
Lipid Characteristics of the Muscle and Perirenal Fat in Young Tudanca Bulls Fed on Different Levels of Grass Silage
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Material
2.2. Chemical Analysis
2.3. Statistical Analysis
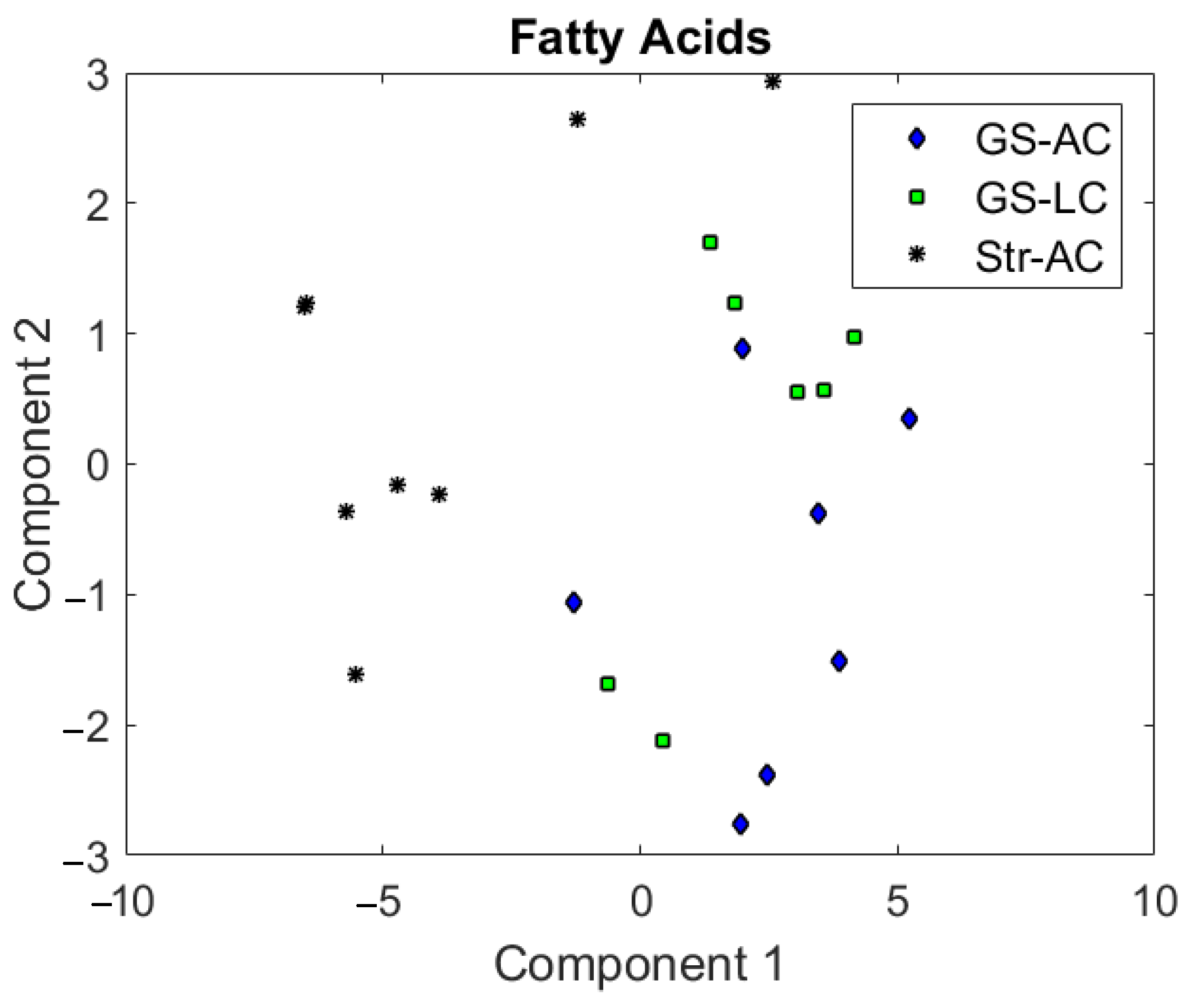
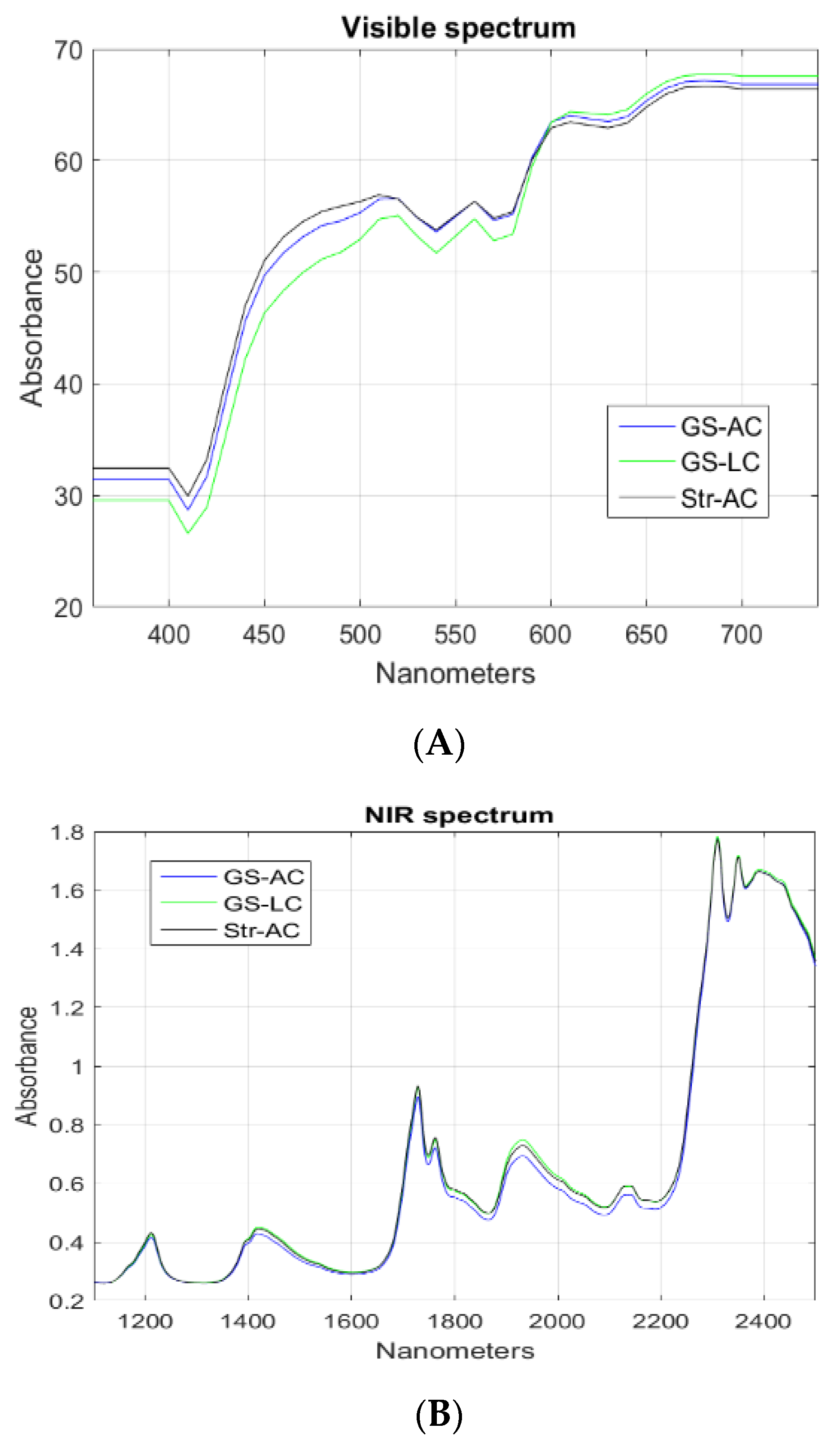
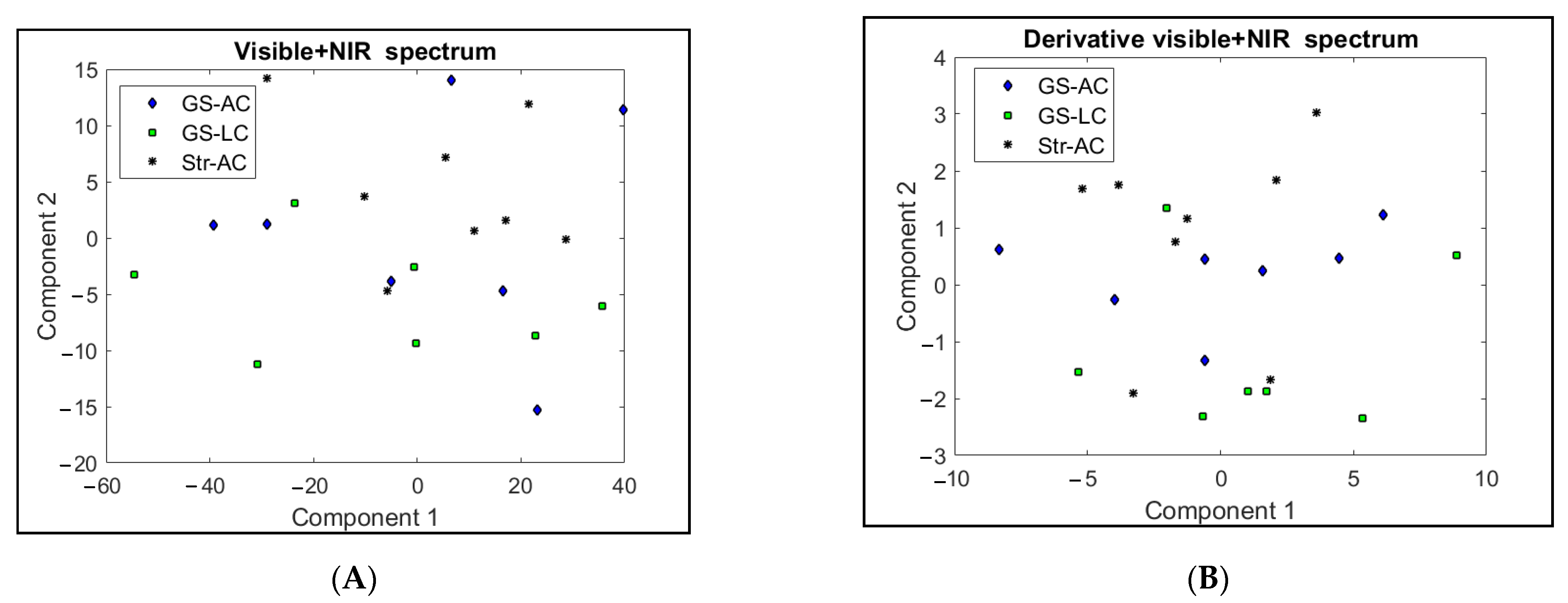
3. Results and Discussion
3.1. Intramuscular Fat Composition
3.2. Perirenal Fat Characteristics and Differentiation between Feeding Systems
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moloney, A.P.; O’Riordan, E.G.; McGee, M.; O’Sullivan, M.G.; Kerry, J.P.; Wilson, S.S.; Monahan, F.J.; Kelly, A.K.; McNenamin, K.; Morán, L. Carcass characteristics, colour and eating quality of beef from late maturing suckler bulls finished at pasture with or without concentrate supplementation or indoors on a high concentrate ration. Anim. Prod. Sci. 2022, 62, 590–600. [Google Scholar] [CrossRef]
- Rodríguez-Vázquez, R.; Pateiro, M.; López-Pedrouso, M.; Gende, A.; Crecente, S.; Serrano, M.P.; González, J.; Lorenzo, J.M.; Zapata, C.; Franco, D. Influence of production system and finishing feeding on meat quality of Rubia Gallega calves. Span. J. Agric. Res. 2020, 18, e0606. [Google Scholar] [CrossRef]
- Prache, S.; Kondjoyan, N.; Delfosse, O.; Chauveau-Duriot, B.; Andueza, D.; Cornu, A. Discrimination of pasture-fed lambs from lambs fed dehydrated alfalfa indoors using different compounds measured in the fat, meat and plasma. Animal 2009, 3, 598–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, R.B.; Kerth, C.R.; Gentry, J.G.; Prevatt, J.W.; Braden, K.W.; Jones, A.R. Determining acceptance of domestic forage- or grain-finished beef by consumers from three southeastern U.S. States. J. Food Sci. 2006, 71, S542–S546. [Google Scholar] [CrossRef]
- Xue, H.; Mainville, D.; You, W.; Nayga Jr, R.M. Consumer preferences and willingness to pay for grass-fed beef: Empirical evidence from in-store experiments. Food Qual. Prefer. 2010, 21, 857–866. [Google Scholar] [CrossRef]
- Cozzi, G.; Brscic, M.; Gottardo, F. Main critical factors affecting the welfare of beef cattle and veal calves raised under intensive rearing systems in Italy: A review. Ital. J. Anim. Sci. 2008, 8 (Suppl. 1), 67–80. [Google Scholar] [CrossRef] [Green Version]
- Daley, C.; Abbott, A.; Doyle, P.S.; Nader, G.A.; Larson, S. A review of fatty acid profiles and antioxidant content in grass-fed and grain-fed beef. Nutr. J. 2010, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Miller, R. Drivers of consumer liking for beef, pork, and lamb: A review. Foods 2020, 9, 428. [Google Scholar] [CrossRef] [Green Version]
- Venkata Reddy, B.; Sivakumar, A.S.; Jeong, D.W.; Woo, Y.B.; Park, S.J.; Lee, S.Y.; Byun, J.Y.; Kim, C.H.; Cho, S.H.; Hawang, I. Beef quality traits of heifer in comparison with steer, bull and cow at various feeding environments. Anim. Sci. J. 2015, 86, 1–16. [Google Scholar] [CrossRef]
- Monahan, F.J.; Schmidt, O.; Moloney, A.P. Meat provenance: Authentication of geographical origin and dietary background of meat. Meat Sci. 2018, 144, 2–14. [Google Scholar] [CrossRef]
- Prache, S.; Martin, B.; Coppa, M. Authentication of grass-fed meat and dairy products from cattle and sheep. Animal 2020, 14, 854–863. [Google Scholar] [CrossRef] [Green Version]
- Monahan, F.J.; Moloney, A.P.; Osorio, M.T.; Röhrle, F.T.; Schmidt, O.; Brennan, L. Authentication of grass-fed beef using bovine muscle, hair or urine. Trends Food Sci. Technol. 2012, 28, 69–76. [Google Scholar] [CrossRef]
- Alfaia, C.P.M.; Alves, S.P.; Martins, S.I.V.; Costa, A.S.H.; Fontes, C.M.G.A.; Lemos, J.P.C.; Bessa, R.J.B.; Prates, J.A.M. Effect of the feeding system on intramuscular fatty acids and conjugated linoleic acid isomers of beef cattle, with emphasis on their nutritional value and discriminatory ability. Food Chem. 2009, 114, 939–946. [Google Scholar] [CrossRef]
- Cama-Moncunill, R.; Moloney, A.P.; Röhrle, F.T.; Luciano, G.; Monahan, F.J. Canonical discriminant analysis of the fatty acid profile of muscle to authenticate beef from grass-fed and other beef production systems: Model development and validation. Food Control 2021, 122, 107820. [Google Scholar] [CrossRef]
- Röhrle, F.T.; Moloney, A.P.; Osorio, M.T.; Luciano, G.; Priolo, A.; Caplan, P.; Monahan, F.J. Carotenoid, colour and reflectance measurements in bovine adipose tissue to discriminate between beef from different feeding systems. Meat Sci. 2011, 88, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Osorio, M.T.; Dwney, G.; Moloney, A.P.; Röhrle, F.T.; Luciano, G. Beef authentication using dietary markers: Chemometric selection and modelling of significant beef biomarkers using concatenated data from multiple analytical methods. Food Chem. 2014, 141, 2795–2801. [Google Scholar] [CrossRef]
- Martínez-Fernández, A.; Argamentería, A.; de la Roza, B. Manejo de Forrajes Para Ensilar; Servicio Regional de Investigación y Desarrollo Agroalimentario del Principado de Asturias: Asturias, Spain, 2014; 280p. [Google Scholar]
- Serrano, E.; Humada, M.J.; Caro, I.; Vázquez, I.; Olaizola, A.M.; Resano, H.; Soto, S.; Mateo, J. Performance, carcass characteristics, economic margin and meat quality in young Tudanca bulls fed on two levels of grass silage and concentrate. Span. J. Agric. Res. 2018, 16, e0609. [Google Scholar] [CrossRef]
- Blanco, M.; Ripoll, G.; Delavaud, C.; Casasúsa, I. Performance, carcass and meat quality of young bulls, steers and heifers slaughtered at a common body weight. Livest. Sci. 2020, 240, 104–156. [Google Scholar] [CrossRef]
- Andrés, S.; Jaramillo, E.; Mateo, J.; Caro, I.; Carballo, D.E.; López, S.; Giráldez, F.J. Grain grinding size of cereals in complete pelleted diets for growing lambs: Effects on animal performance, carcass and meat quality traits. Meat Sci. 2019, 157, 107874. [Google Scholar] [CrossRef]
- Weihrauch, J.L.; Psati, L.P.; Anderson, B.A.; Exler, J.; Blanco, M.; Ripoll, G.; Delavaud, C.; Casasúsa, I. Lipid conversion factors for calculating fatty acid contents of foods. J. Am. Oil Chem. Soc. 1977, 54, 36–40. [Google Scholar] [CrossRef]
- Vieira, C.; Fernández-Diez, A.; Mateo, J.; Bodas, R.; Soto, S.; Manso, T. Effects of addition of different vegetable oils to lactating dairy ewes’ diet on meat quality characteristics of suckling lambs reared on the ewes’ milk. Meat Sci. 2012, 91, 277–283. [Google Scholar] [CrossRef]
- Nam, K.C.; Ahn, D.U. Combination of anaerobic and vacuum packaging to control lipid oxidation and off-odour volatiles of irradiate draw turkey breast. Meat Sci. 2003, 63, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Mestre-Prates, J.A.; Gonçalvez-Quaresma, M.A.; Branquinho Bessa, R.J.; Andrade-Fontes, C.M.G.; Mateus-Alfaia, C.M.P. Simultaneous HPLC quantification of total cholesterol, tocopherols and β-carotene in Barrosa-PDO veal. Food Chem. 2006, 94, 469–477. [Google Scholar] [CrossRef]
- Jin, Q.; Cheng, H.; Wan, F.; Bi, Y.; Liu, G.; Liu, X.; Zhao, H.; You, W.; Liu, Y.; Tan, X. Effects of feeding β-carotene on levels of β-carotene and vitamin A in blood and tissues of beef cattle and the effects on beef quality. Meat Sci. 2015, 110, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Humada, M.J.; Sañudo, C.; Serrano, E. Chemical composition, vitamin E content, lipid oxidation, colour and cooking losses in meat from Tudanca bulls finished on semi-extensive or intensive systems and slaughtered at 12 or 14 months. Meat Sci. 2014, 96, 908–915. [Google Scholar] [CrossRef]
- AOAC. Official method 991.36 fat (crude) in meat and meat products. In Official Methods of Analysis of the AOAC International, 16th ed.; Cunniff, P., Ed.; AOAC: Gaithersburg, MD, USA, 1999; Volume II, pp. 1–15. [Google Scholar]
- Prieto, N.; Andrés, S.; Giráldez, F.J.; Mantecón, A.R.; Lavín, M.P. Potential use of near infrared reflectance spectroscopy (NIRS) for the estimation of chemical composition of oxen meat samples. Meat Sci. 2006, 74, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Jolliffe, I.T. Principal component analysis. In Springer Series in Statistics; Springer: New York, NY, USA, 2002. [Google Scholar]
- Wang, H.; He, Y.; Li, H.; Wu, F.; Qiu, Q.; Niu, W.; Gao, Z.; Su, H.; Cao, B. Rumen fermentation, intramuscular fat fatty acid profiles and related rumen bacterial populations of Holstein bulls fed diets with different energy levels. Appl. Microbiol. Biotechnol. 2019, 103, 4931–4942. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, B.; Xiao, J.; Guo, M.; Zhao, S.; Hu, M.; Cui, Y.; Li, D.; Wang, C.; Ma, S.; et al. Effects of different roughage diets on fattening performance, meat quality, fatty acid composition, and rumen microbe in steers. Front. Nutr. 2022, 9, 885069. [Google Scholar] [CrossRef]
- Warren, H.E.; Scollan, N.D.; Enser, M.; Hughes, S.I.; Richarson, R.I.; Wood, J.D. Effects of breed and a concentrate or grass silage diet on beef quality in cattle of 3 ages. I: Animal performance, carcass quality and muscle fatty acid composition. Meat Sci. 2008, 78, 256–269. [Google Scholar] [CrossRef]
- Nogoy, K.M.; Sun, B.; Shin, S.; Lee, Y.; Xiang, Z.; Seong, L.; Choi, H.; Park, S. Fatty acid composition of grain- and grass-fed beef and their nutritional value and health implication. Food Sci. Anim. Resour. 2022, 42, 18–33. [Google Scholar] [CrossRef]
- Vahmani, P.; Ponnampalam, E.N.; Kraft, J.; Mapiye, C.; Bermingham, E.N.; Watkins, P.J.; Proctor, S.D.; Dugan, M.E.R. Bioactivity and health effects of ruminant meat lipids. Meat Sci. 2020, 165, 108114. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Resconi, V.; Escudero, A.; Campo, M.M. The development of aromas in ruminant meat. Molecules 2013, 18, 6748–6781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Xie, J.; Zhang, L.; Du, R.; Cao, C.; Wang, M.; Acree, T.; Sun, B. Aromatic effect of fat and oxidized fat on a meat-like model reaction system of cysteine and glucose. Flavour Fragr. J. 2015, 30, 320–329. [Google Scholar] [CrossRef]
- Elmore, J.S.; Cooper, S.L.; Enser, M.; Mottram, D.S.; Sinclair, L.A.; Wilkinson, R.G. Dietary manipulation of fatty acid composition in lamb meat and its effect on the volatile aroma compounds of grilled lambs. Meat Sci. 2005, 69, 233–242. [Google Scholar] [CrossRef]
- Mezgebo, G.B.; Monahan, F.J.; McGee, M.; O’Riordan, E.G.; Richardson, I.; Brunton, N.; Moloney, A.P. Fatty acid, volatile and sensory characteristics of beef as affected by grass silage or pasture in the bovine diet. Food Chem. 2017, 235, 86–97. [Google Scholar] [CrossRef] [Green Version]
- Prieto, N.; Dugan, M.E.R.; López-Campos, Ó.; Aalhus, J.L.; Uttaro, B.; Resconi, V.; Escudero, A.; Campo, M.M. At line prediction of PUFA and biohydrogenation intermediates in perirenal and subcutaneous fat from cattle fed sunflower or flaxseed by near infrared spectroscopy. Meat Sci. 2013, 94, 27–33. [Google Scholar] [CrossRef]
- Dian, P.H.M.; Andueza, D.; Jestin, M.; Prado, I.N.; Prache, S. Comparison of visible and near infrared reflectance spectroscopy to discriminate between pasture-fed and concentrate-fed lamb carcasses. Meat Sci. 2008, 80, 1157–1164. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Concentrate (n = 2) | Silage (n = 2) | Straw (n = 2) | |
|---|---|---|---|
| Dry matter (DM) | 90.4 ± 2.3 | 27.1 ± 2.9 | 88.7 ± 4.1 |
| Crude protein | 14.5 ± 0.4 | 10.1 ± 3.0 | 4.4 ± 0.5 |
| Ether extract | 3.7 ± 0.3 | ND | ND |
| Acid detergent fibre | 13.0 ± 1.8 | 32.4 ± 3.8 | 49.8 ± 3.9 |
| Neutral detergent fibre | 24.1 ± 2.9 | 59.2 ± 2.5 | 80.8 ± 6.2 |
| Ash | 6.2 ± 0.8 | 11.0 ± 2.6 | 7.6 ± 1.4 |
| Net energy (milk feed units/ kg DM) | 0.99 ± 0.04 | 0.75 ± 0.05 | 0.43 ± 0.04 |
| 14:0 & | ND | 2.5 ± 0.5 | ND |
| 16:0 & | 17.7 ± 1.7 | 23.9 ± 3.0 | 25.7 ± 2.1 |
| 18:0 & | 3.4 ± 0.3 | 3.2 ± 0.6 | 8.2 ± 0.5 |
| c9-18:1 & | 20.1 ± 2.3 | 5.7 ± 0.9 | 13.8 ± 1.5 |
| 18:2n − 6 & | 55.9 ± 4.0 | 14.8 ± 1.9 | 23.5 ± 2.0 |
| 18:3n − 3 & | 2.9 ± 0.5 | 49.9 ± 9.8 | 28.8 ± 3.8 |
| β-carotene | ND | 79.9 ± 9.2 | ND |
| Retinol | 2.4 ± 0.8 | ND | ND |
| α-tocopherol | 40.0 ± 3.8 | 37.0 ± 9.0 | 1.9 ± 0.3 |
| γ-tocopherol | 2.2 ± 0.5 | 1.3 ± 1.0 | 1.1 ± 0.6 |
| δ-tocopherol | 0.22 ± 0.05 | 0.11 ± 0.04 | ND |
| GS-AC (n = 7) | GS-LC (n = 7) | Str-AC (n = 8) | SEM | p-Value | |
|---|---|---|---|---|---|
| Fatty acids (mg/100 beef) | |||||
| SFA | 1787.0 | 1103.5 | 1310.0 | 0.637 | 0.12 |
| BCFA | 3.2 | 3.0 | 4.7 | 0.020 | 0.69 |
| OCFA | 83.8 | 75.6 | 103.5 | 0.150 | 0.40 |
| 12:0 | ND | ND | 1.0 | 0.007 | - |
| 14:0 | 87.6 | 42.2 | 51.2 | 0.127 | 0.08 |
| iso-C15:0 | 3.2 | 3.0 | 4.7 | 0.020 | 0.69 |
| 15:0 | 46.0 | 45.1 | 53.0 | 0.113 | 0.64 |
| 16:0 | 962.5 | 576.2 | 671.0 | 0.551 | 0.11 |
| 17:0 | 19.7 | 14.3 | 24.8 | 0.053 | 0.32 |
| 18:0 | 668.5 | 422.1 | 504.5 | 0.288 | 0.11 |
| 20:0 | 3.0 | 1.1 | 2.9 | 0.012 | 0.18 |
| 22:0 | ND | 2.5 | 1.6 | 0.025 | - |
| MUFA | 1602.2 | 982.1 | 1255.7 | 0.457 | 0.14 |
| c9-14:1 | 8.0 | 3.0 | 4.4 | 0.028 | 0.23 |
| c9-16:1 | 105.3 | 56.2 | 69.5 | 0.105 | 0.12 |
| c9-17:1 | 15.0 | 13.2 | 21.0 | 0.048 | 0.43 |
| t10-,t11-18:1 | 58.9 b | 50.8 b | 133.1 a | 0.322 | 0.02 |
| c9-18:1 | 1374.6 | 821.4 | 949.2 | 0.619 | 0.08 |
| c11-18:1 | 31.7 b | 34.2 b | 69.7 a | 0.186 | 0.04 |
| c12-18:1 | 1.8 | 0.8 | 3.28 | 0.014 | 0.23 |
| c15-18:1 | 2.9 | 0.2 | 1.9 | 0.016 | 0.17 |
| c11-20:1 | 3.8 | 1.9 | 3.4 | 0.012 | 0.34 |
| PUFA | 416.8 | 383.0 | 566.5 | 0.877 | 0.06 |
| n − 6 | 355.8 b | 327.7 b | 508.2 a | 0.816 | 0.03 |
| n − 3 | 43.5 | 43.0 | 39.2 | 0.089 | 0.84 |
| n − 6/n − 3 | 8.5 b | 7.6 b | 13.3 a | 0.622 | <0.001 |
| t,t-18:2 & | 3.1 | 2.2 | 3.1 | 0.020 | 0.49 |
| 18:2n − 6 | 272.4 b | 243.0 b | 386.4 a | 0.596 | 0.02 |
| 18:3n − 6 | ND | 0.01 | 0.04 | 0.006 | - |
| 18:3n − 3 | 20.6 | 18.8 | 15.3 | 0.040 | 0.37 |
| t,c-c,t-18:2 & | 11.3 | 6.8 | 6.7 | 0.016 | 0.06 |
| 20:2n − 3 | 3.1 | 2.7 b | 6.0 | 0.022 | 0.055 |
| 20:2n − 6 | 1.4 | 3.1 | 2.5 | 0.020 | 0.055 |
| 20:3n − 6 | 13.5 | 13.0 | 20.6 | 0.043 | 0.057 |
| 20:4n − 6 | 65.8 | 67.2 | 90.9 | 0.177 | 0.10 |
| 20:5n − 3 | 6.5 | 8.9 | 8.5 | 0.030 | 0.34 |
| 22:4n − 6 | 4.1 b | 4.4 b | 9.2 a | 0.028 | 0.01 |
| 22:5n − 3 | 16.3 | 15.3 | 15.3 | 0.028 | 0.92 |
| Vitamins (µg/g muscle) | |||||
| β-carotene | 0.60 | 0.63 | ND | 0.077 | - |
| Retinol | 0.25 | 0.21 | 0.21 | 0.015 | 0.42 |
| α-tocopherol | 5.68 | 4.71 | 4.34 | 0.332 | 0.25 |
| TBARS $ | |||||
| Day 0 | 1.10 | 0.95 | 1.37 | 0.070 | 0.08 |
| ΔTBARS | 0.40 | 0.48 | 0.44 | 0.077 | 0.77 |
| GS-AC | GS-LC | Str-AC | SEM | p-Level | |
|---|---|---|---|---|---|
| Aldehydes | 516.2 a | 241.3 b | 529.9 a | 50.23 | 0.046 |
| Pentanal (701) & | 52.9 | 18.6 | 49.9 | 6.95 | 0.08 |
| Hexanal (803) | 341.2 | 152.7 | 327.4 | 36.26 | 0.08 |
| Heptanal (901) | 57.9 | 21.7 | 59.8 | 7.58 | 0.06 |
| 2-Heptenal (958) | 0.7 | 0.1 | 0.7 | 0.14 | 0.11 |
| Octanal (1002) | 28.6 | 14.2 | 40.0 | 4.76 | 0.08 |
| Octenal (1059) | 1.2 | 0.3 | 1.3 | 0.22 | 0.12 |
| Nonanal (1103) | 30.1 | 27.7 | 39.0 | 3.75 | 0.38 |
| 2-Nonenal (1160) | 0.4 | 0.1 | 0.5 | 0.09 | 0.08 |
| Decanal (1204) | 1.1 | 1.6 | 1.6 | 0.19 | 0.40 |
| Dodecanal (1406) | 0.2 | 0.5 | 0.4 | 0.09 | 0.49 |
| Alcohols | 23.6 | 17.2 | 28.0 | 3.17 | 0.39 |
| Pentanol (775) | 9.3 | 2.9 | 7.6 | 1.32 | 0.13 |
| Hexanol (875) | 1.3 | 0.7 | 1.6 | 0.24 | 0.26 |
| Heptanol (973) | 1.9 | 1.7 | 2.3 | 0.55 | 0.90 |
| 1-Octen-3-ol (981) | 6.9 | 4.6 | 8.2 | 1.06 | 0.39 |
| 2-Ethyl-hexanol (1029) | 3.1 | 4.7 | 4.8 | 1.25 | 0.24 |
| Octanol (1073) | 1.8 | 2.4 | 3.3 | 0.43 | 0.36 |
| Ketones | 79.6 | 52.4 | 56.5 | 7.06 | 0.20 |
| 3-Hydroxy-2-butanone (740) | 2.3 | 2.4 | 7.2 | 1.33 | 0.22 |
| Heptanone (890) | 2.2 | 1.7 | 3.0 | 0.32 | 0.25 |
| Octanedione (984) | 5.2 | 2.9 | 5.6 | 1.15 | 0.20 |
| Hydrocarbons | 8.0 | 10.1 | 5.4 | 1.06 | 0.20 |
| Octane (800) | 3.0 | 6.3 | 3.1 | 0.56 | 0.051 |
| 2,2,4,6,6-Pentamethylheptane (985) | 2.0 | 1.6 | 0.1 | 0.63 | 0.40 |
| Unknown (1033) | 1.4 | 0.2 | 1.0 | 0.28 | 0.22 |
| Furans | 8.8 | 3.6 | 9.6 | 1.47 | 0.21 |
| 2-Penthylfuran (988) | 8.8 | 3.6 | 9.6 | 1.47 | 0.21 |
| Fatty acids | 4.1 | 3.3 | 2.8 | 0.50 | 0.57 |
| Hexanoic acid (1008) | 2.7 | 0.6 | 1.6 | 0.43 | 0.15 |
| Dodecanoic acid (1691) | 0.4 | 1.0 | 0.4 | 0.18 | 0.26 |
| Benzene compounds | 45.8 | 45.8 | 48.8 | 3.36 | 0.92 |
| Toluene (767) | 5.0 | 3.6 | 4.1 | 0.51 | 0.56 |
| Benzaldehyde (965) | 40.5 | 40.4 | 43.9 | 3.20 | 0.88 |
| Sulphur compounds | 10.2 | 5.3 | 7.6 | 1.58 | 0.48 |
| Dimethyl disulfide (742) | 3.0 | 1.6 | 1.9 | 0.56 | 0.61 |
| Dimethyl trisulfide (970) | 5.4 | 0.8 | 2.3 | 1.14 | 0.29 |
| GS-AC | GS-LC | Str-AC | SEM | p-Level | |
|---|---|---|---|---|---|
| Fat (% of fresh weight) | 89.5 a | 87.3 b | 88.3 ab | 0.294 | 0.03 |
| Fatty acids (mg/100 fat depot) | |||||
| SFA | 58,095 a | 56,541 a | 53,371 b | 718.1 | 0.01 |
| BCFA | 2183 | 2442 | 2080 | 89.9 | 0.25 |
| OCFA | 3775 | 4275 | 4270 | 118.0 | 0.14 |
| 10:0 | 50 | 49 | 56 | 1.8 | 0.21 |
| 12:0 | 54 | 58 | 65 | 2.8 | 0.30 |
| iso-14:0 | 44 a | 54 a | 29 b | 3.6 | 0.009 |
| 14:0 | 2759 | 2629 | 2897 | 82.0 | 0.42 |
| iso-15:0 | 176 b | 200 b | 120 a | 10.9 | 0.002 |
| anteiso-15:0 | 202 | 241 | 234 | 10.16 | 0.26 |
| 15:0 | 331 | 366 | 411 | 16.2 | 0.12 |
| iso-16:0 | 185 a | 208 a | 135 b | 10.3 | 0.004 |
| 16:0 | 21,084 | 20,248 | 20,565 | 293.5 | 0.54 |
| iso-17:0 | 364 b | 517 a | 369 b | 19.8 | <0.001 |
| anteiso-17:0 | 1119 | 1131 | 1113 | 54.3 | 0.99 |
| 17:0 | 1218 b | 1370 b | 1603 a | 52.2 | 0.003 |
| iso-18:0 | 88 | 91 | 80 | 4.4 | 0.56 |
| 18:0 | 29,949 a | 28,849 a | 25,293 b | 644.2 | 0.002 |
| 19:0 | 81 | 95 | 90 | 7.3 | 0.75 |
| 20:0 | 340 a | 364 a | 282 b | 13.1 | 0.02 |
| 22:0 | 44 | 68 | 30 | 6.8 | 0.07 |
| MUFA | 31,599 b | 30,568 b | 33,825 a | 460.0 | 0.005 |
| c9-14:1 | 136 | 115 | 151 | 8.5 | 0.24 |
| t9-16:1 | 164 | 181 | 175 | 9.0 | 0.76 |
| c9-16:1 | 621 | 665 | 683 | 48.7 | 0.88 |
| c9-17:1 | 248 | 282 | 296 | 15.4 | 0.45 |
| t10-,t11-18:1 | 4832 b | 5473 b | 8862 a | 444.8 | <0.001 |
| c9-18:1 | 25,040 a | 23,298 b | 23,252 b | 296.3 | 0.013 |
| c11-18:1 | 463 a | 394 a | 257 b | 31.3 | 0.013 |
| c7-19:1 | 19 | 18 | 14 | 7.2 | 0.97 |
| c8-20:1 | 30 | 35 | 29 | 4.1 | 0.80 |
| c11-20:1 | 46 | 104 | 105 | 14.0 | 0.15 |
| PUFA | 4321 b | 4459 b | 5667 a | 189.3 | <0.001 |
| n − 6 | 3260 b | 3388 b | 4778 a | 196.3 | <0.001 |
| n − 6/n − 3 | 6.97 b | 6.63 b | 8.35 a | 0.522 | <0.001 |
| t,t-18:2 & | 136 | 122 | 195 | 9.7 | 0.71 |
| 18:2n − 6 | 3232 b | 3370 b | 4698 a | 190.8 | <0.001 |
| 18:3n − 3 | 473 ab | 514 a | 432 b | 12.7 | 0.02 |
| t,c+c,t-18:2 & | 442 a | 365 ab | 322 b | 21.1 | 0.04 |
| 20:2n − 6 | 48 | 59 | 85 | 7.1 | 0.07 |
| 20:4n − 6 | 24 ab | 14 b | 39 a | 4.3 | 0.047 |
| 22:2n − 6 | 6 b | 5 b | 15 a | 4 | 0.04 |
| Vitamins (µg/g perirenal fat) | |||||
| β-carotene | 0.24 b | 0.58 a | 0.03 c | 0.060 | <0.001 |
| Retinol | 1.94 | 2.12 | 1.79 | 0.200 | 0.22 |
| GS-AC | GS-LC | Str-AC | SEM | p-Level | |
|---|---|---|---|---|---|
| Aldehydes | 97.0 | 43.1 | 120.2 | 75.96 | 0.14 |
| 2-Pentenal (710) | 2.2 | 1.8 | 2.4 | 1.41 | 0.70 |
| Hexanal (798) | 71.7 | 32.4 | 93.1 | 58.65 | 0.13 |
| 2-Hexenal (842) | 1.1 | 0.2 | 1.2 | 1.01 | 0.17 |
| Heptanal (895) | 16.4 | 5.1 | 14.5 | 10.81 | 0.11 |
| 2-Heptenal (950) | 1.6 | 0.8 | 2.6 | 1.90 | 0.17 |
| Octanal (998) | 1.8 | 1.0 | 2.6 | 1.97 | 0.31 |
| Nonanal (1100) | 1.2 | 1.0 | 1.8 | 1.15 | 0.39 |
| 2-Nonenal (1157) | 0.6 | 0.4 | 0.8 | 0.51 | 0.21 |
| Alcohols | 3.1 | 1.0 | 3.0 | 2.15 | 0.19 |
| Pentanol (772) | 2.6 | 1.0 | 2.4 | 1.74 | 0.15 |
| Hydrocarbons | 15.1 | 8.5 | 8.6 | 6.81 | 0.35 |
| Octadiene (783) | 0.8 | 0.3 | 0.4 | 0.55 | 0.18 |
| Octene (791) | 5.5 | 3.6 | 2.6 | 2.64 | 0.50 |
| Nonene (885) | 1.9 | 1.0 | 1.1 | 0.97 | 0.16 |
| 2,2,7,7-Tetramethyl-octane (982) | 3.1 | 2.1 | 2.4 | 1.43 | 0.43 |
| Decene (986) | 1.0 | 0.8 | 0.7 | 0.61 | 0.57 |
| Acids and lactones | 4.2 | 4.4 | 6.3 | 2.98 | 0.36 |
| Hexanoic acid (999) | 2.9 | 3.2 | 4.9 | 2.47 | 0.27 |
| Benzene compounds | 1.9 | 2.9 | 1.0 | 2.86 | 0.58 |
| Toluene (768) | 1.6 | 2.3 | 0.6 | 2.87 | 0.51 |
| GS-AC | GS-LC | Str-AC | SEM | p-Level | |
|---|---|---|---|---|---|
| Colour | |||||
| L* | 79.93 | 79.10 | 80.12 | 0.741 | 0.71 |
| a* | 0.98 b | 1.84 a | 0.79 b | 0.270 | 0.03 |
| b* | 9.79 b | 11.91 a | 8.40 b | 0.510 | <0.001 |
| I450–510 & | 171.1 b | 237.4 a | 122.8 c | 11.81 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mateo, J.; Caro, I.; Soto, S.; Andrés, S.; Aláiz-Rodríguez, R.; Serrano, E. Lipid Characteristics of the Muscle and Perirenal Fat in Young Tudanca Bulls Fed on Different Levels of Grass Silage. Animals 2023, 13, 261. https://doi.org/10.3390/ani13020261
Mateo J, Caro I, Soto S, Andrés S, Aláiz-Rodríguez R, Serrano E. Lipid Characteristics of the Muscle and Perirenal Fat in Young Tudanca Bulls Fed on Different Levels of Grass Silage. Animals. 2023; 13(2):261. https://doi.org/10.3390/ani13020261
Chicago/Turabian StyleMateo, Javier, Irma Caro, Sergio Soto, Sonia Andrés, Rocío Aláiz-Rodríguez, and Emma Serrano. 2023. "Lipid Characteristics of the Muscle and Perirenal Fat in Young Tudanca Bulls Fed on Different Levels of Grass Silage" Animals 13, no. 2: 261. https://doi.org/10.3390/ani13020261