


Increased Prevalence and New Evidence of Multi-Species Chelonid Herpesvirus 5 (ChHV5) Infection in the Sea Turtles of Mabul Island, Borneo

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Isolation and Polymerase Chain Reaction (PCR) of ChHV5 Regions
2.3. Sanger Sequencing and Phylogeography of the ChHV5 Region
2.4. Ethics Statement
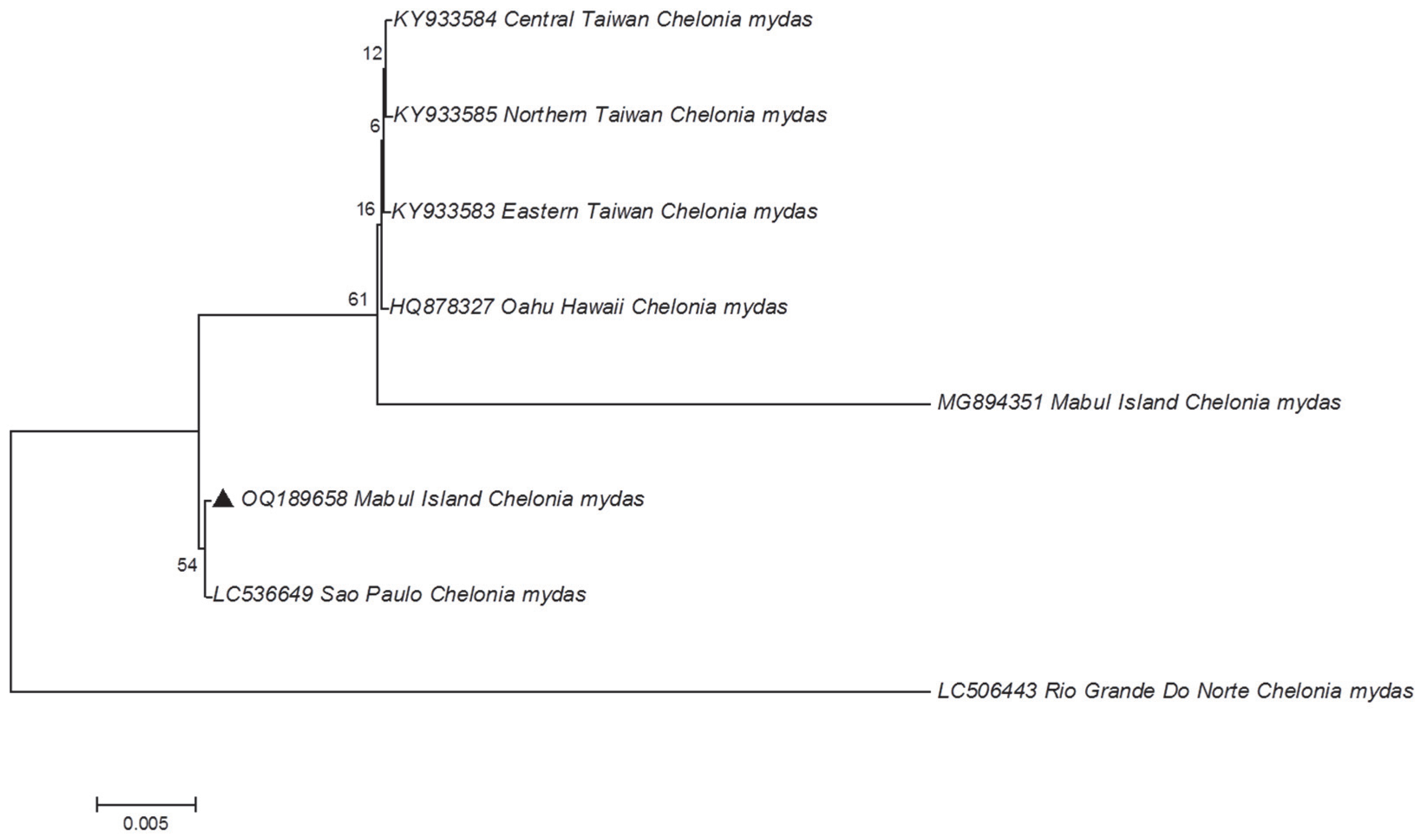
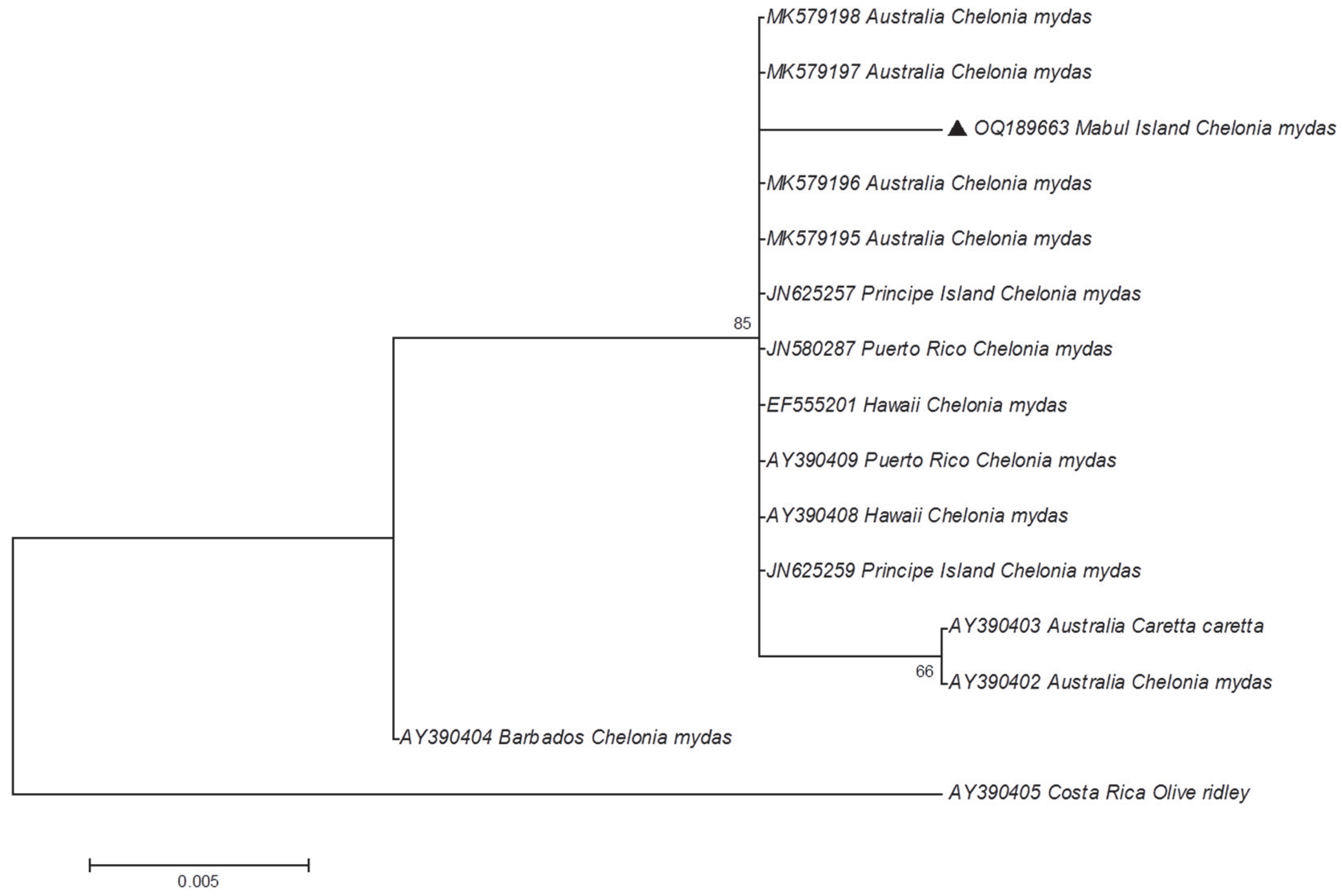
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, K.; Ariel, E.; Burgess, G.; Read, M. A Review of Fibropapillomatosis in Green Turtles (Chelonia mydas). Vet. J. 2016, 212, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Whitmore, L.; Yetsko, K.; Farrell, J.A.; Page-Karjian, A.; Daniel, W.; Shaver, D.J.; Duffy, D.J. Evolutionary Comparisons of Chelonid Alphaherpesvirus 5 (ChHV5) Genomes from Fibropapillomatosis-Afflicted Green (Chelonia mydas), Olive Ridley (Lepidochelys olivacea) and Kemp’s Ridley (Lepidochelys kempii) Sea Turtles. Animals 2021, 11, 2489. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.A.; Lutz, P.L. Marine turtles as sentinels of ecosystem health: Is fibropapillomatosis an indicator? EcoHealth 2004, 1, 275–283. [Google Scholar] [CrossRef]
- Li, T.H.; Hsu, W.L.; Lan, Y.C.; Balazs, G.H.; Work, T.M.; Tseng, C.T.; Chang, C.C. Identification of Chelonid herpesvirus 5 (ChHV5) in Endangered Green Turtles (Chelonia mydas) with Fibropapillomatosis in Asia. Bull. Mar. Sci. 2017, 93, 1011–1022. [Google Scholar] [CrossRef]
- Rodenbusch, C.R.; Baptistotte, C.; Werneck, M.R.; Pires, T.T.; Melo, M.T.D.; De Ataíde, M.W.; Canal, C.W. Fibropapillomatosis in green turtles Chelonia mydas in Brazil: Characteristics of tumors and virus. Dis. Aquat. Org. 2014, 111, 207–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejía-Radillo, R.Y.; Zavala-Norzagaray, A.A.; Chávez-Medina, J.A.; Aguirre, A.A.; Escobedo-Bonilla, C.M. Presence of Chelonid Herpesvirus 5 (ChHV5) in Sea Turtles in Northern Sinaloa, Mexico. Dis. Aquat. Org. 2019, 132, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, A.L.; Palaniappan, P.; Subbiah, V.K. First Evidence of Chelonid Herpesvirus 5 (ChHV5) Infection in Green Turtles (Chelonia mydas) from Sabah, Borneo. Pathogens 2021, 10, 1404. [Google Scholar] [CrossRef] [PubMed]
- Flint, M.; Limpus, C.J.; Patterson-Kane, J.C.; Murray, P.J.; Mills, P.C. Corneal Fibropapillomatosis in Green Sea Turtles (Chelonia mydas) in Australia. J. Comp. Pathog. 2010, 142, 341–346. [Google Scholar] [CrossRef]
- Page-Karjian, A.; Serrano, M.E.; Cartzendafner, J.; Morgan, A.; Ritchie, B.W.; Gregory, C.R.; Harms, C.A. Molecular assessment of chelonid alphaherpesvirus 5 infection in tumor-free green (Chelonia mydas) and loggerhead (Caretta caretta) sea turtles in North Carolina, USA, 2015–2019. Animals 2020, 10, 1964. [Google Scholar] [CrossRef]
- Hendrickson, J.R. The Green Sea Turtle, Chelonia mydas (Linn.) in Malaya and Sarawak. In Proceedings of the Zoological Society of London; Blackwell Publishing Ltd.: Oxford, UK, 1958; Volume 130, pp. 455–535. [Google Scholar] [CrossRef]
- Adnyana, W.; Ladds, P.W.; Blair, D. Observations of Fibropapillomatosis in Green Turtles (Chelonia mydas) in Indonesia. Aust. Vet. J. 1997, 75, 737–742. [Google Scholar] [CrossRef]
- Lim, K.K.; Syed Hussein, M.A.; Palaniappan, P. Abundance, placement and sexual identity of the epizoic barnacle Chelonibia testudinaria relative to the size and species of host turtles in Mabul Island, Malaysia. J. Mar. Biol. Assoc. UK 2020, 100, 1299–1309. [Google Scholar] [CrossRef]
- Bolten, A.B. Techniques for measuring sea turtles. Res. Man. Tech. Cons. Sea Turt. 1999, 4, 110–114. [Google Scholar]
- Stephens, S.H.; White, V.C.; Bremer, J.R.A. Assessment of genetic tissue sampling methods for the critically endangered Kemp’s Ridley Sea Turtle (Lepidochelys kempii). Mar. Turt. News 2010, 128, 19. [Google Scholar]
- Alfaro-Núñez, A.; Gilbert, M.T.P. Validation of a sensitive PCR assay for the detection of Chelonid Fibropapilloma-associated Herpesvirus in Latent Turtle Infections. J. Virol. Met. 2014, 206, 38–41. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. biol. and evol. 1987, 4, 406–425. [Google Scholar]
- Herbst, L.H. Fibropapillomatosis of Marine Turtles. Ann. Rev. Fish Dis. 1994, 4, 389–425. [Google Scholar] [CrossRef]
- Work, T.M.; Balazs, G.H.; Rameyer, R.A.; Morris, R.A. Retrospective Pathology Survey of Green Turtles Chelonia mydas with Fibropapillomatosis in the Hawaiian Islands, 1993–2003. Dis. Aquat. Org. 2004, 62, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Wynne, S.P. (Department of Fisheries and Marine Resources, Government of Anguilla, The Valley, Anguilla). Biometric relationships, size class structures, and growth rates of foraging Hawksbill (Eretmochelys imbricata) and Green (Chelonia mydas) Sea Turtles in Anguilla, with observations on occurrence and prevalence of fibropapilloma. Anguilla Fish. Mar. Resour. Res. Bull. 2016, 4, 1–9. [Google Scholar]
- Williams, E.H., Jr.; Bunkley-Williams, L.; Peters, E.C.; Pinto-Rodriguez, B.; Matos-Morales, R.; Mignucci-Giannoni, A.A.; Boulon, R.H. An epizootic of cutaneous fibropapillomas in green turtles Chelonia mydas of the Caribbean: Part of a panzootic? J. Aquat. Anim. Health 1994, 6, 70–78. [Google Scholar] [CrossRef]
- D’Amato, A.F.; Moraes-Neto, M. First documentation of fibropapillomas verified by histopathology in Eretmochelys imbricata. Mar. Turt. News 2000, 89, 12–13. [Google Scholar]
- Alfaro-Núñez, A.; Bojesen, A.M.; Bertelsen, M.F.; Wales, N.; Balazs, G.H.; Gilbert, M.T.P. Further Evidence of Chelonid Herpesvirus 5 (ChHV5) Latency: High Levels of ChHV5 DNA Detected in Clinically Healthy Marine Turtles. PeerJ 2016, 4, e2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frandsen, H.R.; Wilson, H.M.; Walker, S.; Purvin, C.M.; Dutton, P.; Lacasella, E.L.; SHAVER, D.J. First olive ridley sea turtle (Lepidochelys olivacea) stranding in Texas, USA and identification of chelonid alphaherpesvirus 5 (ChHV5) variant present in tumor tissue. Herpetol. Rev. 2021, 52, 492–499. [Google Scholar]
- Work, T.M.; Dagenais, J.; Weatherby, T.M.; Balazs, G.H.; Ackermann, M. In Vitro Replication of Chelonid Herpesvirus 5 in Organotypic Skin Cultures from Hawaiian Green Turtles (Chelonia mydas). J. Virol. 2017, 91, e00404–e00417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page-Karjian, A.; Torres, F.; Zhang, J.; Rivera, S.; Diez, C.; Moore, P.A.; Brown, C. Presence of Chelonid Fibropapilloma-associated Herpesvirus in Tumored and Non-tumored Green Turtles, as Detected by Polymerase Chain Reaction, in Endemic and Non-endemic Aggregations, Puerto Rico. SpringerPlus 2012, 1, 35. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, K.J.; Witherington, B.E.; Lohmann, C.M.; Salmon, M. Orientation, navigation, and natal beach homing in sea turtles. In The Biology of Sea Turtles; CRC Press: Boca Raton, FL, USA, 2017; pp. 108–135. [Google Scholar]
- Chaves, A.; Aguirre, A.A.; Blanco-Pena, K.; Moreira-Soto, A.; Monge, O.; Torres, A.M.; Jiménez, M. Examining the role of transmission of chelonid alphaherpesvirus 5. EcoHealth 2017, 14, 530–541. [Google Scholar] [CrossRef]
- Farrell, J.A.; Yetsko, K.; Whitmore, L.; Whilde, J.; Eastman, C.B.; Ramia, D.R.; Duffy, D.J. Environmental DNA monitoring of oncogenic viral shedding and genomic profiling of sea turtle fibropapillomatosis reveals unusual viral dynamics. Comm. Biol. 2021, 4, 1–17. [Google Scholar] [CrossRef]
- Jones, K.; Burgess, G.; Budd, A.M.; Huerlimann, R.; Mashkour, N.; Ariel, E. Molecular evidence for horizontal transmission of chelonid alphaherpesvirus 5 at green turtle (Chelonia mydas) foraging grounds in Queensland, Australia. PLoS ONE 2020, 15, e0227268. [Google Scholar] [CrossRef]
- Bolten, A.B. Variation in sea turtle life history patterns: Neritic vs. oceanic developmental stages. Biol. Sea Turt. 2003, 2, 243–257. [Google Scholar]
- Amorocho, D.; Reina, R. Feeding ecology of the East Pacific green sea turtle Chelonia mydas at Gorgona National Park, Colombia. End. Spec. Res. 2007, 3, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Greenblatt, R.J.; Work, T.M.; Balazs, G.H.; Sutton, C.A.; Casey, R.N.; Casey, J.W. The Ozobranchus Leech is a Candidate Mechanical Vector for the Fibropapilloma-associated Turtle Herpesvirus found Latently Infecting Skin Tumors on Hawaiian Green Turtles (Chelonia mydas). Virology 2004, 321, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Rodenbusch, C.R.; Marks, F.S.; Canal, C.W.; Reck, J. Marine leech Ozobranchus margoi parasitizing loggerhead turtle (Caretta caretta) in Rio Grande do Sul, Brazil. Rev. Bras. Par. Vet. 2012, 21, 301–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, R.A.; Christodoulides, N.; Jensen, I.M.; Becker, D.J.; Mansfield, K.L.; Savage, A.E. Gene expression changes with tumor disease and leech parasitism in the juvenile green sea turtle skin transcriptome. Gene 2021, 800, 145800. [Google Scholar] [CrossRef] [PubMed]
- Rittenburg, L.T.; Kelley, J.R.; Mansfield, K.L.; Savage, A.E. Marine leech parasitism of sea turtles varies across host species, seasons, and the tumor disease fibropapillomatosis. Dis. Aquat. Org. 2021, 143, 1–12. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Primer Name | Primer Sequences 5′–3′ | Size (bp) |
|---|---|---|---|
| Capsid protein gene | UL18 | F: GTGGAACCCCGCCGGGTAAT R: TGATCCGGGCCGAGTAGCGC | 140 |
| Glycoprotein H gene | UL22 | F: AACGCCCTTTCCTCCGACCCATATT R: GCTGGGGGAGCATCGTGCAAA | 179 |
| Glycoprotein B gene | UL27 | F: CTAGATACATACTGGCCRTGCTCGTC R: GCCAGCGACCATCCGGAG | 143 |
| Turtle Species | No. of Samples | Turtles with FP Tumor (PCR-Positive) | Turtles without FP Tumor (PCR-Positive) |
|---|---|---|---|
| Green | 63 | 1 | 26 |
| Hawksbill | 5 | 0 | 3 |
| Olive Ridley | 1 | 0 | 1 |
| TOTAL | 31 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robben, D.M.; Palaniappan, P.; Loganathan, A.L.; Subbiah, V.K. Increased Prevalence and New Evidence of Multi-Species Chelonid Herpesvirus 5 (ChHV5) Infection in the Sea Turtles of Mabul Island, Borneo. Animals 2023, 13, 290. https://doi.org/10.3390/ani13020290
Robben DM, Palaniappan P, Loganathan AL, Subbiah VK. Increased Prevalence and New Evidence of Multi-Species Chelonid Herpesvirus 5 (ChHV5) Infection in the Sea Turtles of Mabul Island, Borneo. Animals. 2023; 13(2):290. https://doi.org/10.3390/ani13020290
Chicago/Turabian StyleRobben, Dexter Miller, Pushpa Palaniappan, Aswini Leela Loganathan, and Vijay Kumar Subbiah. 2023. "Increased Prevalence and New Evidence of Multi-Species Chelonid Herpesvirus 5 (ChHV5) Infection in the Sea Turtles of Mabul Island, Borneo" Animals 13, no. 2: 290. https://doi.org/10.3390/ani13020290