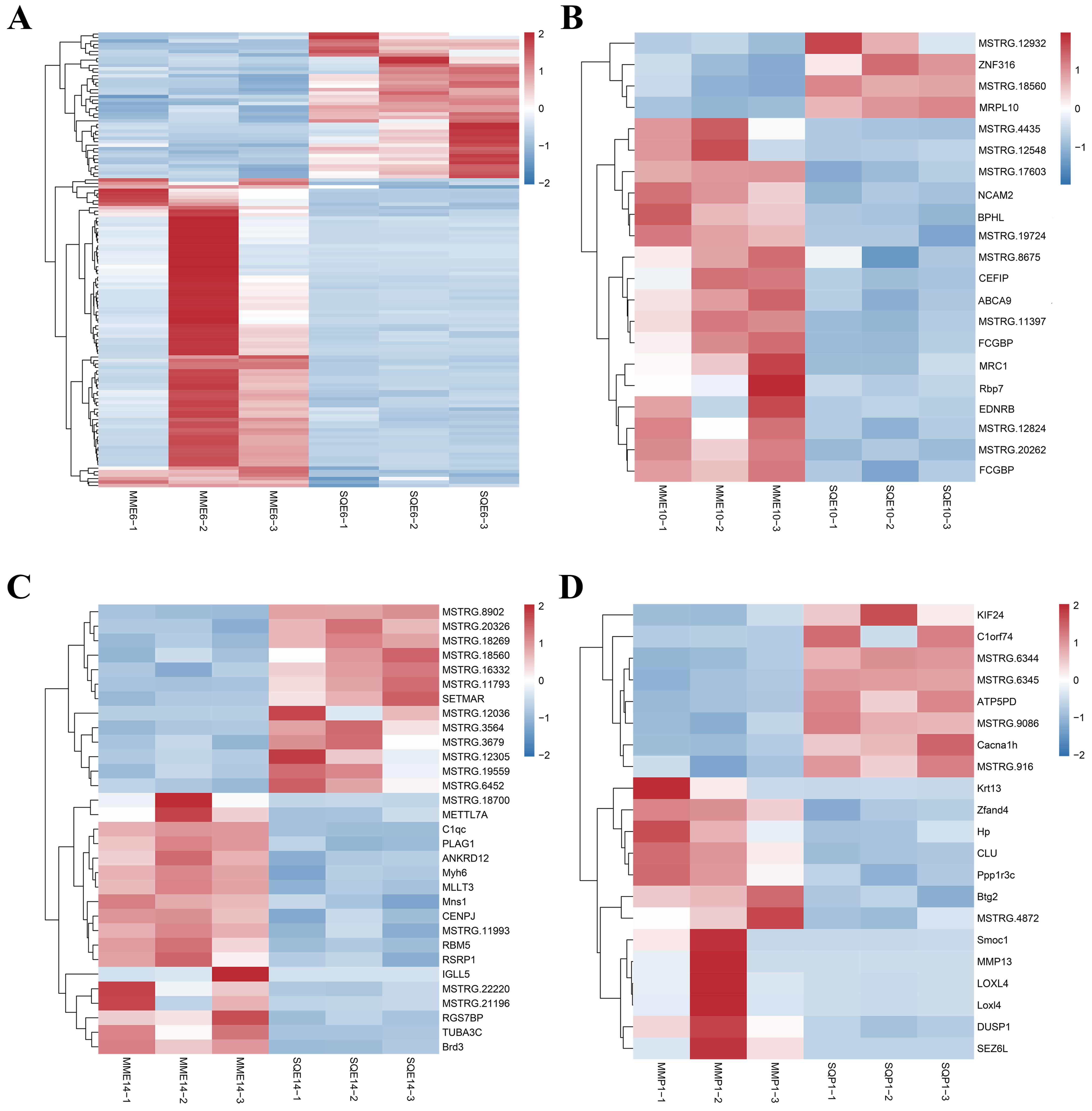
3.3. Analysis of DEGs
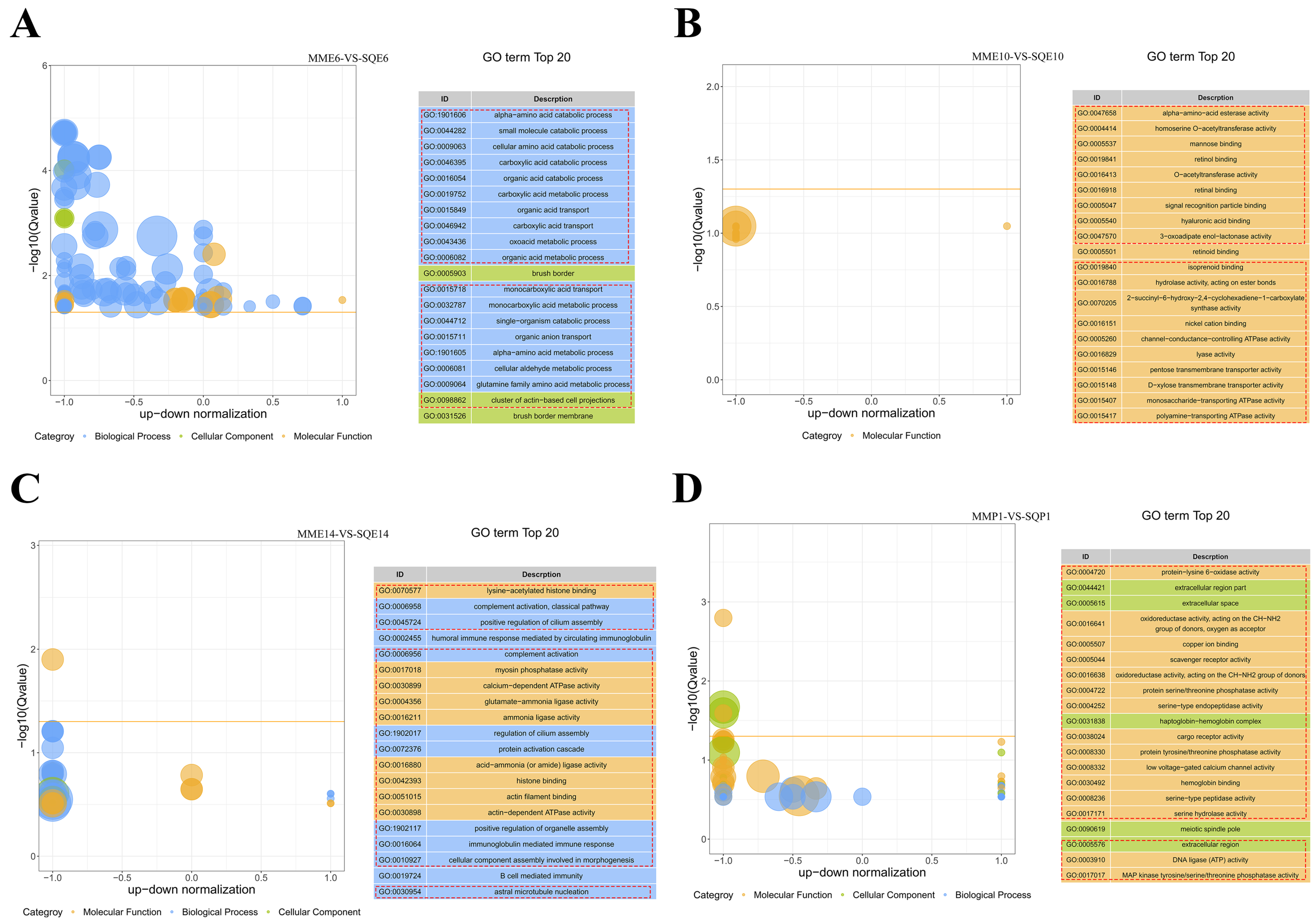
3.4. GO and KEGG Enrichment Analysis
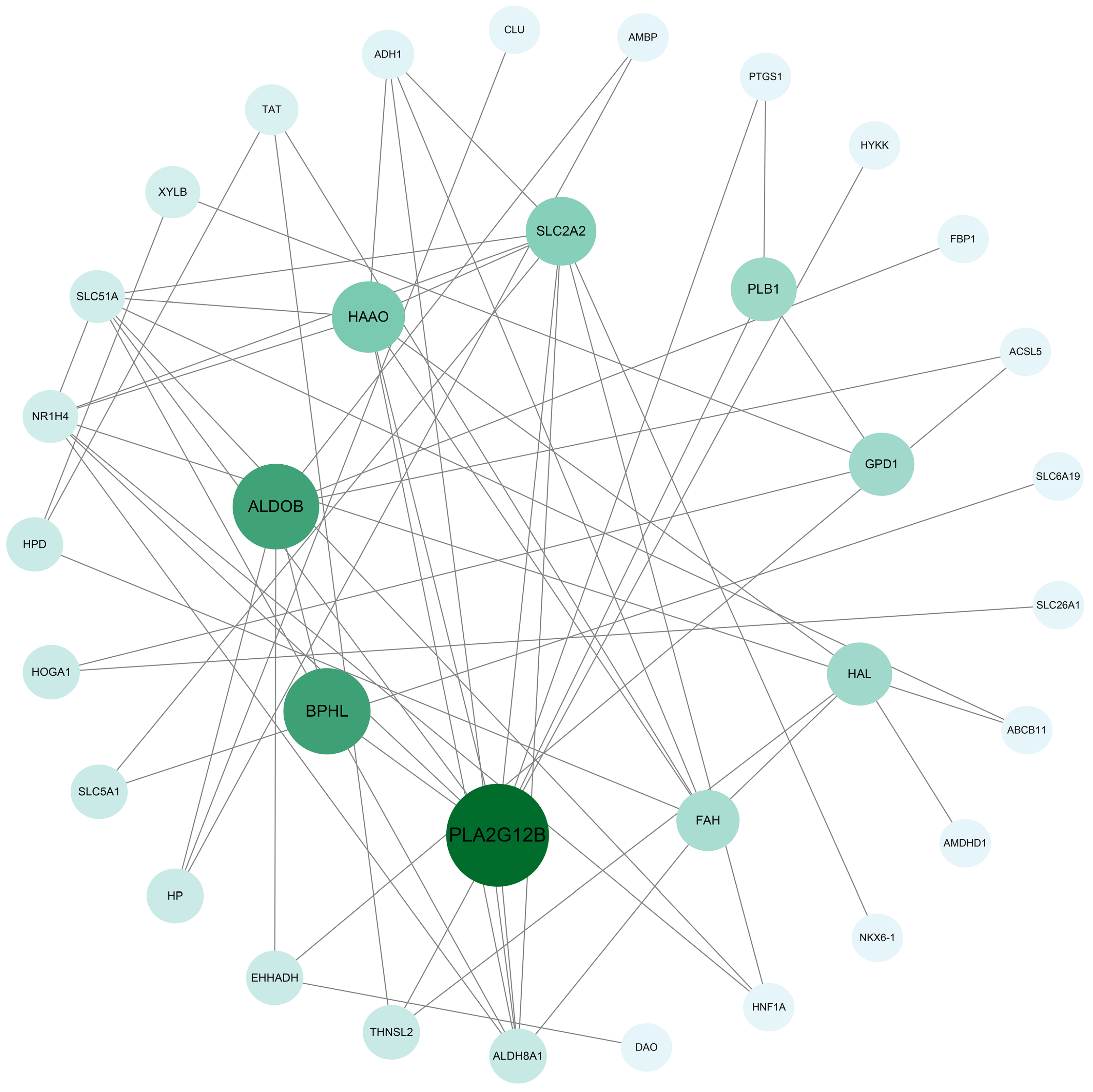
3.5. Construction of Gene Interaction Network
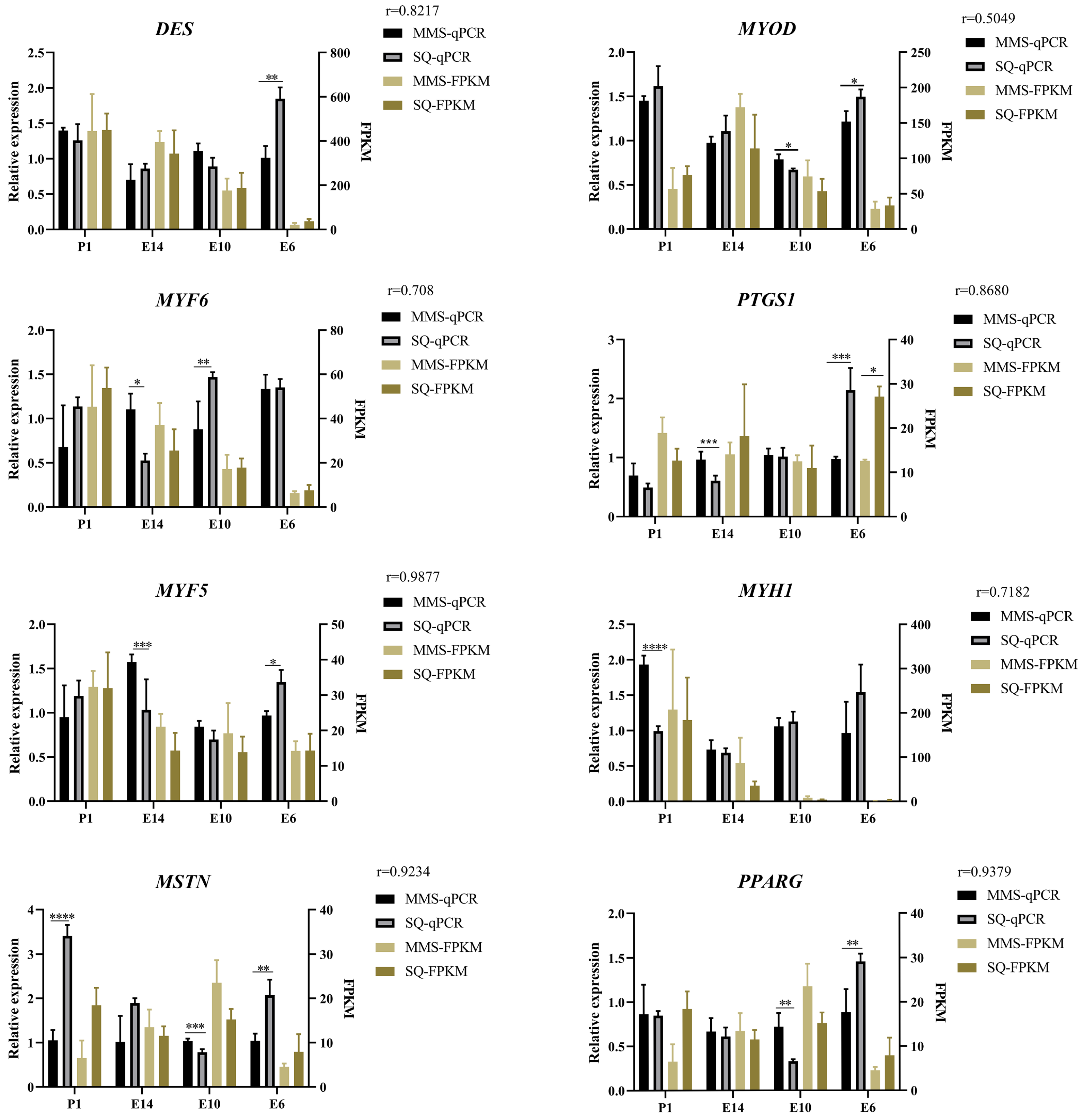
3.6. Validation of Myogenic DEGs by RT-qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, L. Introduction of excellent breeds of meat pigeons. Agriculture 2018, 15, 41–43. (In Chinese) [Google Scholar]
- Tang, Q.; Bu, Z.; Mu, C.; Chang, L.; Fu, S.; Zhang, R. Status and introduction of germplasm resources for meat pigeon breeding in China. Chin. Livest. Poult. Breed. 2018, 14, 165–168. (In Chinese) [Google Scholar]
- Wu, Z.; Mu, C.; Song, C.; Zhao, H.; Bu, Z. Measurement of body size and slaughtering of three strains of European broiler pigeons. Today Anim. Husb. Vet. Med. 2018, 34, 6–7. (In Chinese) [Google Scholar]
- Chen, M. The Growth and Nutrition Regulation of Domestic Pigeon (Columba livia). Poult. Sci. 2019, 8, 49–52. [Google Scholar]
- Guo, L.; Huang, W.; Chen, B.; Jebessa Bekele, E.; Chen, X.; Cai, B.; Nie, Q. Gga-Mir-133a-3p Regulates Myoblasts Proliferation and Differentiation by Targeting PRRX1. Front. Genet. 2018, 9, 577. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Bu, Z.; Song, C.; Mu, C.; Zhao, H. Determination of production performance of different strains of European meat pigeons. J. Domest. Anim. Ecol. 2018, 39, 73–76. (In Chinese) [Google Scholar]
- Hu, Z.; Xu, H.; Lu, Y.; He, Q.; Yan, C.; Zhao, X.; Tian, Y.; Yang, C.; Zhang, Z.; Qiu, M.; et al. MUSTN1 Is an Indispensable Factor in the Proliferation, Differentiation and Apoptosis of Skeletal Muscle Satellite Cells in Chicken. Exp. Cell Res. 2021, 407, 112833. [Google Scholar] [CrossRef]
- Buckingham, M. Gene Regulatory Networks and Cell Lineages That Underlie the Formation of Skeletal Muscle. Proc. Natl. Acad. Sci. USA 2017, 114, 5830–5837. [Google Scholar] [CrossRef]
- Zhao, P.; Hoffman, E.P. Embryonic Myogenesis Pathways in Muscle Regeneration. Dev. Dyn. 2004, 229, 380–392. [Google Scholar] [CrossRef]
- Braun, T.; Gautel, M. Transcriptional Mechanisms Regulating Skeletal Muscle Differentiation, Growth and Homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef]
- Hu, Z.; Cao, J.; Ge, L.; Zhang, J.; Zhang, H.; Liu, X. Characterization and Comparative Transcriptomic Analysis of Skeletal Muscle in Pekin Duck at Different Growth Stages Using RNA-Seq. Animals 2021, 11, 834. [Google Scholar] [CrossRef]
- Xue, Q.; Zhang, G.; Li, T.; Ling, J.; Zhang, X.; Wang, J. Transcriptomic Profile of Leg Muscle during Early Growth in Chicken. PLoS ONE 2017, 12, e0173824. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Du, H.; Yang, C.; Li, Q.; Qiu, M.; Song, X.; Yu, C.; Jiang, X.; Liu, L.; Hu, C.; et al. Comparative Transcriptome Analysis Reveals Regulators Mediating Breast Muscle Growth and Development in Three Chicken Breeds. Anim. Biotechnol. 2019, 30, 233–241. [Google Scholar] [CrossRef]
- Ding, H.; Lin, Y.; Zhang, T.; Chen, L.; Zhang, G.; Wang, J.; Xie, K.; Dai, G. Transcriptome Analysis of Differentially Expressed MRNA Related to Pigeon Muscle Development. Animals 2021, 11, 2311. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Chen, C.; Han, S.; Chen, L.; Ding, H.; Lin, Y.; Zhang, G.; Xie, K.; Wang, J.; Dai, G. Integrated Analysis Reveals a LncRNA–MiRNA–MRNA Network Associated with Pigeon Skeletal Muscle Development. Genes 2021, 12, 1787. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Li, X.; Shen, X.; Xu, D.; Tian, Y.; Huang, Y. Transcriptomic Investigation of Embryonic Pectoral Muscle Reveals Increased Myogenic Processes in Shitou Geese Compared to Wuzong Geese. Br. Poult. Sci. 2021, 62, 650–657. [Google Scholar] [CrossRef]
- Scheuermann, G.N.; Bilgili, S.F.; Tuzun, S.; Mulvaney, D.R. Comparison of Chicken Genotypes: Myofiber Number in Pectoralis Muscle and Myostatin Ontogeny. Poult. Sci. 2004, 83, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-Level Expression Analysis of RNA-Seq Experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Li, F.; Yu, Z.; Wang, Z.; Chen, W.; Xu, X.; Wang, J.; Li, X.; Wu, Z.; Zhang, X.; et al. Comparison and Transcriptome Analysis of Breast Muscle Development in Different Breeds of Pigeon Squabs. China Poult. 2023, 45, 1–10. (In Chinese) [Google Scholar]
- Chen, C.; Chen, W.; Ding, H.; Chen, L.; Zhang, G.; Xie, K.; Dai, G.; Wang, J.; Zhang, T. Developmental pattern of pectoral muscle fibers and expression analysis of related genes in embryonic stage of White King pigeon. Chin. J. Anim. Sci. 2023, 59, 99–104. (In Chinese) [Google Scholar]
- An, F.; Wang, J. Study on Differentiation during Embryonic Development across Selective and Ancestral Breeds. Anim. Sci. J. 2017, 88, 835–840. [Google Scholar] [CrossRef]
- Chen, F.; Wu, P.; Shen, M.; He, M.; Chen, L.; Qiu, C.; Shi, H.; Zhang, T.; Wang, J.; Xie, K.; et al. Transcriptome Analysis of Differentially Expressed Genes Related to the Growth and Development of the Jinghai Yellow Chicken. Genes 2019, 10, 539. [Google Scholar] [CrossRef]
- Zhang, Y.; Lv, X.; Chen, L.; Liu, Y. The Role and Function of CLU in Cancer Biology and Therapy. Clin. Exp. Med. 2023, 23, 1375–1391. [Google Scholar] [CrossRef]
- Bondesen, B.A.; Mills, S.T.; Kegley, K.M.; Pavlath, G.K. The COX-2 Pathway Is Essential during Early Stages of Skeletal Muscle Regeneration. Am. J. Physiol. Cell Physiol. 2004, 287, C475–C483. [Google Scholar] [CrossRef]
- Yang, H.; Jiang, J.; Xu, X.; He, J.; He, C.; Ma, H. Molecular Characterization, Tissue Expression Profile and SNP Analysis of the Porcine NR1H4 Gene. Mol. Biol. Rep. 2014, 41, 7009–7014. [Google Scholar] [CrossRef] [PubMed]
- Kayan, A.; Uddin, M.J.; Kocamis, H.; Tesfaye, D.; Looft, C.; Tholen, E.; Schellander, K.; Cinar, M.U. Association and Expression Analysis of Porcine HNF1A Gene Related to Meat and Carcass Quality Traits. Meat Sci. 2013, 94, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Agnetti, G.; Herrmann, H.; Cohen, S. New Roles for Desmin in the Maintenance of Muscle Homeostasis. FEBS J. 2022, 289, 2755–2770. [Google Scholar] [CrossRef] [PubMed]
- Battistelli, C.; Garbo, S.; Maione, R. MyoD-Induced Trans-Differentiation: A Paradigm for Dissecting the Molecular Mechanisms of Cell Commitment, Differentiation and Reprogramming. Cells 2022, 11, 3435. [Google Scholar] [CrossRef] [PubMed]
- Lazure, F.; Blackburn, D.M.; Corchado, A.H.; Sahinyan, K.; Karam, N.; Sharanek, A.; Nguyen, D.; Lepper, C.; Najafabadi, H.S.; Perkins, T.J.; et al. Myf6/MRF4 Is a Myogenic Niche Regulator Required for the Maintenance of the Muscle Stem Cell Pool. EMBO Rep. 2020, 21, e49499. [Google Scholar] [CrossRef]
- Murray, J.; Whitson, R.H.; Itakura, K. Reduced Prostaglandin I2 Signaling in Arid5b−/− Primary Skeletal Muscle Cells Attenuates Myogenesis. FASEB J. 2018, 32, 1868–1879. [Google Scholar] [CrossRef]
- Comai, G.; Tajbakhsh, S. Molecular and Cellular Regulation of Skeletal Myogenesis. Curr. Top. Dev. Biol. 2014, 110, 1–73. [Google Scholar]
- Ahn, J.S.; Kim, D.-H.; Park, H.-B.; Han, S.-H.; Hwang, S.; Cho, I.-C.; Lee, J.-W. Ectopic Overexpression of Porcine Myh1 Increased in Slow Muscle Fibers and Enhanced Endurance Exercise in Transgenic Mice. Int. J. Mol. Sci. 2018, 19, 2959. [Google Scholar] [CrossRef]
- Song, S.-Z.; He, Z.-Y.; Cheng, Y.; Yu, B.-L.; Zhang, T.; Li, D. MSTN Modification in Goat Mediated by TALENs and Performance Analysis. Yi Chuan 2022, 44, 531–542. [Google Scholar]
- Berger, J.; Wagner, J.A. Physiological and Therapeutic Roles of Peroxisome Proliferator-Activated Receptors. Diabetes Technol. Ther. 2002, 4, 163–174. [Google Scholar] [CrossRef]
- Dammone, G.; Karaz, S.; Lukjanenko, L.; Winkler, C.; Sizzano, F.; Jacot, G.; Migliavacca, E.; Palini, A.; Desvergne, B.; Gilardi, F.; et al. PPARγ Controls Ectopic Adipogenesis and Cross-Talks with Myogenesis During Skeletal Muscle Regeneration. Int. J. Mol. Sci. 2018, 19, 2044. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Zhu, C.; Luo, Y.; Li, S.; Wang, Q.; Han, Y.; Wu, Z.; Li, X.; Liang, Y.; Chen, Y.; et al. Transcriptomic Analysis on Pectoral Muscle of European Meat Pigeons and Shiqi Pigeons during Embryonic Development. Animals 2023, 13, 3267. https://doi.org/10.3390/ani13203267
Li F, Zhu C, Luo Y, Li S, Wang Q, Han Y, Wu Z, Li X, Liang Y, Chen Y, et al. Transcriptomic Analysis on Pectoral Muscle of European Meat Pigeons and Shiqi Pigeons during Embryonic Development. Animals. 2023; 13(20):3267. https://doi.org/10.3390/ani13203267
Chicago/Turabian StyleLi, Fada, Chenyu Zhu, Yongquan Luo, Songchao Li, Qi Wang, Yuanhao Han, Zhongping Wu, Xiujin Li, Yayan Liang, Yitian Chen, and et al. 2023. "Transcriptomic Analysis on Pectoral Muscle of European Meat Pigeons and Shiqi Pigeons during Embryonic Development" Animals 13, no. 20: 3267. https://doi.org/10.3390/ani13203267