
A Window of Vulnerability: Chronic Environmental Stress Does Not Impair Reproduction in the Swordfish Xiphias gladius
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Transcriptomic Analysis
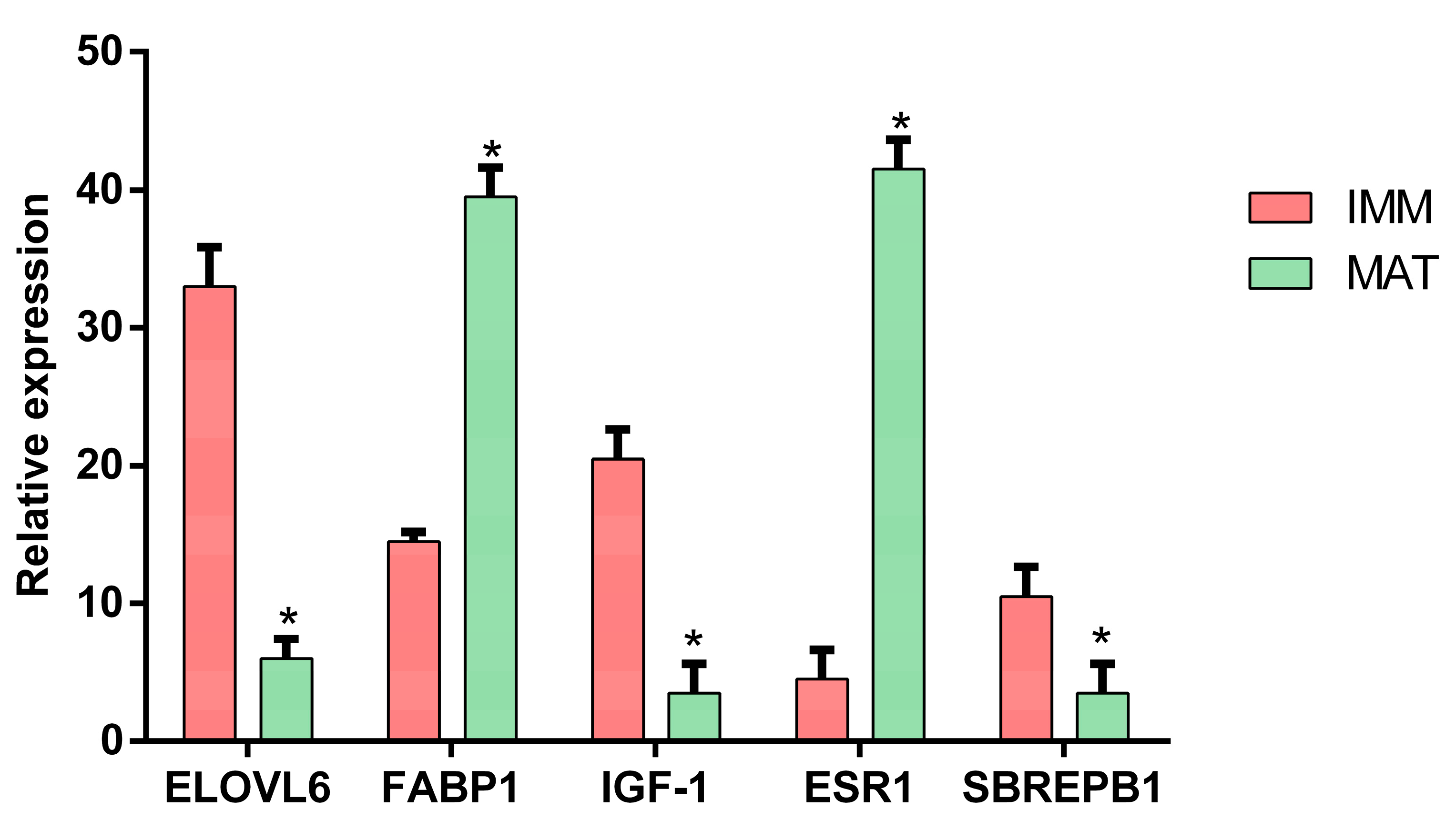
2.3. Experimental Validation
2.4. Histological Analysis
3. Results
3.1. Transcriptomic Analysis
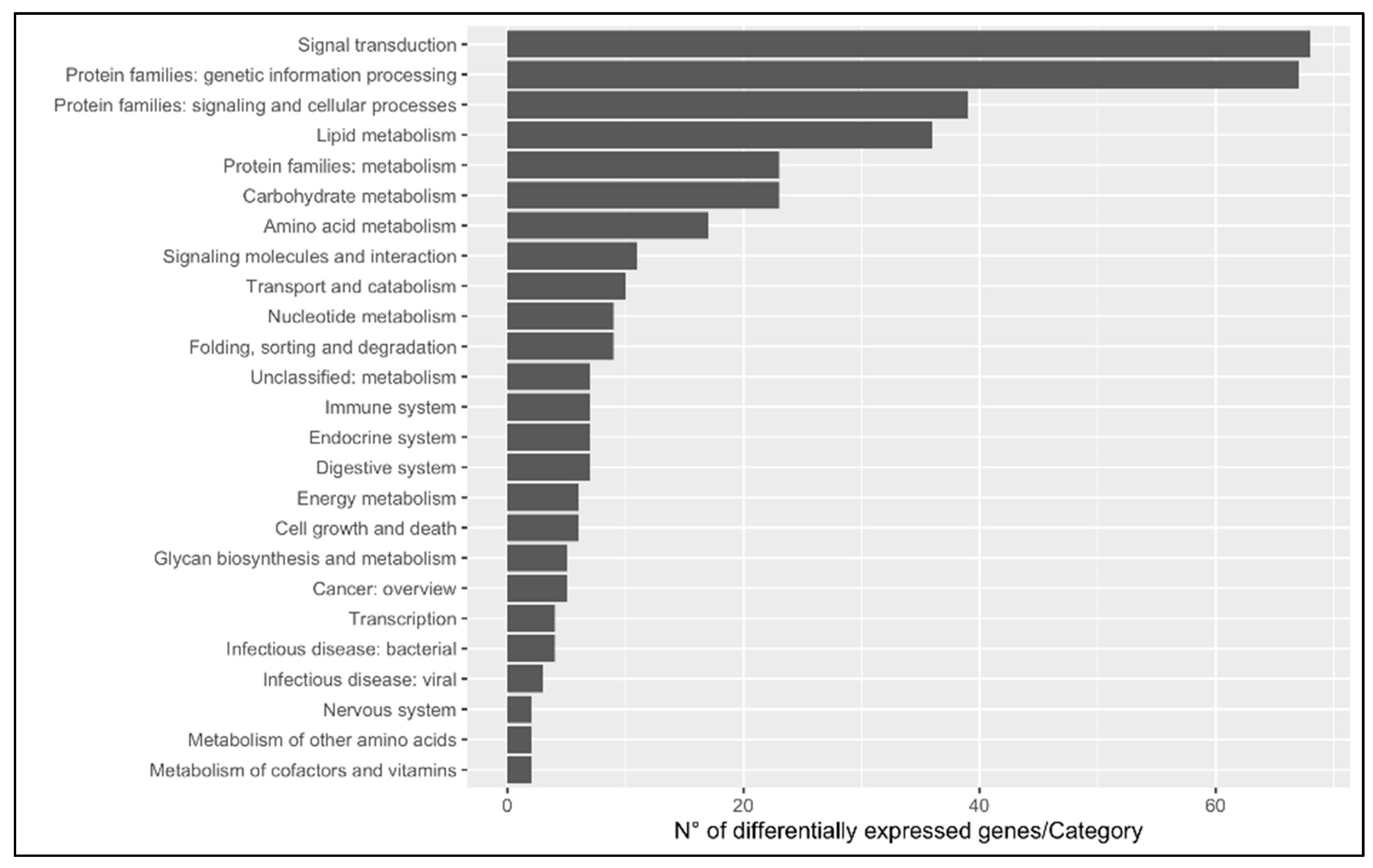
3.1.1. DEGs
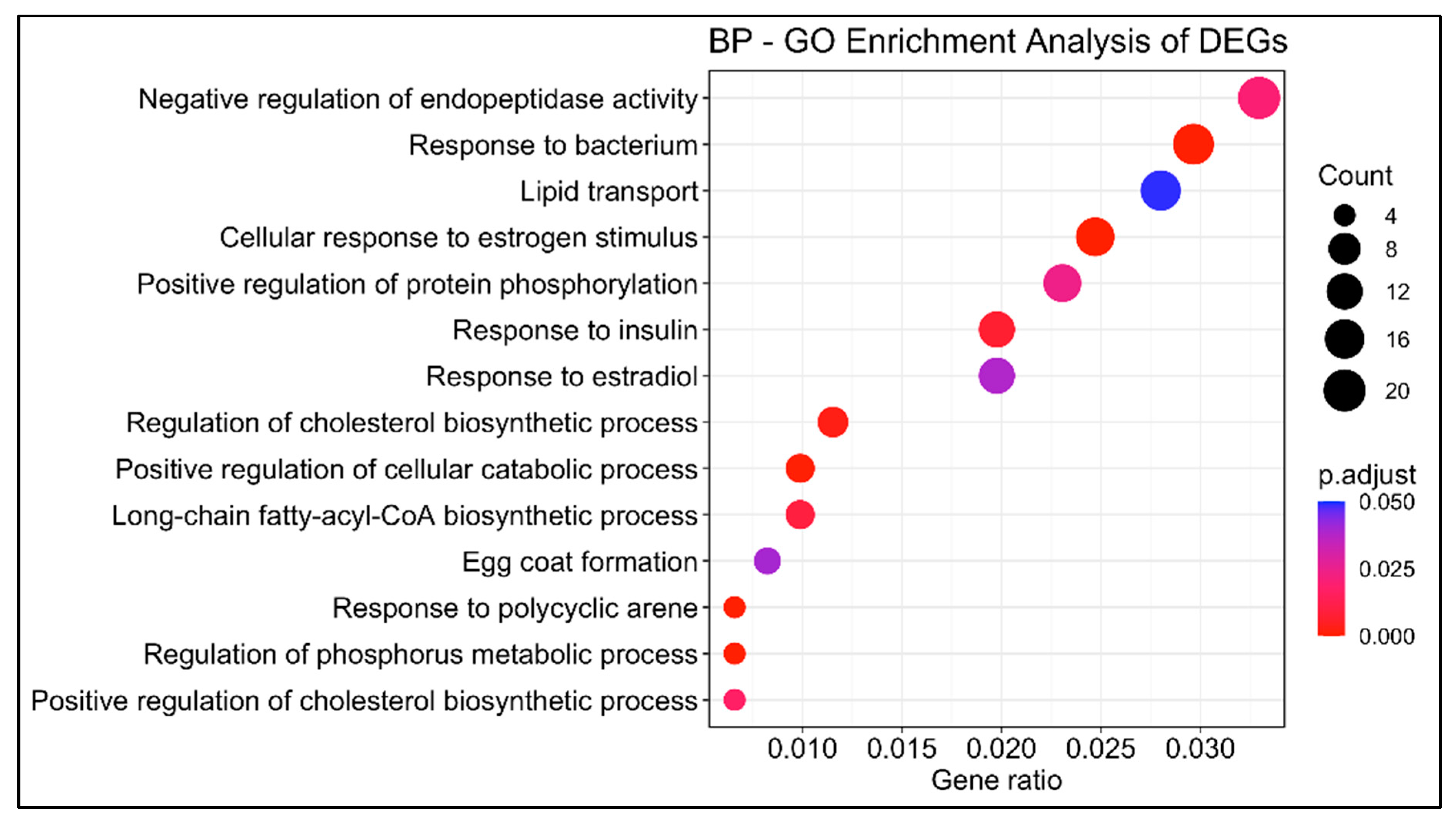
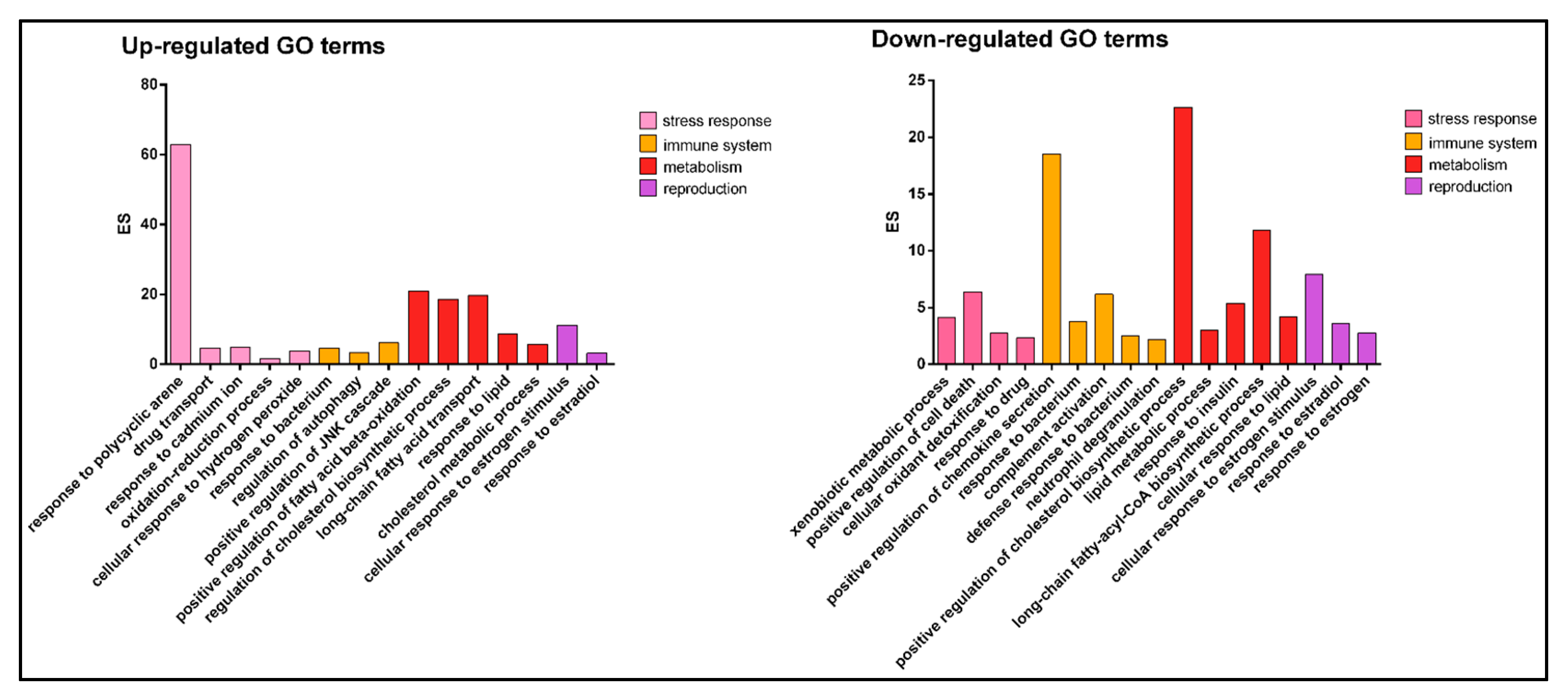
3.1.2. Gene Ontology
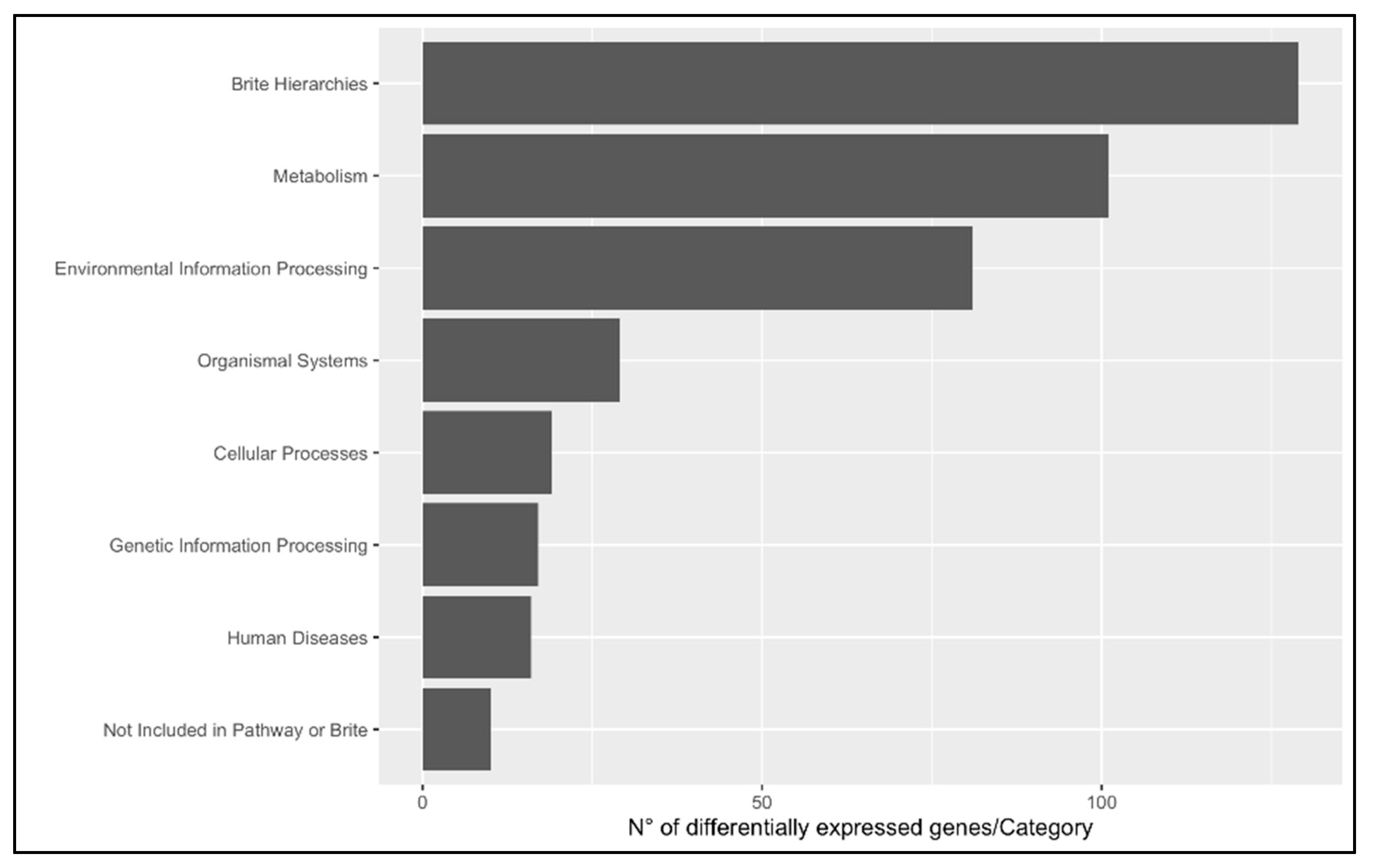
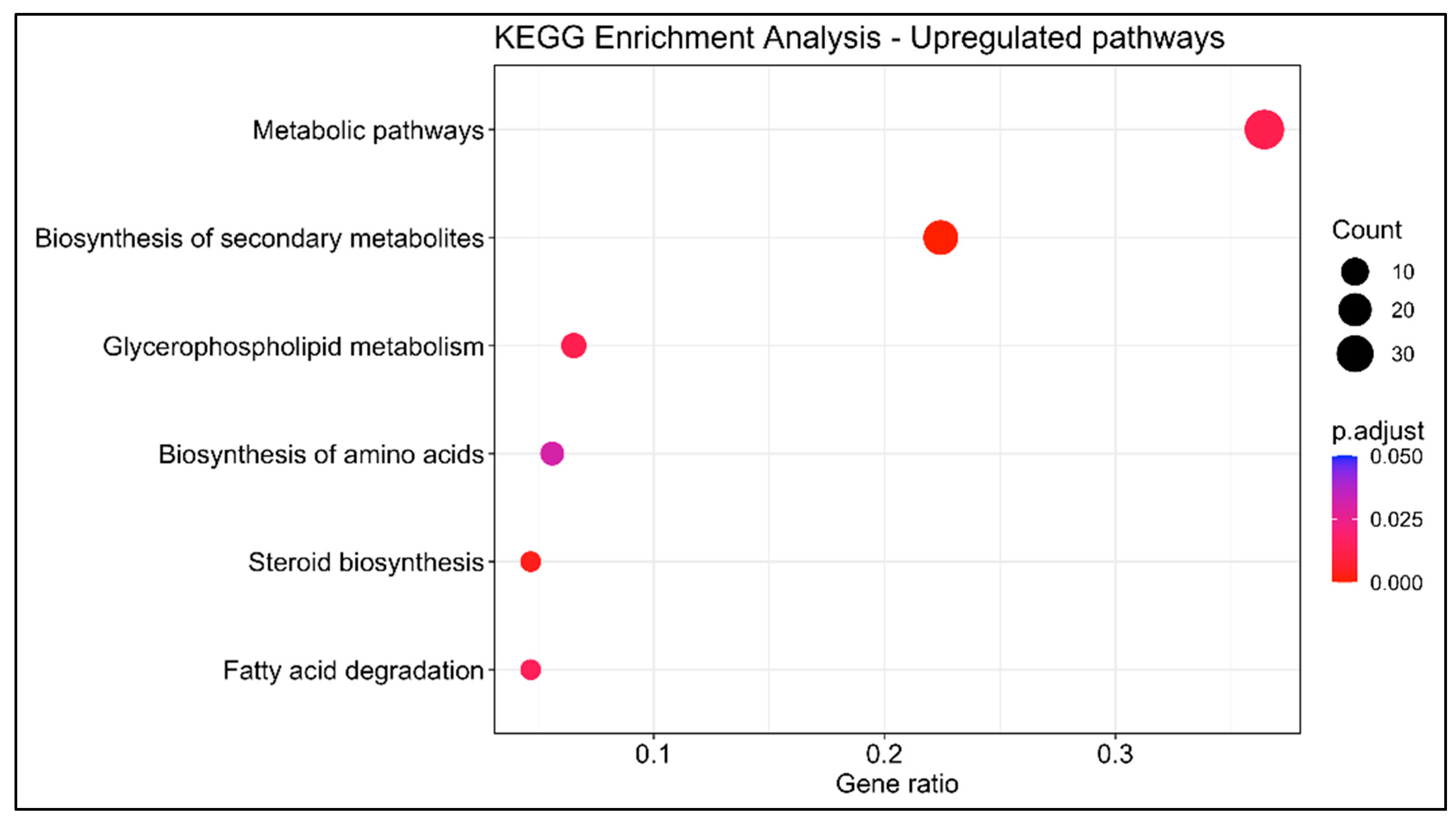
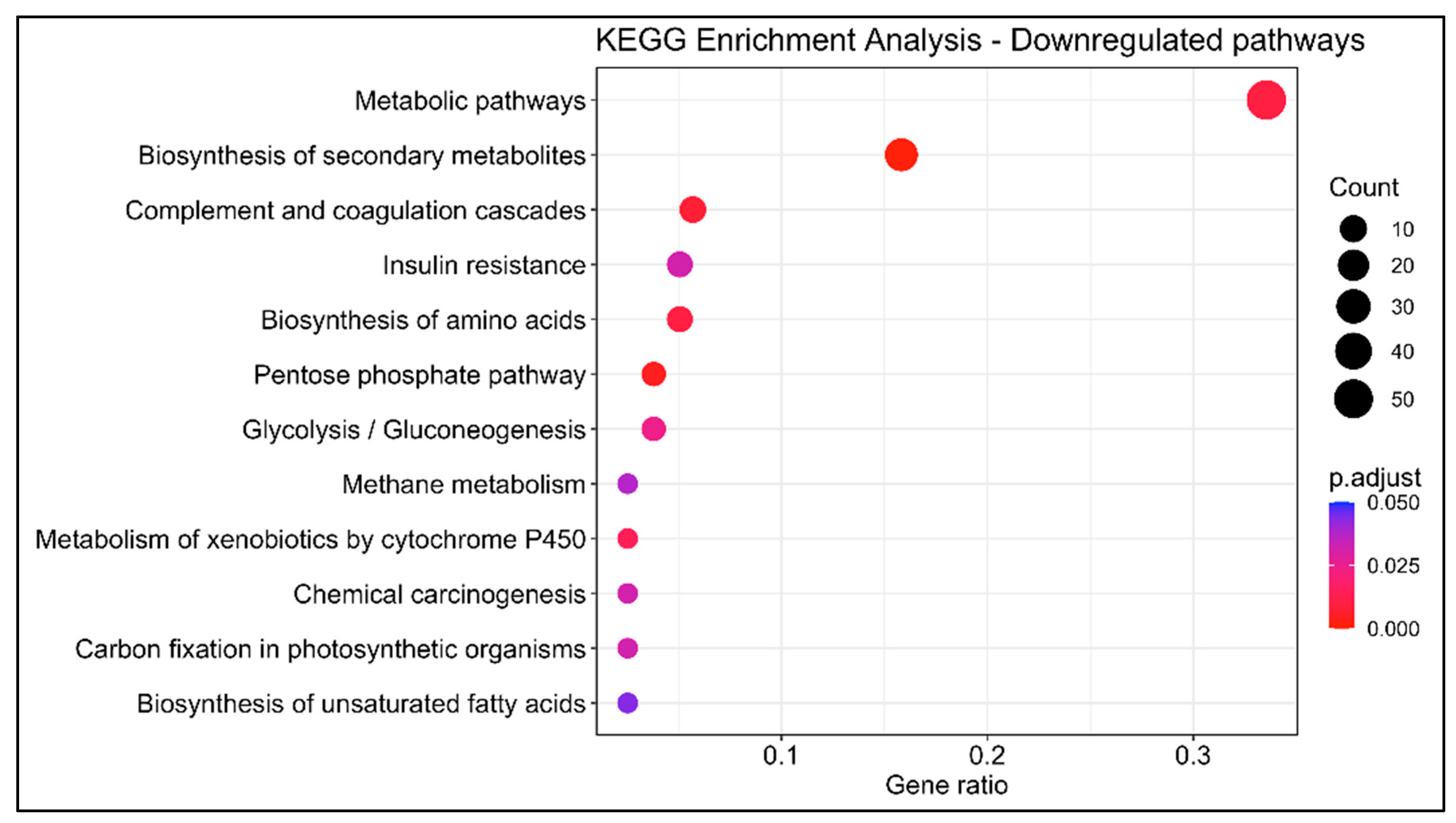
3.1.3. KEGG Enrichment Analysis
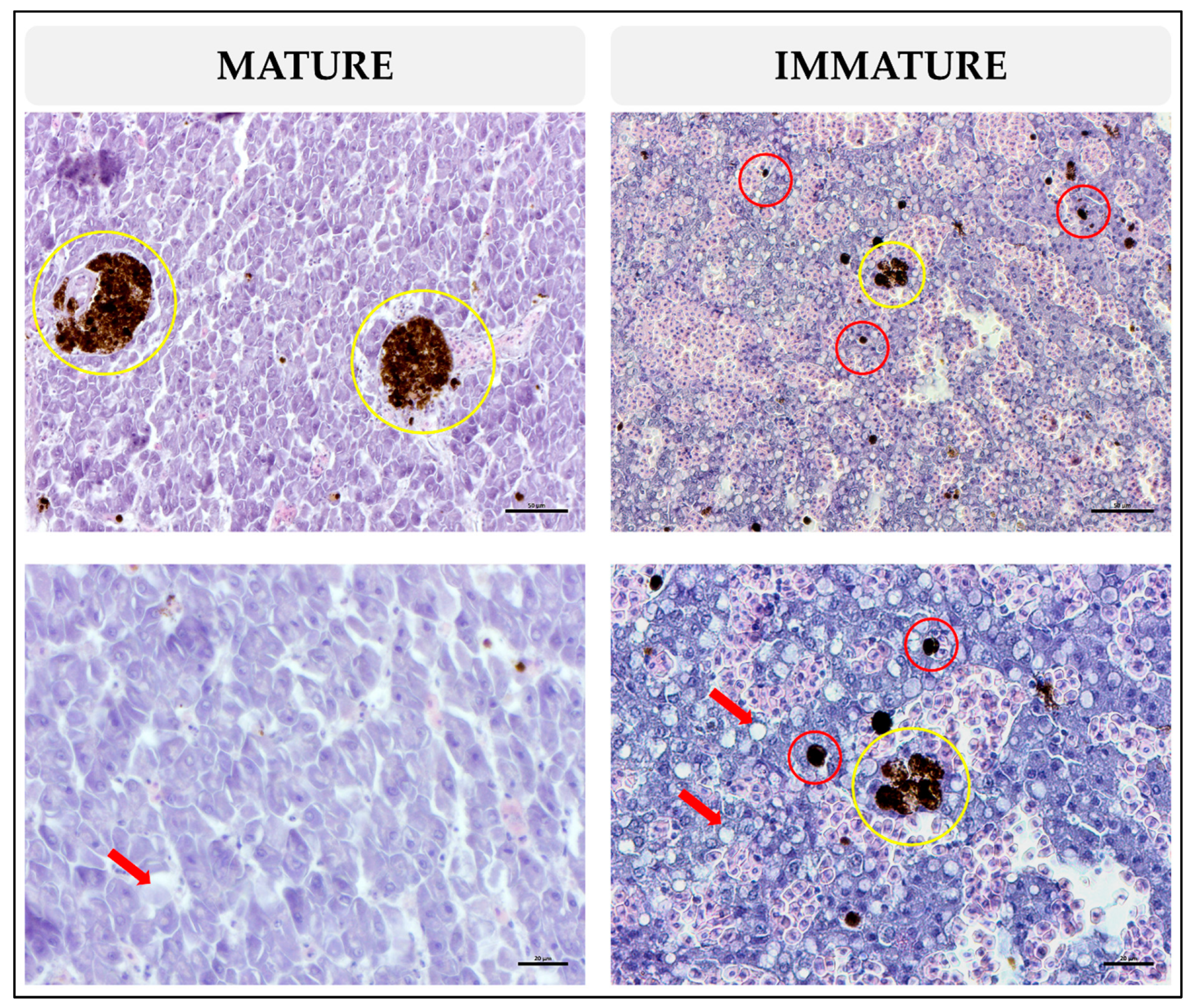
3.2. Histological Analysis
3.2.1. MMs and MMCs Related to Fish Size
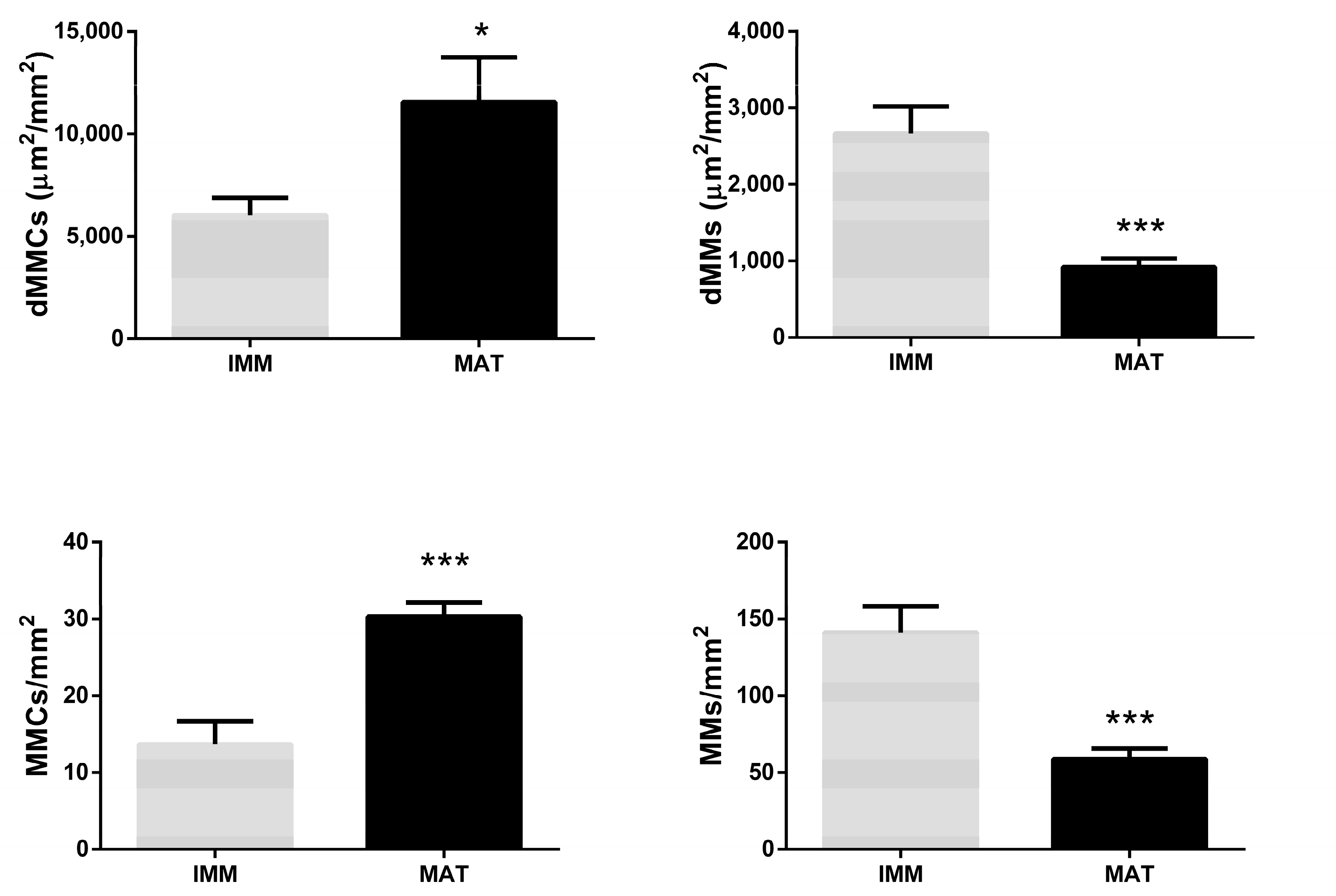
3.2.2. MMs and MMCs Related to Fish Sexual Maturity
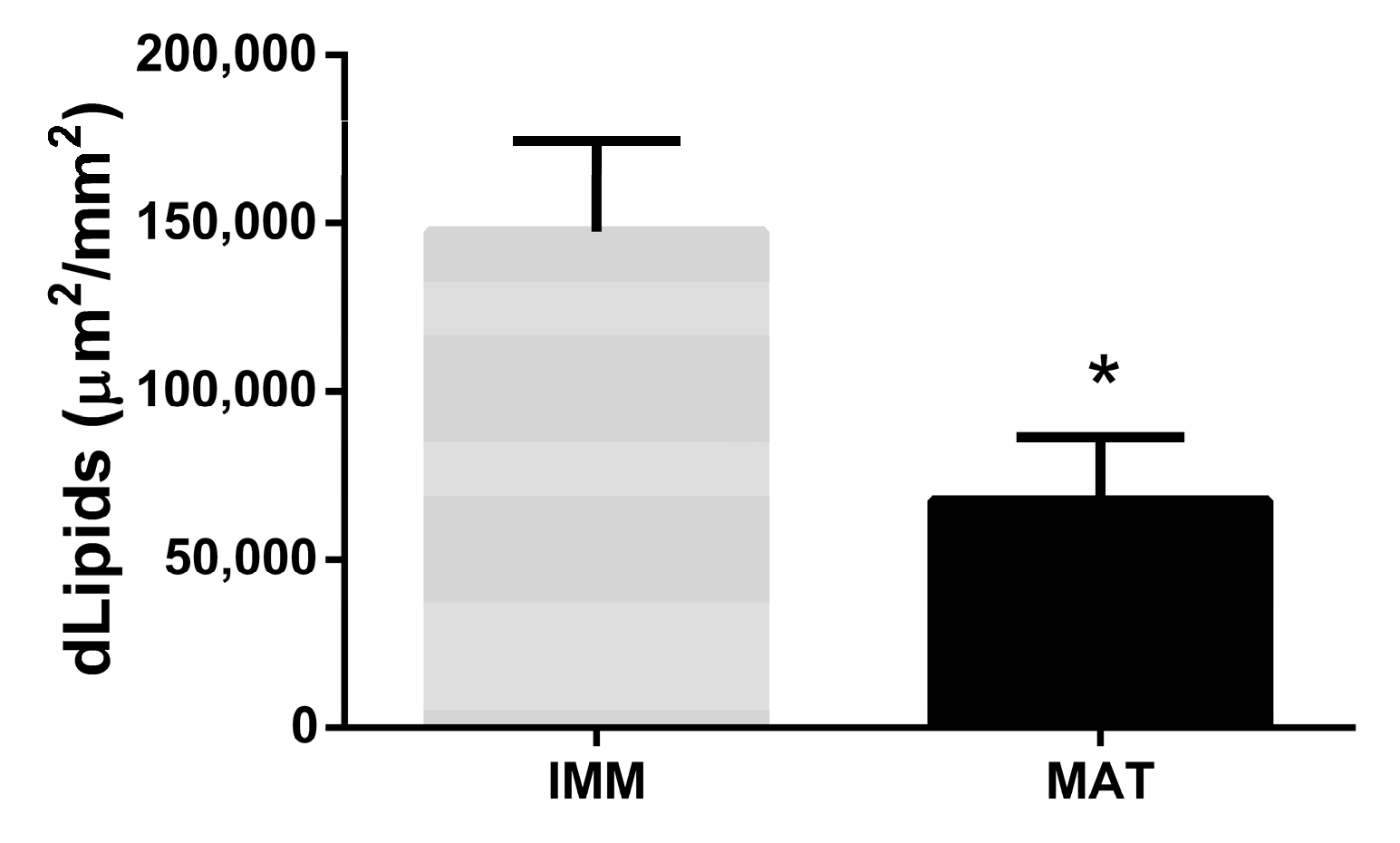
3.2.3. Lipids Related to Fish Size and Sexual Maturity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICCAT Report of the 2016 Mediterranean Swordfish Stock Assessment Meeting. Collect. Vol. Sci. Pap. ICCAT 2016, 73, 1005–1096.
- Gomez Vives, M.J.; Macías, D.; Hattour, A.; de La Serna, J.M.; Gómez-Vives, M.J.; Godoy, D. Reproductive characteristics of swordfish (Xiphias gladius) caught in the southwestern Mediterranean during 2003. Sci. Pap. ICCAT 2005, 58, 454–469. [Google Scholar]
- Marisaldi, L.; Basili, D.; Candelma, M.; Sesani, V.; Pignalosa, P.; Gioacchini, G.; Carnevali, O. Maturity Assignment Based on Histology-Validated Macroscopic Criteria: Tackling the Stock Decline of the Mediterranean Swordfish (Xiphias Gladius). Aquat. Conserv. 2020, 30, 303–314. [Google Scholar] [CrossRef]
- Gioacchini, G.; Marisaldi, L.; Basili, D.; Candelma, M.; Pignalosa, P.; Aiese Cigliano, R.; Sanseverino, W.; Hardiman, G.; Carnevali, O. A de Novo Transcriptome Assembly Approach Elucidates the Dynamics of Ovarian Maturation in the Swordfish (Xiphias Gladius). Sci. Rep. 2019, 9, 7375. [Google Scholar] [CrossRef] [Green Version]
- Arocha, F. Oocyte Development and Maturity Classification of Swordfish from the North-Western Atlantic. J. Fish Biol. 2002, 60, 13–27. [Google Scholar] [CrossRef]
- Abid, N.; Laglaoui, A.; Arakrak, A.; Bakkali, M. The Reproductive Biology of Swordfish (Xiphias Gladius) in the Strait of Gibraltar. J. Mar. Biol. Assoc. United Kingd. 2019, 99, 649–659. [Google Scholar] [CrossRef]
- Machado, A.M.; Muñoz-Merida, A.; Fonseca, E.; Veríssimo, A.; Pinto, R.; Felício, M.; da Fonseca, R.R.; Froufe, E.; Castro, L.F.C. Liver Transcriptome Resources of Four Commercially Exploited Teleost Species. Sci. Data 2020, 7, 214. [Google Scholar] [CrossRef]
- Valenzuela-Quiñonez, F. How Fisheries Management Can Benefit from Genomics? Brief. Funct. Genom. 2016, 15, 352–357. [Google Scholar] [CrossRef] [Green Version]
- Basili, D.; Gioacchini, G.; Todisco, V.; Candelma, M.; Marisaldi, L.; Pappalardo, L.; Carnevali, O. Opsins and Gonadal Circadian Rhythm in the Swordfish (Xiphias gladius) Ovary: Their Potential Roles in Puberty and Reproductive Seasonality. Gen. Comp. Endocrinol. 2021, 303, 113707. [Google Scholar] [CrossRef]
- Harshman, L.G.; Zera, A.J. The Cost of Reproduction: The Devil in the Details. Trends Ecol. Evol. 2007, 22, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Lochmiller, R.L.; Deerenberg, C. Trade-Offs in Evolutionary Immunology: Just What Is the Cost of Immunity? Oikos 2000, 88, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.C.; Moore, C.J.; Saal, F.S.V.; Swan, S.H. Plastics, the Environment and Human Health: Current Consensus and Future Trends. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2153. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Landscape Modification and Habitat Fragmentation: A Synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Radchuk, V.; Reed, T.; Teplitsky, C.; van de Pol, M.; Charmantier, A.; Hassall, C.; Adamík, P.; Adriaensen, F.; Ahola, M.P.; Arcese, P.; et al. Adaptive Responses of Animals to Climate Change Are Most Likely Insufficient. Nat. Commun. 2019, 10, 3109. [Google Scholar] [CrossRef] [Green Version]
- Gernhöfer, M.; Pawert, M.; Schramm, M.; Müller, E.; Triebskorn, R. Ultrastructural Biomarkers as Tools to Characterize the Health Status of Fish in Contaminated Streams. J. Aquat. Ecosyst. Stress Recovery 2001, 8, 241–260. [Google Scholar] [CrossRef]
- Martin, S.A.M.; Douglas, A.; Houlihan, D.F.; Secombes, C.J. Starvation Alters the Liver Transcriptome of the Innate Immune Response in Atlantic Salmon (Salmo Salar). BMC Genom. 2010, 11, 418. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Wen, X.; Hu, Y.; Zhang, X.; Wang, D.; Yin, S. Copper Nanoparticles Induced Oxidation Stress, Cell Apoptosis and Immune Response in the Liver of Juvenile Takifugu fasciatus. Fish Shellfish Immunol. 2019, 84, 648–655. [Google Scholar] [CrossRef]
- Shoieb, S.M.; El-Ghiaty, M.A.; Alqahtani, M.A.; El-Kadi, A.O.S. Cytochrome P450-Derived Eicosanoids and Inflammation in Liver Diseases. Prostaglandins Other Lipid Mediat. 2019, 147, 106400. [Google Scholar] [CrossRef]
- Ayisi, C.L.; Yamei, C.; Zhao, J.L. Genes, Transcription Factors and Enzymes Involved in Lipid Metabolism in Fin Fish. Agri Gene 2018, 7, 7–14. [Google Scholar] [CrossRef]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.K. Fish Oil Replacement in Finfish Nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Bemanian, V.; Male, R.; Goksøyr, A. The Aryl Hydrocarbon Receptor-Mediated Disruption of Vitellogenin Synthesis in the Fish Liver: Cross-Talk between AHR- and ERα -Signalling Pathways. Comp. Hepatol. 2004, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Leaños-Castañeda, O.; van der Kraak, G. Functional Characterization of Estrogen Receptor Subtypes, ERα and ERβ, Mediating Vitellogenin Production in the Liver of Rainbow Trout. Toxicol. Appl. Pharmacol. 2007, 224, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Araújo, B.C.; Wade, N.M.; de Mello, P.H.; Rodrigues-Filho, J.; Garcia, C.E.O.; de Campos, M.F.; Botwright, N.A.; Hashimoto, D.T.; Moreira, R.G. Characterization of Lipid Metabolism Genes and the Influence of Fatty Acid Supplementation in the Hepatic Lipid Metabolism of Dusky Grouper (Epinephelus marginatus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2018, 219–220, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hamid, N.; Junaid, M.; Wang, Y.; Pu, S.Y.; Jia, P.P.; Pei, D.S. Chronic Exposure to PPCPs Mixture at Environmentally Relevant Concentrations (ERCs) Altered Carbohydrate and Lipid Metabolism through Gut and Liver Toxicity in Zebrafish. Environ. Pollut. 2021, 273, 116494. [Google Scholar] [CrossRef]
- Lai, W.; Xu, D.; Li, J.; Wang, Z.; Ding, Y.; Wang, X.; Li, X.; Xu, N.; Mai, K.; Ai, Q. Dietary Polystyrene Nanoplastics Exposure Alters Liver Lipid Metabolism and Muscle Nutritional Quality in Carnivorous Marine Fish Large Yellow Croaker (Larimichthys Crocea). J. Hazard Mater. 2021, 419, 126454. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Bi, Q.; Pribytkova, E.; Wei, Y.; Sun, B.; Jia, L.; Liang, M. Different Lipid Scenarios in Three Lean Marine Teleosts Having Different Lipid Storage Patterns. Aquaculture 2021, 536, 736448. [Google Scholar] [CrossRef]
- Hiraoka, Y.; Okochi, Y.; Ohshimo, S.; Shimose, T.; Ashida, H.; Sato, T.; Ando, Y. Lipid and Fatty Acid Dynamics by Maternal Pacific Bluefin Tuna. PLoS ONE 2019, 14, e0222824. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and Functions of Lipids and Fatty Acids in Teleost Fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Dang, M.; Nowell, C.; Nguyen, T.; Bach, L.; Sonne, C.; Nørregaard, R.; Stride, M.; Nowak, B. Characterisation and 3D Structure of Melanomacrophage Centers in Shorthorn Sculpins (Myoxocephalus Scorpius). Tissue Cell 2019, 57, 34–41. [Google Scholar] [CrossRef]
- Fishelson, L. Cytomorphological Alterations of the Thymus, Spleen, Head-Kidney, and Liver in Cardinal Fish (Apogonidae, Teleostei) as Bioindicators of Stress. J. Morphol. 2006, 267, 57–69. [Google Scholar] [CrossRef]
- Haaparanta, A.; Valtonen, E.T.; Hoffmann, R.; Holmes, J. Do Macrophage Centres in Freshwater Fishes Reflect the Differences in Water Quality? Aquat. Toxicol. 1996, 34, 253–272. [Google Scholar] [CrossRef]
- Mela, M.; Randi, M.A.F.; Ventura, D.F.; Carvalho, C.E.V.; Pelletier, E.; Oliveira Ribeiro, C.A. Effects of Dietary Methylmercury on Liver and Kidney Histology in the Neotropical Fish Hoplias malabaricus. Ecotoxicol. Environ. Saf. 2007, 68, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Vigliano, F.A.; Bermúdez, R.; Quiroga, M.I.; Nieto, J.M. Evidence for Melano-Macrophage Centres of Teleost as Evolutionary Precursors of Germinal Centres of Higher Vertebrates: An Immunohistochemical Study. Fish Shellfish Immunol. 2006, 21, 467–471. [Google Scholar] [CrossRef]
- Nowak, B.F.; Dang, M.; Webber, C.; Neumann, L.; Bridle, A.; Bermudez, R.; Evans, D. Changes in the Splenic Melanomacrophage Centre Surface Area in Southern Bluefin Tuna (Thunnus maccoyii) Are Associated with Blood Fluke Infections. Pathogens 2021, 10, 79. [Google Scholar] [CrossRef]
- Widdicombe, M.; Power, C.; van Gelderen, R.; Nowak, B.F.; Bott, N.J. Relationship between Southern Bluefin Tuna, Thunnus maccoyii, Melanomacrophage Centres and Cardicola spp. (Trematoda: Aporocotylidae) Infection. Fish Shellfish Immunol. 2020, 106, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Qualhato, G.; de Sabóia-Morais, S.M.T.; Silva, L.D.; Rocha, T.L. Melanomacrophage Response and Hepatic Histopathologic Biomarkers in the Guppy Poecilia reticulata Exposed to Iron Oxide (Maghemite) Nanoparticles. Aquat. Toxicol. 2018, 198, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Stosik, M.P.; Tokarz-Deptuła, B.; Deptuła, W. Melanomacrophages and Melanomacrophage Centres in Osteichthyes. Cent. Eur. J. Immunol. 2019, 44, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Ardeshir, R.A.; Rastgar, S.; Salati, A.P.; Zabihi, E.; Movahedinia, A.; Feizi, F. The Effect of Nonylphenol Exposure on the Stimulation of Melanomacrophage Centers, Estrogen and Testosterone Level, and ERα Gene Expression in Goldfish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 254, 109270. [Google Scholar] [CrossRef]
- Basilone, G.; Gargano, A.; Corriero, A.; Zupa, R.; Santamaria, N.; Mangano, S.; Ferreri, R.; Pulizzi, M.; Mazzola, S.; Bonanno, A.; et al. Liver Melanomacrophage Centres and CYP1A Expression as Response Biomarkers to Environmental Pollution in European Anchovy (Engraulis encrasicolus) from the Western Mediterranean Sea. Mar. Pollut. Bull. 2018, 131, 197–204. [Google Scholar] [CrossRef]
- Passantino, L.; Santamaria, N.; Zupa, R.; Pousis, C.; Garofalo, R.; Cianciotta, A.; Jirillo, E.; Acone, F.; Corriero, A. Liver Melanomacrophage Centres as Indicators of Atlantic Bluefin Tuna, Thunnus thynnus L. Well-Being. J. Fish Dis. 2014, 37, 241–250. [Google Scholar] [CrossRef]
- Mai, Y.; Peng, S.; Li, H.; Lai, Z. Histological, Biochemical and Transcriptomic Analyses Reveal Liver Damage in Zebrafish (Danio rerio) Exposed to Phenanthrene. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 225, 108582. [Google Scholar] [CrossRef] [PubMed]
- Limonta, G.; Mancia, A.; Benkhalqui, A.; Bertolucci, C.; Abelli, L.; Fossi, M.C.; Panti, C. Microplastics Induce Transcriptional Changes, Immune Response and Behavioral Alterations in Adult Zebrafish. Sci. Rep. 2019, 9, 15775. [Google Scholar] [CrossRef] [PubMed]
- Hook, S.E.; Mondon, J.; Revill, A.T.; Greenfield, P.A.; Smith, R.A.; Turner, R.D.R.; Corbett, P.A.; Warne, M.S.J. Transcriptomic, Lipid, and Histological Profiles Suggest Changes in Health in Fish from a Pesticide Hot Spot. Mar. Environ. Res. 2018, 140, 299–321. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. Omics 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Gioacchini, G.; Rossi, G.; Carnevali, O. Host-Probiotic Interaction: New Insight into the Role of the Endocannabinoid System by in Vivo and Ex Vivo Approaches OPEN. Sci. Rep. 2017, 7, 1261. [Google Scholar] [CrossRef] [Green Version]
- Imai, M.; Mizoguchi, T.; Wang, M.; Li, Y.; Hasegawa, Y.; Tonoki, A.; Itoh, M. The Guppy (Poecilia reticulata) Is a Useful Model for Analyzing Age-Dependent Changes in Metabolism, Motor Function, and Gene Expression. Exp. Gerontol. 2022, 160, 111708. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.J.; Jiang, X.; Ahn, H.R.; Choi, J.; Kim, S.R.; Jung, K.H. Systematic Analysis of Cold Stress Response and Diurnal Rhythm Using Transcriptome Data in Rice Reveals the Molecular Networks Related to Various Biological Processes. Int. J. Mol. Sci. 2020, 21, 6872. [Google Scholar] [CrossRef]
- Zudaire, I.; Murua, H.; Grande, M.; Pernet, F.; Bodin, N. Accumulation and Mobilization of Lipids in Relation to Reproduction of Yellowfin Tuna (Thunnus albacares) in the Western Indian Ocean. Fish. Res. 2014, 160, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, M.D. Composition, Accumulation and Utilization of Yolk Lipids in Teleost Fish. Rev. Fish Biol. Fish. 1996, 6, 259–286. [Google Scholar] [CrossRef]
- Casanova-Nakayama, A.; Wenger, M.; Burki, R.; Eppler, E.; Krasnov, A.; Segner, H. Endocrine Disrupting Compounds: Can They Target the Immune System of Fish? Mar. Pollut. Bull. 2011, 63, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Seemann, F.; Knigge, T.; Duflot, A.; Marie, S.; Olivier, S.; Minier, C.; Monsinjon, T. Sensitive Periods for 17β-Estradiol Exposure during Immune System Development in Sea Bass Head Kidney. J. Appl. Toxicol. 2015, 36, 815–826. [Google Scholar] [CrossRef]
- Burgos-Aceves, M.A.; Cohen, A.; Smith, Y.; Faggio, C. Estrogen Regulation of Gene Expression in the Teleost Fish Immune System. Fish Shellfish Immunol. 2016, 58, 42–49. [Google Scholar] [CrossRef]
- Shivers, K.Y.; Amador, N.; Abrams, L.; Hunter, D.; Jenab, S.; Quiñones-Jenab, V. Estrogen Alters Baseline and Inflammatory-Induced Cytokine Levels Independent from Hypothalamic–Pituitary–Adrenal Axis Activity. Cytokine 2015, 72, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsolini, S.; Guerranti, C.; Perra, G.; Focardi, S. Polybrominated Diphenyl Ethers, Perfluorinated Compounds and Chlorinated Pesticides in Swordfish (Xiphias gladius) from the Mediterranean Sea. Environ. Sci. Technol. 2008, 42, 4344–4349. [Google Scholar] [CrossRef]
- Storelli, M.M.; Marcotrigiano, G.O. Occurrence and Accumulation of Organochlorine Contaminants in Swordfish from Mediterranean Sea: A Case Study. Chemosphere 2006, 62, 375–380. [Google Scholar] [CrossRef]
- Mortensen, A.S.; Arukwe, A. Interactions between Estrogen- and Ah-Receptor Signalling Pathways in Primary Culture of Salmon Hepatocytes Exposed to Nonylphenol and 3,3′,4,4′-Tetrachlorobiphenyl (Congener 77). Comp. Hepatol. 2007, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Gräns, J.; Wassmur, B.; Celander, M.C. One-Way Inhibiting Cross-Talk between Arylhydrocarbon Receptor (AhR) and Estrogen Receptor (ER) Signaling in Primary Cultures of Rainbow Trout Hepatocytes. Aquat. Toxicol. 2010, 100, 263–270. [Google Scholar] [CrossRef]
- Matthews, J.; Gustafsson, J.-Å. Estrogen Receptor and Aryl Hydrocarbon Receptor Signaling Pathways. Nucl. Recept. Signal. 2006, 4, nrs.04016. [Google Scholar] [CrossRef]
- Rüegg, J.; Swedenborg, E.; Wahlström, D.; Escande, A.; Balaguer, P.; Pettersson, K.; Pongratz, I. The Transcription Factor Aryl Hydrocarbon Receptor Nuclear Translocator Functions as an Estrogen Receptor β-Selective Coactivator, and Its Recruitment to Alternative Pathways Mediates Antiestrogenic Effects of Dioxin. Mol. Endocrinol. 2008, 22, 304–316. [Google Scholar] [CrossRef] [Green Version]
- Swedenborg, E.; Pongratz, I. AhR and ARNT Modulate ER Signaling. Toxicology 2010, 268, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Safe, S.; Wormke, M. Receptor r Cross-Talk and Mechanisms of Action. Chem. Res. Toxicol. 2003, 16, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Agius, C.; Roberts, R.J. Melano-Macrophage Centres and Their Role in Fish Pathology. J. Fish Dis. 2003, 26, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Steinel, N.C.; Bolnick, D.I. Melanomacrophage Centers as a Histological Indicator of Immune Function in Fish and Other Poikilotherms. Front. Immunol. 2017, 8, 827. [Google Scholar] [CrossRef] [Green Version]
- Carreras-Colom, E.; Constenla, M.; Dallarés, S.; Carrassón, M. Natural Variability and Potential Use of Melanomacrophage Centres as Indicators of Pollution in Fish Species from the NW Mediterranean Sea. Mar. Pollut. Bull. 2022, 176, 113441. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Length | Weight | dMMC | dMM | MMC/mm2 | MM/mm2 | |
|---|---|---|---|---|---|---|
| Length | 1 | 0.982066 | 0.627270 | −0.628303 | 0.820992 | −0.546776 |
| Weight | 0.982066 | 1 | 0.642745 | −0.532282 | 0.817944 | −0.436625 |
| dMMC | 0.627270 | 0.642745 | 1 | −0.276178 | 0.550470 | −0.244675 |
| dMM | −0.628303 | −0.532282 | −0.276178 | 1 | −0.442146 | 0.954888 |
| MMC/mm2 | 0.820992 | 0.817944 | 0.550470 | −0.442146 | 1 | −0.347469 |
| MM/mm2 | −0.546776 | −0.436625 | −0.244675 | 0.954888 | −0.347469 | 1 |
| Length | Weight | dLipids | |
|---|---|---|---|
| Length | 1 | 0.982066 | −0.52939 |
| Weight | 0.982066 | 1 | −0.40834 |
| dLipids | −0.52939 | −0.40834 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gioacchini, G.; Filippi, S.; Debernardis, R.; Marisaldi, L.; Aiese Cigliano, R.; Carnevali, O. A Window of Vulnerability: Chronic Environmental Stress Does Not Impair Reproduction in the Swordfish Xiphias gladius. Animals 2023, 13, 269. https://doi.org/10.3390/ani13020269
Gioacchini G, Filippi S, Debernardis R, Marisaldi L, Aiese Cigliano R, Carnevali O. A Window of Vulnerability: Chronic Environmental Stress Does Not Impair Reproduction in the Swordfish Xiphias gladius. Animals. 2023; 13(2):269. https://doi.org/10.3390/ani13020269
Chicago/Turabian StyleGioacchini, Giorgia, Sara Filippi, Rossella Debernardis, Luca Marisaldi, Riccardo Aiese Cigliano, and Oliana Carnevali. 2023. "A Window of Vulnerability: Chronic Environmental Stress Does Not Impair Reproduction in the Swordfish Xiphias gladius" Animals 13, no. 2: 269. https://doi.org/10.3390/ani13020269