1. Introduction
The dusky grouper (
Epinephelus marginatus) is a marine teleost in a vulnerable condition on the Red List of the International Union for Nature Conservation (IUNC) [
1,
2]. Late sexual maturation [
1,
3,
4,
5], overfishing, and especially the lack of adequate and specific management [
2] are factors that intensify the species’ natural conservation status. The consumer demand for this species is mainly due to the positive commercial characteristics presented, such as the large size, and fillets with great texture, color, and flavor. In addition to these characteristics,
E. marginatus is management-resistant, making it a promising species for Brazilian aquaculture, enabling their production in captivity and reducing extractive fishing [
1,
4,
5,
6,
7,
8,
9,
10]. However, information related to the physiology/nutrition/aquaculture of this species is scarce in the literature, as species–specific studies are necessary to assist their rearing in captivity, mainly on topics related to the reduction of the total cost of production.
Captive production emerges as a viable alternative for the conservation of
E. marginatus; however, as observed for other teleost species commonly cultivated, the production of specific diets considering the physiological characteristics of the species can be the main limiting factor for the increase in the production of this species, since most of the production costs (approximately 70% of the total cost) are directly related to nutrition [
11]. The high cost of production is directly linked to use of ingredients such as fish meal and oil, which are expensive ingredients and unavailable on the world market. Additionally, it is highly impactful on the environment, as part of these inputs are produced from natural fishing stocks [
12,
13,
14,
15]. In recent decades, there has been a striving effort to replace, mainly, fish oil (FO) with vegetable oils (VOs), such as soybean, coconut, and olive oil. However, the main limitation for the use of these alternative lipid sources in marine fish nutrition is related to their low levels of long-chain polyunsaturated fatty acids of the n-3 series (LC-PUFA n-3) [
14,
16,
17], since, in general, marine fish have limited capacity to synthesize these fatty acids from their precursors, which are acquired exclusively through diet [
6,
15,
18]. In fact, the influence of LC-PUFA n-3 on the physiology of
E. marginatus does not exist in the specialized literature.
Docosahexaenoic acid (DHA, 22:6n-3) and eicosapentaenoic acid (EPA, 20:5n-3) are the physiologically most relevant LC-PUFAs for marine fish. These fatty acids (FAs) play fundamental roles in marine fish, and adequate levels of these substrates are essential for proper development since the inadequate incorporation of these FAs in diets can negatively influence important physiological processes, such as tissue development, the prevention of various pathologies, and the balance of the immune system, in addition to impacting, in general, the lipid metabolism of the target species [
13,
14,
15,
19,
20]. Additionally, the lack of these ingredients in diets can result in a low-quality final product, generating a fillet with low nutritional value, as these FAs are relevant to human nutrition [
13,
16]. In addition to the adequate balance of DHA and EPA in diet, factors such as temperature can reduce the quality of the final product, since temperature can affect nutrient intake and the way in which fish use the energy acquired through diet, since most teleosts are ectotherms [
13,
15,
17]. Furthermore, DHA and EPA are LC-PUFAs present in biological membranes, and, because they have a specific chemical structure, they constitute fluidity [
15]. Fluidity is an important factor for cell membranes, as the greater the fluidity, the more correctly and quickly intra- and extracellular communications are carried out, facilitating the biological processes mentioned above. It is important to highlight that temperature can affect the fluidity of membranes in different ways: (1) low temperature = ordered liquid state, which is the semi-solid state in the bilayer; and (2) high temperatures = disordered liquid, which is the most fluid state in the bilayer) [
13,
15]. Additionally, the fluidity of membranes varies according to the different physiological processes mentioned [
13,
14,
15,
19,
20].
In addition to the important properties previously mentioned, DHA and EPA are among the main constituents of phospholipids, which are essential lipids that control the fluidity of the biological membranes of important tissues such as the nervous and visual ones, being essential for the structuring and functioning of cells in these and other tissues [
13,
21,
22]. Phospholipids are known to be structural components of biological membranes that respond to changes in temperature [
23,
24]. Therefore, the FA composition of this lipid class is relevant in rearing marine fish, since most cultivated species are exposed, at some period of the production cycle, to suboptimal temperatures. As, normally, the composition of the diet offered to farmed fish together with abiotic factors, especially temperature, directly influence the composition of tissues and, consequently, the growth performance, health, and quality of the final product, investigations focused on this interface on species with high productive potential and which are ecologically pressure, such as as
E. marginatus, are considered essential to improve its productivity.
Considering all the information previously described, the commercial production of E. marginatus can be considered a viable alternative for reducing the pressure on the species’ natural stocks; however, nutrition is still seen as the main barrier to large-scale production. Given the importance of DHA, EPA, and the relationship of dietary lipid composition with water temperature in the physiology and growth performance of marine fish, and, due to the lack of information related to these aspects in the nutrition of E. marginatus, this study proposed to investigate whether the effects of different DHA/EPA ratios can interfere with the growth performance and FA composition (in triacylglycerol and phospholipids) of the lipolytic and lipogenic tissues of E. marginatus juveniles. Additionally, considering the role of the liver as an important lipid deposit in the teleost, the morphology of this tissue was qualitatively evaluated.
2. Materials and Methods
2.1. Diet Manufacturing
The diet formulation and proximate composition are presented in
Table 1, while the FA profile of the experimental diets and oils used for manufacturing are shown in
Table 2. The fish and poultry meals used were previously defatted by immersion and washing in hexane (fish and poultry meals 3:1 solution). Four isoproteic, isolipidic, and isoenergetic diets were formulated containing different lipid sources. The experimental diets were formulated using a lipid blend containing coconut oil (source of saturated fatty acids—SFA), olive oil (source of monounsaturated fatty acids—MUFA), concentrated arachidonic acid (ARA) oil, and different inclusion levels of DHA and EPA concentrated oil. Each diet consisted of one dietary treatment, including equal levels of coconut, olive, and ARA oils, varying in the levels of EPA and DHA concentrated oils:
- -
D1: 1.00% EPA and 0.50% DHA;
- -
D2: 0.75% EPA and 0.75% DHA;
- -
D3: 0.50% EPA and 1.00% DHA;
- -
D4: 0.00% EPA and 1.50% DHA.
The diets were manufactured, and the experimental-blend oils were vacuum-coated to the pellets during the mixing process. A vacuum pump was attached to evacuate the air, and the lipid-blend oils were infused into the pellets when the atmospheric pressure was re-equilibrated. This method was performed according to previous studies with other marine teleost species [
17,
25]. After the vacuum-coated process, the pellets were dried in forced air circulation stoves for 24 h, and, subsequently, stored at −20 °C until the beginning of the experiment. The proximate composition of the experimental diets was analyzed in triplicate according to the methodology implemented by [
26] (
Table 1).
2.2. Experimental Design, Fish Handling, Productive Performance, and Biological Index
Epinephelus marginatus juveniles were obtained from Redemar Alevinos (Ilhabela, North Coast of São Paulo State, Brazil) and transported to the Marine Experimental Facilities at the Centre of Marine Biology of the University of São Paulo (CEBIMar/USP), also located in São Sebastião, North Coast of São Paulo State, Brazil. The fish were maintained for two weeks in a 10,000 L tank and hand-fed twice a day (08:00 and 17:00 h) until apparent satiation with a commercial marine fish diet (Guabipirá, Guabi Nutrição e Saúde Animal SA, SP, Brazil). After that, the animals were transferred to twenty-four 500L experimental tanks in a recirculation system equipped with a UV, biological filter, mechanical filter, pump, and water heater. In this study, two temperatures were considered. Twelve tanks were maintained at an average temperature of 28.63 ± 0.06 °C (triplicate for each experimental diet, namely, higher temperature—HT), and another twelve tanks were kept at an average temperature of 23.17 ± 0.12 °C (triplicate for each experimental diet, namely, lower temperature—LT). Seawater was supplied continuously at a flow of ~10 L min−1 (both temperatures), the water salinity was 31 PSU (both temperatures), the dissolved oxygen was 4.55 ± 0.06 mg L−1 (HT) and 5.30 ± 0.03 mg L−1 (LT), the total ammonia nitrogen (TAN) was lower than 0.05mg L−1 (both temperatures), the nitrite level was lower than 0.05 mg L−1 (both temperatures), and the nitrate level was lower than 1.20 mg L−1 (both temperatures). The water temperature and dissolved oxygen were measured daily (YSI model 55, YSI Inc., Yellow Springs, OH, USA), and the total ammonia nitrogen, nitrite and nitrate levels were measured every three days (API test kits, Mars Fishcare Inc., Chalfont, PA, USA).
At the beginning of the experiment, each tank held 9 fish with an average body mass of 38.85 ± 0.70 g (n = 9 animals per tank; n = 27 animals per experimental diet group; n = 108 animals per temperature group (HT and LT); n = 216 total animals used in this study). Additionally, ten animals from the initial stock were anesthetized with 0.1% benzocaine (ethyl-p-aminobenzoate) and samples of liver, muscle, the whole brain, and both eyes were immediately frozen in liquid nitrogen and maintained at −80 °C until analyses. During the nutritional experiment, fish were hand-fed at 08:00 and 17:00 h with the different diets at a rate of 3% biomass/day, for eight weeks (56 days), and this rate was adjusted every fifteen days. At the end of the experiment trial, all animals were anesthetized with 0.1% benzocaine, and their biometrical parameters were recorded. Then, three fish per tank (nine fish/experimental diets at each temperature) were euthanized and dissected. Thereafter, the liver was quickly removed and weighed to calculate the hepatosomatic index (HIS). Samples of liver and muscle, the whole brain, and both eyes were frozen in liquid nitrogen and maintained at −80 °C until analyses. Other samples of the liver were fixed in formalin solution (4%) for 24 h, and then preserved in 70% ethanol until the histological analyses.
The productive performance and biological index were evaluated using the following parameter metrics:
Weight gained (WG, g) = Wf − Wi;
Daily gain rate (DGR, g/day) = (Wf − Wi)/t;
Specific growth rate (SGR, %d) = (LnWf − LnWi) × 100/t;
Hepatosomatic index (HSI, %) = 100 × (liver weight/Wf);
Survival rate (SR, %) = Nf × 100/Ni.
where:
Wf and Wi = final (Wf) and initial (Wi) fish weight, respectively;
Nf and Ni = final (Nf) and initial (Ni) number of fish, respectively;
t = duration of experiment, in days.
Fish handling and the procedures used in the sampling of the animals were performed in accordance with the Animal Ethics Committee of the Institute of Biosciences, University of São Paulo (IB/USP) (CEUA protocol 356/2019).
2.3. Total Lipids and Fatty Acid Profile Analyses
The total lipids of the experimental diets and tissues were extracted with a mixture of chloroform, methanol, and water (respectively, 2:1:0.5) following [
27] modified by [
28] for aquatic organisms. Next, the total lipid content was quantified by the colorimetric method of [
29], using cod liver oil (cod liver oil fatty acid methyl esters, Sigma Diagnostics, St. Louis, MO, USA) to yield the standard curve. The extracted lipids from the liver, muscle, brain, and eyes were separated into triacylglycerol (TG) and phospholipid (PL) fractions by thin-layer chromatography using an activated silica column [
30]. Triacylglycerol and phospholipid fractions of liver, muscle, brain, and eyes were methylated with acetyl chloride (5% HCl in methanol) and converted into fatty acid methyl-esters [
31].
Fatty acid methyl-ester (FAME) analysis was carried out in a Varian gas chromatograph (GC; Model 3900, Walnut Creek, CA, USA) coupled with a flame ionization detector (FID) and a CP-8410 autosampler. The capillary column for analyzing the FA was a CP Wax (30 m length × 0.25 mm inner diameter with 0.25 μm thickness) using hydrogen as the carrier gas at a linear velocity of 22 cm/s. The following temperature program was used: 170 °C for 1 min, followed by a ramp of 2.5 °C/min until reaching 240 °C, and a final waiting time of 5 min, totaling 31 min of running. In the injector, the temperature was 250 °C and, in the flame ionization detector (FID), the temperature was 260 °C. Using known methyl-ester standard FAME (Supelco, 37 components; Sigma-Aldrich, St. Louis, MO, USA; Mixture, Me93; Larodan and Qualmix, PUFA fish M, Menhaden Oil, Larodan), FAs were identified by comparison with retention time. Data were presented as a percentage of the total FAME based on the peak area analyzed.
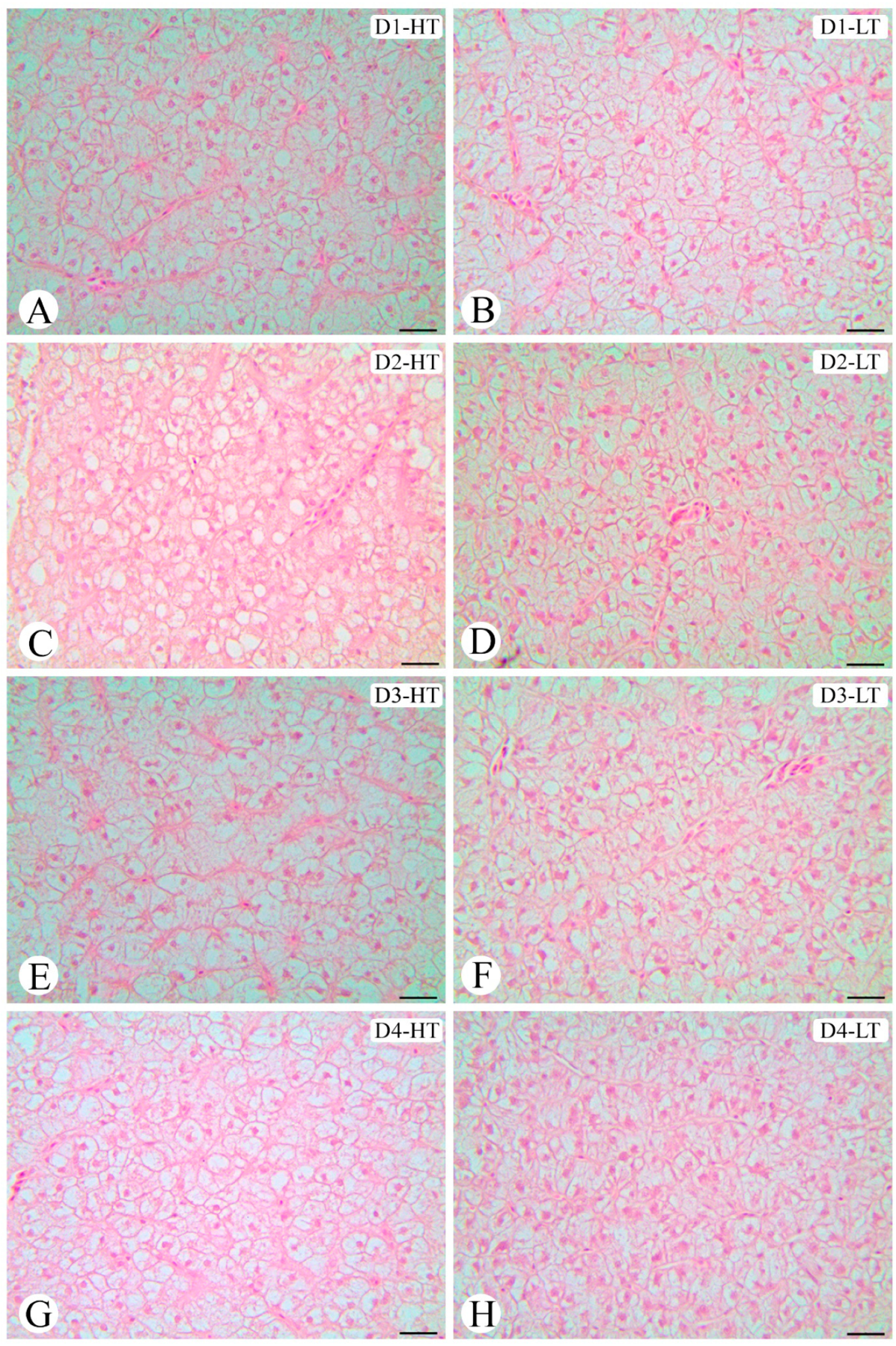
2.4. Liver Histology Analysis
Liver samples were fixed in formalin solution (4%) for 24 h, dehydrated through an ascending series of increasing concentrations of ethanol, cleared in dimethylbenzene solution (xylene), and embedded in Paraplast
® according to routine histological procedures for the preparation of liver permanent histological cross-sections [
32]. Liver sections (5 μm thick) were obtained using a microtome (Leica Histo Core Auto Cut), equipped with disposable blades and mounted on Poly-
L-Lysine-solution-coated slides. The slides were stained with hematoxylin–eosin, examined, and documented using, respectively, a light microscope (Leica DM1000 LED light microscope) coupled to a camera (Leica MC170HD photography camera) and a computerized image capture system (Leica LAS Interactive Measurements; 1260 pixels by 960 pixels) to evaluate the hepatocyte morphology and to measure the average area of these cells.
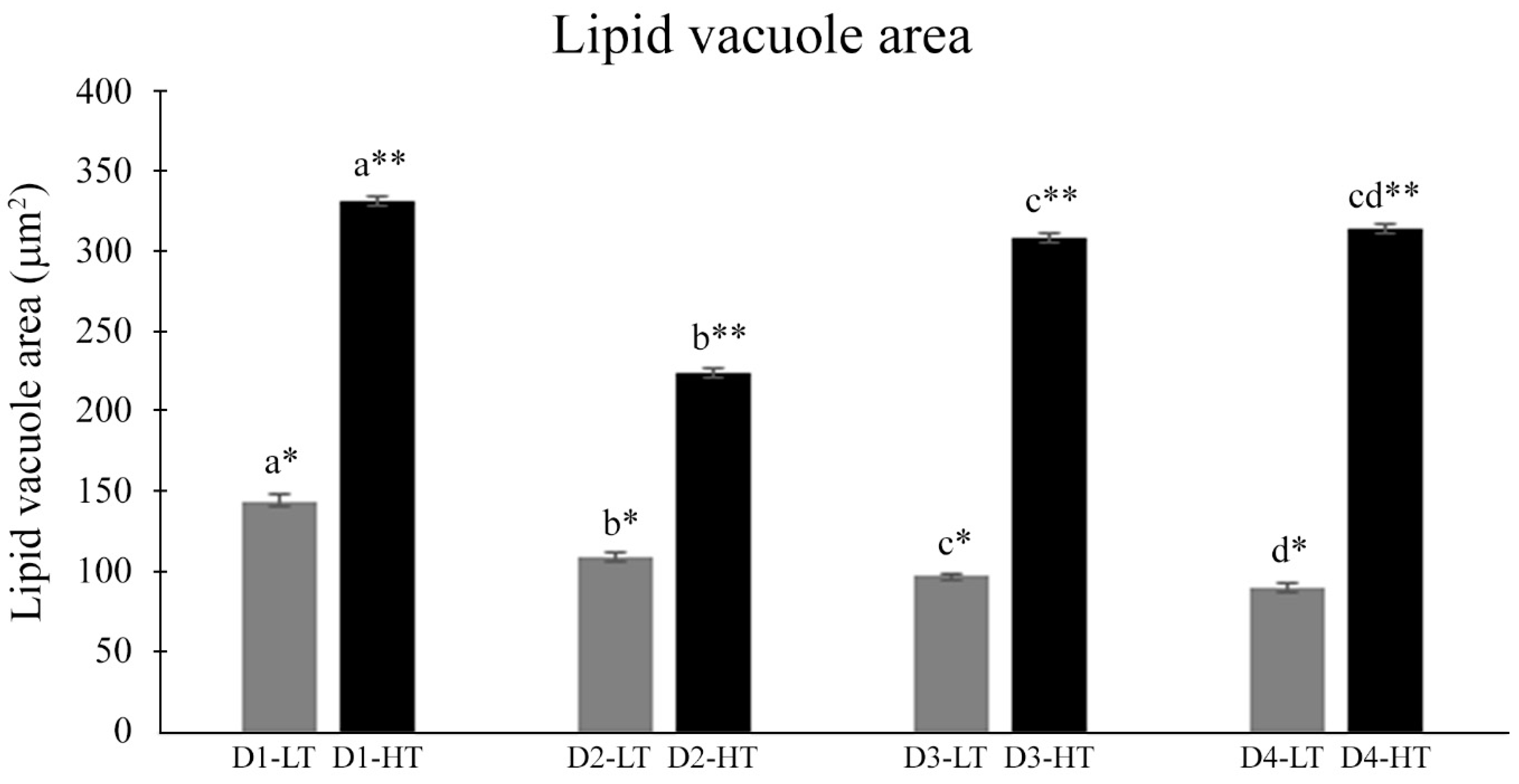
For the hepatocyte morphological analyses, the average area of these cells was considered. Within this purpose, fifteen cells per section were quantified, and three sections per sample (
n = 3 fish/tank;
n = 45 cells/fish measured; 135 cells per experimental group) with 100 μm distance between each section (totaling 300 μm distance from the first section to the last part). The “LAS Interactive Measurements” was also used to obtain the measurements of the average hepatocyte area (in μm
2) according to previous studies performed with teleost and invertebrate’s species, such as
Rachycentron canadum [
17,
25],
Morone saxatilis [
33],
Steindachneridion parahybae [
34,
35], and
Litopenaeus vannamei [
36].
2.5. Statistical Analyses
A descriptive analysis was performed, and all values were expressed as the mean ± standard error of the mean (M ± SEM). Data normality and homogeneity of variance were analyzed, and comparisons between experimental groups were performed using two-way ANOVA, considering diet and temperature as factors, and followed by Tukey’s HSD tests using the statistical software SigmaStat for Windows ver. 3.10 (Systat Software, San Jose, CA, USA). Statistical difference was used considering a significance level of 5% (p < 0.05).
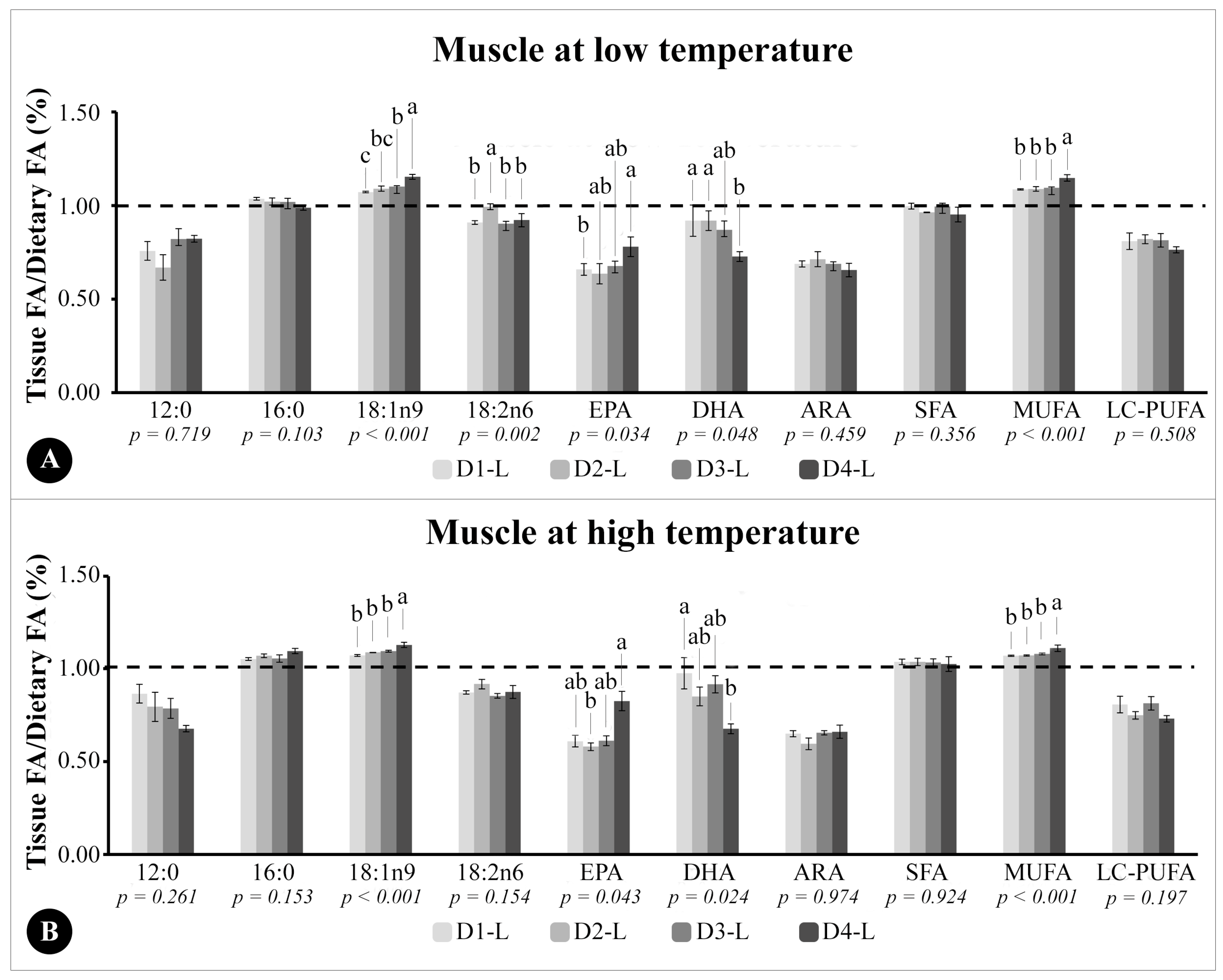
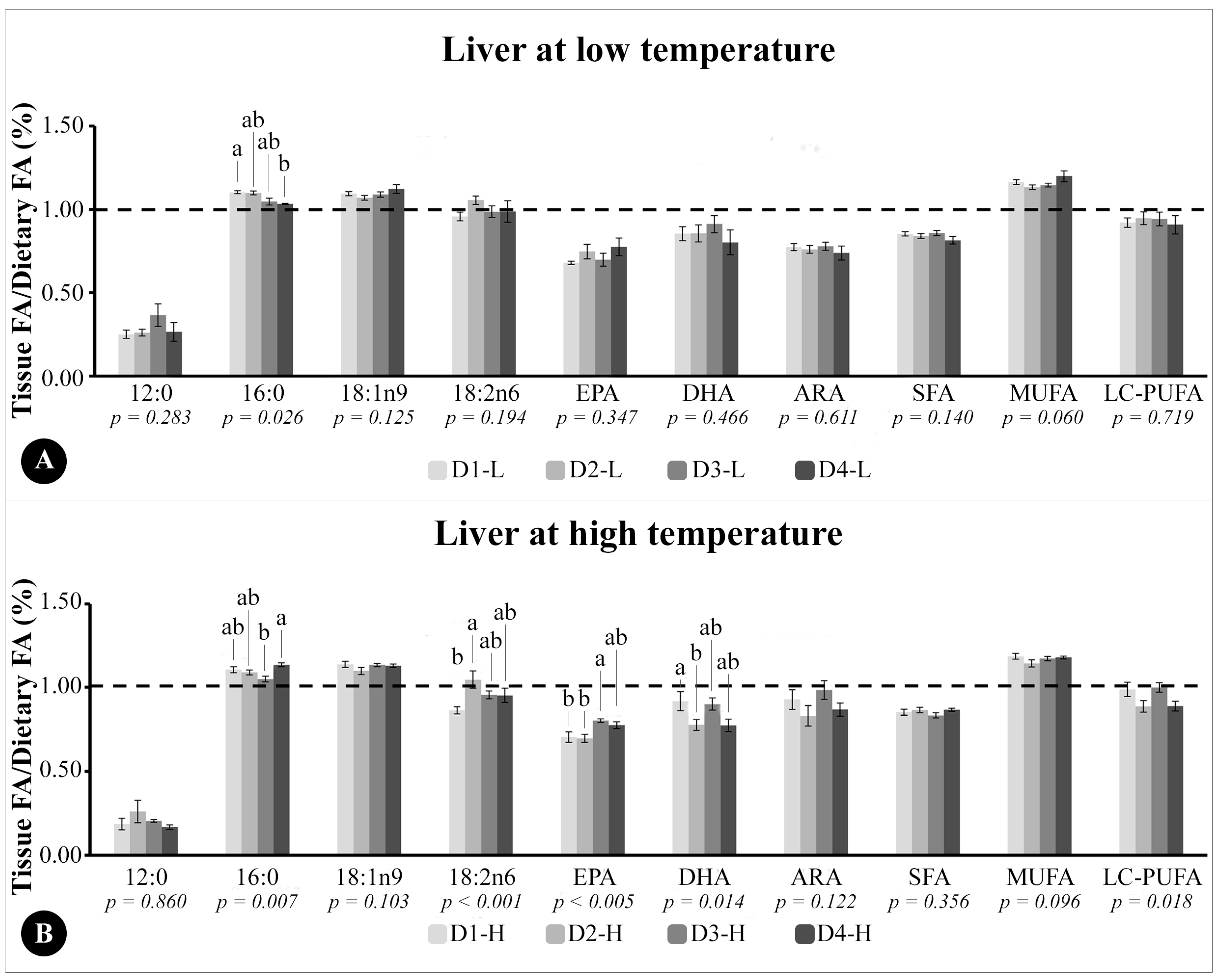
The ratio between tissue FA composition and dietary FA composition (tissue/diet) in liver and muscle TG was calculated as follows: the mean (%) of a specific FA divided by the mean (%) of the same FA in each tissue. The dotted line in the graphics indicates higher (above 1.0), lower (below 1.0), or equal levels (1.0) of a specific FA deposited in the tissues relating to the same FA in the diet. This ratio was calculated for muscle and liver tissues (and not for eyes and brain), since they are FA storage tissues and of interest (mainly muscle) for aquaculture, according to previous studies [
25].
4. Discussion
The results obtained in this study showed that different dietary DHA/EPA levels did not affect the growth performance of
E. marginatus at the lower temperature but affect the growth performance at the higher temperature, with a better DGR and WG in fish fed with the higher DHA level. This result suggests that dusky grouper has a lower nutritional requirement of DHA when kept at 23.17 ± 0.12 °C, possibly not being necessary to include this FA in greater levels in the colder seasons, such as autumn and winter. However, at 28.63 ± 0.06 °C, the species showed better growth performance when fed with a higher level of DHA, possibly indicating that, in warmer seasons, such as spring and summer, a greater inclusion of DHA is necessary for the better growth of juveniles. Previous studies with
Salmo salar [
37],
Oncorhynchus mykiss [
38],
R. canadum [
17,
39], and
M. saxatilis [
33] also demonstrated different requirements of LC-PUFA in the diet according to rearing conditions, such as temperature. These factors can directly influence species growth, also impacting LC-PUFA levels in tissues, as observed in
Tor putitora, a tropical fish species that showed higher levels of DHA at a high temperature (32 °C) [
40]. Jobling and Bendiksen [
23] further describe that, in fish, as ectothermic animals, temperature can define the rates of chemical reactions that occur in these organisms, and, thus, dictate the speed with which physiological processes occur. As a result, several metabolic processes, such as cellular respiration and oxygen consumption, may undergo changes. Therefore, the standard metabolic rate (SMR) can be affected (high temperature = high SMR; low temperature = low SMR), while the maximum metabolic rate (MMR) can vary and stabilize. And when animals are able to maintain their vital activities, much of the energy is directed towards functions such as growth, for example.
It is important to highlight that all diets in this experiment were fish-oil-free and supplemented with VO (coconut oil and olive oil), and this FO substitution did not impact the growth performance and survival of
E. marginatus juveniles. Studies with different fish species, such as
Argyrosomus regius [
41],
Totoaba mcdonald [
16],
Trachinotus ovatus [
42],
R. canadum [
25], and even species of the genus
Epinephelus [
18,
43,
44,
45,
46], also addressed the importance of replacing FO with VO, suggesting that different species have specific physiological requirements for DHA and EPA. Therefore, although the inclusion of FO is a traditional practice in marine fish nutrition, it is possible to replace it with VO since the diets were adequately supplemented with LC-PUFA n3, such as DHA and EPA [
14], as performed in this study. Moreover, FA, such as DHA and EPA are essential for marine fish, especially in captivity, as they depend on food to obtain them and cannot synthesize them again [
15]. These LC-PUFA can be used to obtain energy through β-oxidation but are mainly present in biological membranes. β-oxidation is a four-step process that results in acetyl-CoA or ketone bodies. These molecules are made available to the organism for various processes and tissues, which can use them in the most optimized way possible, such as in the process of growth and reproduction. Nervous tissue, for example, can use ketone bodies in the absence of glucose to obtain energy. The energy obtained aids growth as the organism has sufficient energy sources in the system to distribute as needed. Furthermore, as previously stated, DHA and EPA are FAs present in biological membranes that configure the fluidity and proper functioning of the lipid bilayer, as mentioned in the literature referenced throughout the manuscript. Furthermore, the fluidity in cell membranes gives them the ability to fuse, and among the processes that we can mention are endo- and exocytosis, the fertilization of the egg by a sperm, cell division, and other processes [
13]. Other factors are important for the occurrence of physiological processes, such as temperature, which can be an ally, favoring and facilitating the progress of processes, or can trigger imbalances in these processes, making their occurrence difficult. As the environmental temperature is modified, one of the only cellular components that can be remodeled is the plasma membrane, through the remodeling/restructuring of the lipid composition, called homeoviscosal adaptation [
12]. The LC-PUFA EPA, DHA, and ARA help maintain membrane fluidity at low temperatures. Due to their low melting point, they occupy more space in the lipid bilayer, increasing the fluidity and stability of the plasma membrane [
12,
13]. When the temperature is high, the membrane becomes even more fluid. Saturated fats, such as those provided by coconut oil, tend to become more rigid at temperatures of 25°C, while unsaturated fats, such as those provided by olive oil, tend to remain fluid; the same happens with the LC-PUFA DHA and EPA, which are unsaturated [
14]. Therefore, fish fed diets supplemented with LC-PUFAs may present greater thermal tolerance, maximizing growth and survival, since the constitution and functioning of cell membranes can directly affect the maintenance of physiological homeostasis and the immune system of fish [
12,
13,
14].
In the TG fraction of muscle, DHA and EPA levels reflected dietary levels; however, the T/D ratio indicated that DHA and EPA were not proportionally retained in muscular tissue, especially in fish fed the highest levels of DHA (D4). Similarly, in
T. ovatus [
42] and
E. cocoides [
18], the DHA/EPA ratio above 1.00 did not result in a greater deposition of DHA in the muscle. Thus, the results of this study suggest that, for better levels of DHA deposition in
E. marginatus fillets, the DHA/EPA ratio should be close to 1.00 and less than 3.00. Moreover, the physiological requirement for LC-PUFA is different from the necessity to maintain high levels of LC-PUFA in tissues [
47]. Thus, the dietary inclusion of DHA and EPA should be assessed in commercially relevant marine species aiming to make a better use of marine resources. Moreover, in marine fish, the oxidation of LC-PUFA is stimulated when the dietary levels of these substrates are in excess [
48].
In the liver, both fractions reflected the DHA present in the diet and, similar to muscle, PL had higher levels of LC-PUFA than TG, especially DHA. This result is consonant with the studies conducted so far that suggest that DHA in the polar lipids of fish tissues is highly conserved in the liver and muscle, among other tissues [
49,
50,
51]. Although the different dietary DHA/EPA ratios did not affect the sum of LC-PUFA in the liver, when fish were fed 0.5 of EPA and 1.0 of DHA (D3), the levels of DHA deposition were not different from those fish that received 1.5 of DHA (D4). Possibly, the balance in the proportion of these FAs in the metabolism is more important than the amount itself. Thus, increasing dietary DHA levels and depriving fish of EPA affect the FA profile in TG and PL. Studies with
T. ovatus have shown that DHA is important in the immune system and that lysozyme activity increases as dietary DHA increases. However, lysozyme activity decreased when dietary DHA levels were much higher than those of EPA (2.2) [
42]. Similar results were observed in large yellow croaker,
Larmichthys crocea; when dietary LC-PUFA n-3 increased from 1.37% to 2.25%, lysozyme activity decreased [
52]. DHA is important in numerous metabolic and physiological processes in animals, including marine fish, so receiving a proper content of DHA in the diet is imperative [
15,
19]. However, excessive levels of dietary DHA could reduce immune parameters such as lysozyme activity or even have adverse effects in fish tissue such as hepatic lesions, as described in European sea bass,
Dicentrarchus labrax [
53].
EPA levels in both hepatic fractions, TG and PL, reflected the diet, and decreased as the dietary DHA/EPA ratio increased. The reduction of EPA, especially in the PL composition, could be prejudicial to the immunological response of
E. marginatus juveniles, as EPA is a mediator of inflammatory processes, and, through phospholipase, A2 action is cleaved from PL to synthesize eicosanoids of the anti-inflammatory series [
54,
55]. In fish, the immunological response mediated by eicosanoids is partially related to the EPA and ARA levels, which are important in the regulation of the inflammatory response. The n-3/n-6 ratio in the feed is an index to consider for feed formulation, as imbalances can trigger adverse physiological consequences [
56,
57,
58].
The diet profile of the DHA/EPA ratio was less reflected in the organs of the nervous system analyzed in this study. In the eyes, the DHA content of the PL fraction was not affected by the diet, but when fish were fed with the highest DHA inclusion and maintained at the higher temperature (D4-HT), a remodeling of FA was observed, increasing MUFA (especially 18:1n-9) and PUFA (not a specific FA) levels and causing a decrease in SFA (especially 16:0). Environmental factors such as water temperature affect the metabolic rate of fish [
59], and, in response to fluctuations in water temperature and the metabolic rate, fish can restructure the phospholipid and FA composition of cell membranes toward increasing fluidity [
24]. In this context, MUFA and PUFA play an important role in membrane composition, as the presence of unsaturation in the membrane creates “bends” in the lipid layers that ultimately support membrane fluidity [
60]. Our results suggest that juvenile of
E. marginatus, due to higher metabolic demand at the higher temperature, deposited higher levels of MUFA and PUFA in the PL fraction of the eyes, since the increase in the temperature increases the metabolic rate of fish, and, this tissue, belonging to the nervous system, is very energy-consuming. However, this profile was not observed in the brain. Consonant with this pattern, in the TG fraction of the eyes, the levels of 18:0 decreased at a high temperature. The TG are molecules used by animals, including teleosts, as energy stores [
13], and SFA are preferentially oxidized due to their fast energy delivery [
61]. Thus, fish maintained at a high temperature possibly increase the metabolic rate and SFA oxidation to balance the energy demand.
In general, LC-PUFA levels were preserved in these tissues regardless of the temperature or the levels at which these FAs were included in the diet. This pattern reflects the importance of DHA and EPA in nervous tissues such as the brain and eyes of marine teleosts. The results demonstrated that, in
E. marginatus juveniles, DHA is more preserved than EPA, especially in the PL of these tissues, in which there are no differences in the levels of DHA content. This profile was expected, and the fact that the FA profile of PL is more conserved, especially in nervous tissues, regardless of the diet, has already been observed in fish [
62]. In the present study, it was also observed that, even when fed higher (D1) or equal (D2) levels of EPA than DHA, fish retained more DHA, suggesting that EPA is more oxidized, as was also observed in
R. canadum [
63]. Beyond metabolic alterations, our study also revealed morphological alterations in the liver, both related to temperature and the dietary DHA/EPA ratio. The liver is a key organ in fish metabolism [
13] and energy modulation is vital for fish at a low temperature [
64]. Cold stress can lead to considerable energy expenditure [
65] and this probably explains the reduced lipid deposition pattern and hepatic lipid droplet area of fish kept at a low temperature compared to fish kept at a high temperature. Moreover, the reduction in lipid droplet area was related to an increased dietary DHA/EPA ratio, a result that corroborates the FA profile, which showed a decrease in EPA content in liver levels of fish kept at a low temperature as the dietary DHA/EPA increased. On the other hand, the higher deposition of lipid droplets in fish kept at a high temperature could be related to the increase of lipid deposition in the liver of fish kept at this temperature; however, when maintained at the higher temperature, there is no difference in the lipid vacuole area of fish fed D3 and D4, suggesting that there is a limit for DHA deposition from dietary intake.
Some studies have been performed aiming to, through nutritional manipulation, increase DHA and EPA retention in the fillets of commercial marine fish species [
25,
66,
67] due to consumer pressure for a healthier product [
47], as these FAs promote benefits in human health [
68,
69]. Considering that: (1) aquaculture is the unique way to offer dusky grouper fillet to the market, considering the conservation state of this species [
2]; (2) the production of aquafeeds is one of the most expensive points in the productive cycle [
70]; and (3) alternative ingredients to FO such as algal oils are still difficult to scale and have a costly production [
47], studies that access the dietary ratio of DHA/EPA in marine fish nutrition are important in order to better understand the physiology of these species and further improve fillet quality.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}