
Serum Anti-Müllerian Hormone Levels and Estrous Monitoring of GnRH Agonist Deslorelin-Induced Estrus in Bitches: A Pilot Study
 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Estrus Induction
2.3. Cytological Examination
2.4. Hormone Analysis
2.4.1. AMH Assay
2.4.2. Estradiol-17ß Assay
2.4.3. Progesterone Assay
2.5. Ultrasonographic Examination of Ovaries
2.6. Statistical Analysis
3. Results
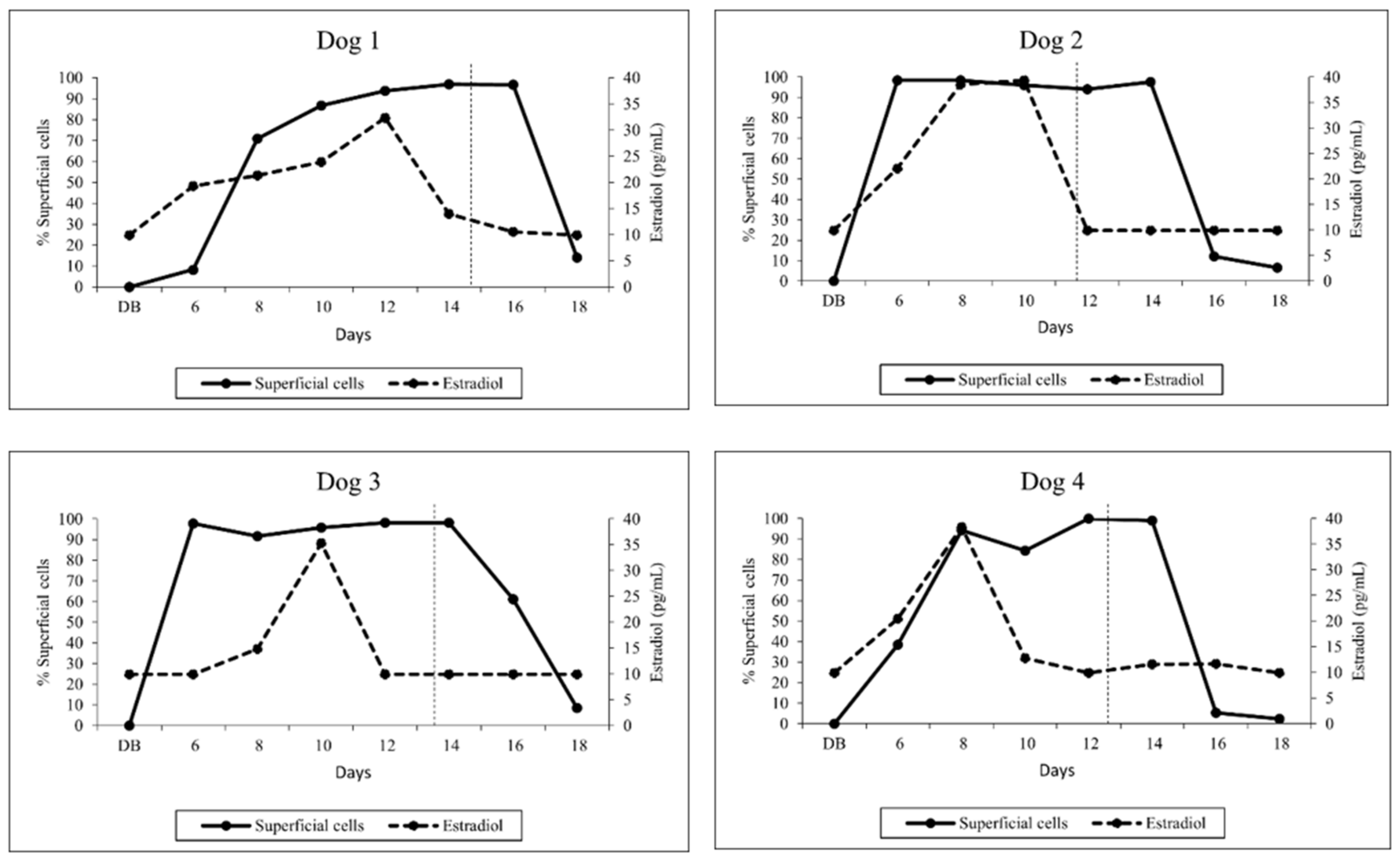
3.1. Vaginal Cytology
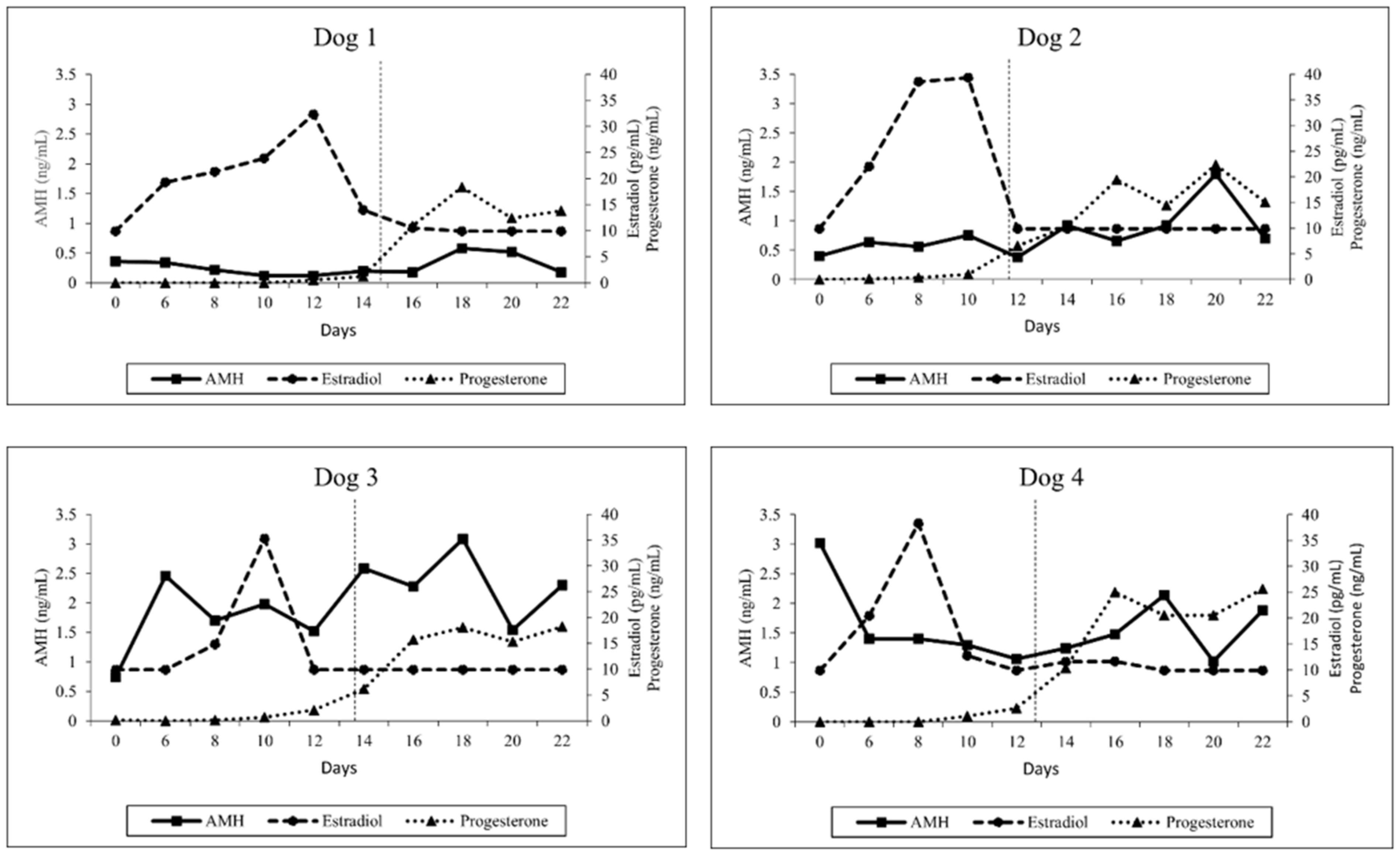
3.2. Hormone Analysis
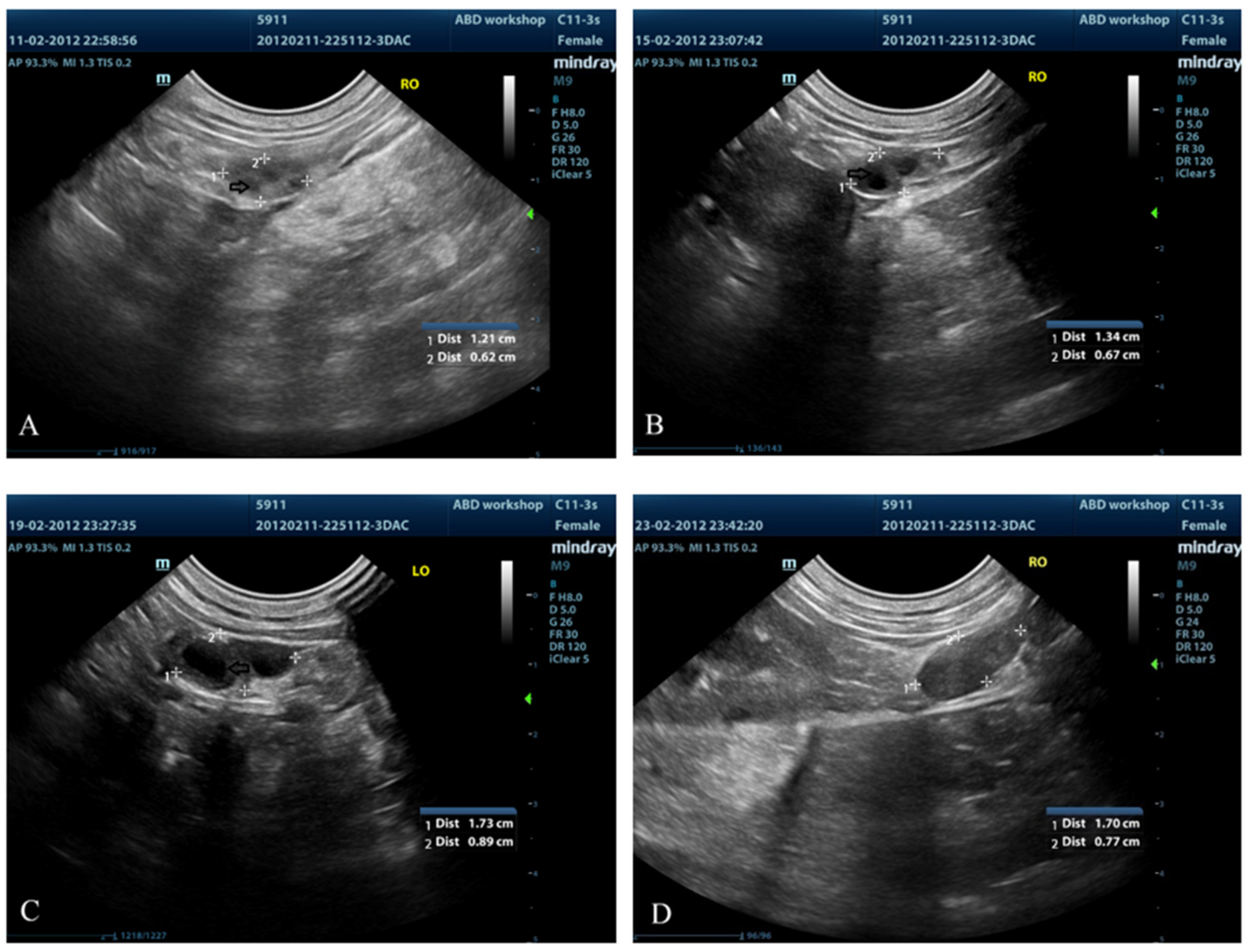
3.3. Ovarian Ultrasound
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trigg, T.E. Use of a GnRH analogue implant to produce reversible long-term suppression of reproductive function in male and female domestic dogs. J. Reprod. Fertil. Suppl. 2001, 57, 255. [Google Scholar] [PubMed]
- Rubion, S.; Desmoulins, P.O.; Rivière-Godet, E.; Kinziger, M.; Salavert, F.; Rutten, F.; Flochlay-Sigognault, A.; Driancourt, M.A. Treatment with a subcutaneous GnRH agonist containing controlled release device reversibly prevents puberty in bitches. Theriogenology 2006, 66, 1651–1654. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, E.; Fontbonne, A. Clinical use of GnRH agonists in canine and feline species. Reprod. Domest. Anim. 2011, 46, 344–353. [Google Scholar] [CrossRef]
- Fontaine, E.; Mir, F.; Vannier, F.; Gérardin, A.; Albouy, M.; Navarro, C.; Fontbonne, A. Induction of fertile oestrus in the bitch using Deslorelin, a GnRH agonist. Theriogenology 2011, 76, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Walter, B.; Otzdorff, C.; Brugger, N.; Braun, J. Estrus induction in Beagle bitches with the GnRH-agonist implant containing 4.7 mg Deslorelin. Theriogenology 2011, 75, 1125–1129. [Google Scholar] [CrossRef] [PubMed]
- Schäfer-Somi, S.; Kaya, D.; Aslan, S. Prepubertal Use of Long-Term GnRH Agonists in Dogs: Current Knowledge and Recommendations. Animals 2022, 12, 2267. [Google Scholar] [CrossRef] [PubMed]
- Kutzler, M.A. Induction and synchronization of estrus in dogs. Theriogenology 2005, 64, 766–775. [Google Scholar] [CrossRef]
- Volkmann, D.H.; Kutzler, M.A.; Wheeler, R.; Krekeler, N. The use of deslorelin implants for the synchronization of es-trous in diestrous bitches. Theriogenology 2006, 66, 1497–1501. [Google Scholar] [CrossRef]
- Ponglowhapan, S.; Suthamnatpong, A.; Khamanarong, A.; Akaraphutoporn, E.; Kulabsri, S.; Jamikorn, U. Failure to conceive in deslorelin-induced oestrous bitches with regard to removal of hormone implants after ovulation. Thai J. Vet. Med. 2018, 48, 211–217. [Google Scholar]
- Borges, P.; Fontaine, E.; Maenhoudt, C.; Payan-Carreira, R.; Santos, N.; Leblond, E.; Fontaine, C.; Fontbonne, A. Fertility in Adult Bitches Previously Treated with a 4.7 mg Subcutaneous Deslorelin Implant. Reprod. Domest. Anim. 2015, 50, 965–971. [Google Scholar] [CrossRef]
- Kutzler, M.; Lamb, S.V.; Volkmann, D. Comparison between vestibular and subcutaneous insertion of deslorelin implants for oestrus induction in bitches. Reprod. Domest. Anim. 2009, 44 (Suppl. 2), 83–86. [Google Scholar] [CrossRef]
- Van Haaften, B.; Bevers, M.M.; van den Brom, W.E.; Okkens, A.C.; van Sluijs, F.J.; Willemse, A.H.; Dieleman, S.J. Increasing sensitivity of the pituitary to GnRH from early to late anoestrus in the beagle bitch. J. Reprod. Fertil. 1994, 101, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Wolf, T.; Meyer, H.; Kutzler, M. Litter size response to oestrous induction with deslorelin (Ovuplant®) in dogs. Reprod. Domest. Anim. 2012, 47, 387–388. [Google Scholar] [CrossRef]
- Nagashima, J.B.; Hansen, B.S.; Songsasen, N.; Travis, A.J.; Place, N.J. Anti-Müllerian hormone in the domestic dog during the anestrus to oestrous transition. Reprod. Domest. Anim. 2016, 51, 158–164. [Google Scholar] [CrossRef]
- Walter, B.; Feulner, H.; Otzdorff, C.; Klein, R.; Reese, S.; Meyer-Lindenberg, A. Changes in anti-Müllerian hormone concentrations in bitches throughout the oestrous cycle. Theriogenology 2019, 127, 114–119. [Google Scholar] [CrossRef]
- Place, N.J.; Hansen, B.S.; Cheraskin, J.L.; Cudney, S.E.; Flander, J.A.; Newmark, A.D.; Barry, B.; Scarlett, J.M. Measurement of serum anti-Müllerian hormone concentration in female dogs and cats before and after ovariohysterectomy. J. Vet. Diagn. Invest. 2011, 23, 524–527. [Google Scholar] [CrossRef] [Green Version]
- Holst, B.S. Diagnostic possibilities from a serum sample–Clinical value of new methods within small animal reproduction, with focus on anti-Müllerian hormone. Reprod. Domest. Anim. 2017, 52, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Umer, S.; Zhao, S.J.; Sammad, A.; Weldegebriall Sahlu, B.; Pang, Y.; Zhu, H. AMH: Could It Be Used as A Biomarker for Fertility and Superovulation in Domestic Animals? Genes 2019, 10, 1009. [Google Scholar] [CrossRef] [Green Version]
- Alebic, M.Š.; Stojanovic, N.; Dewailly, D. Discordance between serum anti-Müllerian hormone concentrations and antral follicle counts: Not only technical issues. Hum. Reprod. 2018, 33, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Alson, S.S.E.; Bungum, L.J.; Giwercman, A.; Henic, E. Anti-müllerian hormone levels are associated with live birth rates in ART, but the predictive ability of anti-müllerian hormone is modest. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 225, 199–204. [Google Scholar] [CrossRef]
- Bertone-Johnson, E.R.; Manson, J.E.; Purdue-Smithe, A.C.; Steiner, A.Z.; Eliassen, A.H.; Hankinson, S.E.; Rosner, B.A.; Whitcomb, B.W. Anti-Müllerian hormone levels and incidence of early natural menopause in a prospective study. Hum. Reprod. 2018, 33, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Walter, B. Anti-Müllerian hormone in dogs and cats reproduction. Reprod. Domest. Anim. 2020, 55, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Hollinshead, F.K.; Walker, C.; Hanlon, D.W. Determination of the normal reference interval for anti-Müllerian hormone (AMH) in bitches and use of AMH as a potential predictor of litter size. Reprod. Domest. Anim. 2017, 52, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, B.; Flock, U.; Leykam, C.; Otzdorff, C.; Simmet, K.; Hecht, W.; Kempker, L.; Aupperle-Lellbach, H.; Reese, S. Serum anti-Müllerian hormone concentration as a diagnostic tool to identify testicular tissue in canine disorders of sexual development. Domest. Anim. Endocrinol. 2022, 78, 106654. [Google Scholar] [CrossRef] [PubMed]
- Rota, A.; Vannozzi, I.; Marianelli, S.; Gavazza, A.; Lubas, G. Laboratory and clinical evaluation of a FEIA method for canine serum progesterone assay. Reprod. Domest. Anim. 2016, 51, 69–74. [Google Scholar] [CrossRef]
- Themmen, A.P.N.; Kalra, B.; Visser, J.A.; Kumar, A.; Savjani, G.; de Gier, J.; Jaques, S. The use of anti-Müllerian hormone as diagnostic for gonadectomy status in dogs. Theriogenology 2016, 86, 1467–1474. [Google Scholar] [CrossRef]
- Hornakova, L.; Vrbovska, T.; Pavl’ak, M.; Valencakova-Agyagosova, A.; Halo, M.; Hajurka, J. The evaluation of blood concentrations of testosterone, 17β-oestradiol and anti-Mu¨llerian hormone in dogs with cryptorchidism and testicular tumours. Pol. J. Vet. Sci. 2017, 20, 677–685. [Google Scholar]
- Fontbonne, A.; Malandain, E. Ovarian ultrasonography and follow-up of estrus in the bitch and queen. Waltham Focus. 2006, 16, 22–29. [Google Scholar]
- Bergeron, L.H.; Nykamp, S.G.; Brisson, B.A.; Madan, P.; Gartley, C.J. An evaluation of B-mode and color Doppler ultrasonography for detecting periovulatory events in the bitch. Theriogenology 2013, 79, 274–283. [Google Scholar] [CrossRef]
- Maenhoudt, C.; Santos, N.R.; Fontaine, E.; Mir, F.; Raynaud, K.; Navarro, C.; Fontbonne, A. Results of GnRH agonist implants in oestrous induction and oestrous suppression in bitches and queens. Reprod. Domest. Anim. 2012, 47, 393–397. [Google Scholar] [CrossRef] [Green Version]
- Concannon, P.W. Reproductive cycles of the domestic bitch. Anim. Reprod. Sci. 2011, 124, 200–210. [Google Scholar] [CrossRef]
- Hollinshead, F.; Hanlon, D. Normal progesterone profiles during estrus in the bitch: A prospective analysis of 1420 estrous cycles. Theriogenology 2019, 125, 37–42. [Google Scholar] [CrossRef]
- Bouchard, G.F.; Solorzano, N.; Concannon, P.W.; Youngquist, R.S.; Bierschwal, C.J. Determination of ovulation time in bitches based on teasing, vaginal cytology, and elisa for progesterone. Theriogenology 1991, 35, 603–611. [Google Scholar] [CrossRef]
- Turna Yilmaz, O.; Toydemir, T.S.; Kirsan, I.; Gunay Ucmak, Z.; Caliskan Karacam, E. Anti-Müllerian hormone as a diagnostic tool for ovarian remnant syndrome in bitches. Vet. Res. Commun. 2015, 39, 159–162. [Google Scholar] [CrossRef]
- Pir Yagci, I.; Peckan, M.; Polat, I.M.; Kalender, H.; Macun, H.C. Does serum anti-Müllerian hormone levels always discriminate presence of the ovaries in adult bitches? Comparison of two ELISA kits. Reprod. Domest. Anim. 2016, 51, 910–915. [Google Scholar] [CrossRef]
- Alkan, K.K.; Ceylan, A.; Alkan, H.; Ozen, D.; Bayraktarogly, A.G.; Kaymaz, M. Immunohistochemical and qPCR determination of the expression and serum level of anti-Müllerian hormone in pre-pubertal, intact and ovarian remnant syndrome detected bitches. Reprod. Domest. Anim. 2019, 54, 979–986. [Google Scholar] [CrossRef]
- Guzel, Y.; Aba, Y.A.; Yakin, K.; Oktem, O. Menstrual cycle characteristics of young females with occult primary ovarian insufficiency at initial diagnosis and one-year follow-up with serum amh level and antral follicle count. PLoS ONE. 2017, 12, e0188334. [Google Scholar] [CrossRef] [Green Version]
- Victoria, M.; Labrosse, J.; Krief, F.; Cédrin-Durnerin, I.; Comtet, M.; Grynberg, M. Anti Müllerian hormone: More than a biomarker of female reproductive function. J. Gynecol. Obstet. Hum. Reprod. 2019, 48, 19–24. [Google Scholar] [CrossRef]
- McGee, E.A.; Hsueh, A.J. Initial and cyclic recruitment of ovarian follicles. Endocr. Rev. 2000, 21, 200–214. [Google Scholar]
- Durlinger, A.L.; Gruijters, M.J.; Kramer, P.; Karels, B.; Kumar, T.R.; Matzuk, M.M.; Rose, U.M.; de Jong, F.H.; Uilenbroek, J.T.; Grootegoed, J.A.; et al. Anti-Müllerian hormone attenuates the effects of FSH on follicle development in the mouse ovary. Endocrinology. 2001, 142, 4891–4899. [Google Scholar] [CrossRef]
- McGee, E.A.; Smith, R.; Spears, N.; Nachtigal, M.W.; Ingraham, H.; Hsueh, A.J. Müllerian inhibitory substance induces growth of rat preantral ovarian follicles. Biol. Reprod. 2001, 64, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirobe, S.; He, W.W.; Gustafson, M.L.; MacLaughlin, D.T.; Donahoe, P.K. Müllerian inhibiting substance gene expression in the cycling rat ovary correlates with recruited or graafian follicle selection. Biol. Reprod. 1994, 50, 1238–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, S.S.; Mahaffey, M.B.; Miller, D.M.; Thompson, F.N.; Chakraborty, P.K. Ultrasonographic appearance of the ovaries of dogs during the follicular and luteal phase of the estrous cycle. Am. J. Vet. Res. 1992, 53, 209–215. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Day | Mean Number of Follicles | Mean Ovarian Area (cm2) |
|---|---|---|
| 8 | 7.25 ± 0.96 a | 0.74 ± 0.11 A |
| 10 | 3.50 ± 3.12 b | 0.76 ± 0.24 AB |
| 12 | 6.50 ± 6.36 b | 0.75 ± 0.23 A |
| 14 | 5.50 ± 4.95 b | 0.73 ± 0.24 A |
| 16 | 5.0 ± 3.0 ab | 0.80 ± 0.36 B |
| 18 | 4.0 ± 1.41 b | 0.83 ± 0.38 AB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chotimanukul, S.; Goericke-Pesch, S.; Suwimonteerabutr, J.; Singlor, J.; Sangkrachang, E.; Tummaruk, P.; Ponglowhapan, S. Serum Anti-Müllerian Hormone Levels and Estrous Monitoring of GnRH Agonist Deslorelin-Induced Estrus in Bitches: A Pilot Study. Animals 2023, 13, 258. https://doi.org/10.3390/ani13020258
Chotimanukul S, Goericke-Pesch S, Suwimonteerabutr J, Singlor J, Sangkrachang E, Tummaruk P, Ponglowhapan S. Serum Anti-Müllerian Hormone Levels and Estrous Monitoring of GnRH Agonist Deslorelin-Induced Estrus in Bitches: A Pilot Study. Animals. 2023; 13(2):258. https://doi.org/10.3390/ani13020258
Chicago/Turabian StyleChotimanukul, Sroisuda, Sandra Goericke-Pesch, Junpen Suwimonteerabutr, Jinda Singlor, Ekkaphot Sangkrachang, Padet Tummaruk, and Suppawiwat Ponglowhapan. 2023. "Serum Anti-Müllerian Hormone Levels and Estrous Monitoring of GnRH Agonist Deslorelin-Induced Estrus in Bitches: A Pilot Study" Animals 13, no. 2: 258. https://doi.org/10.3390/ani13020258