
Survey of Zoonotic Diarrheagenic Protist and Hepatitis E Virus in Wild Boar (Sus scrofa) of Portugal
 , ,
, ,  , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
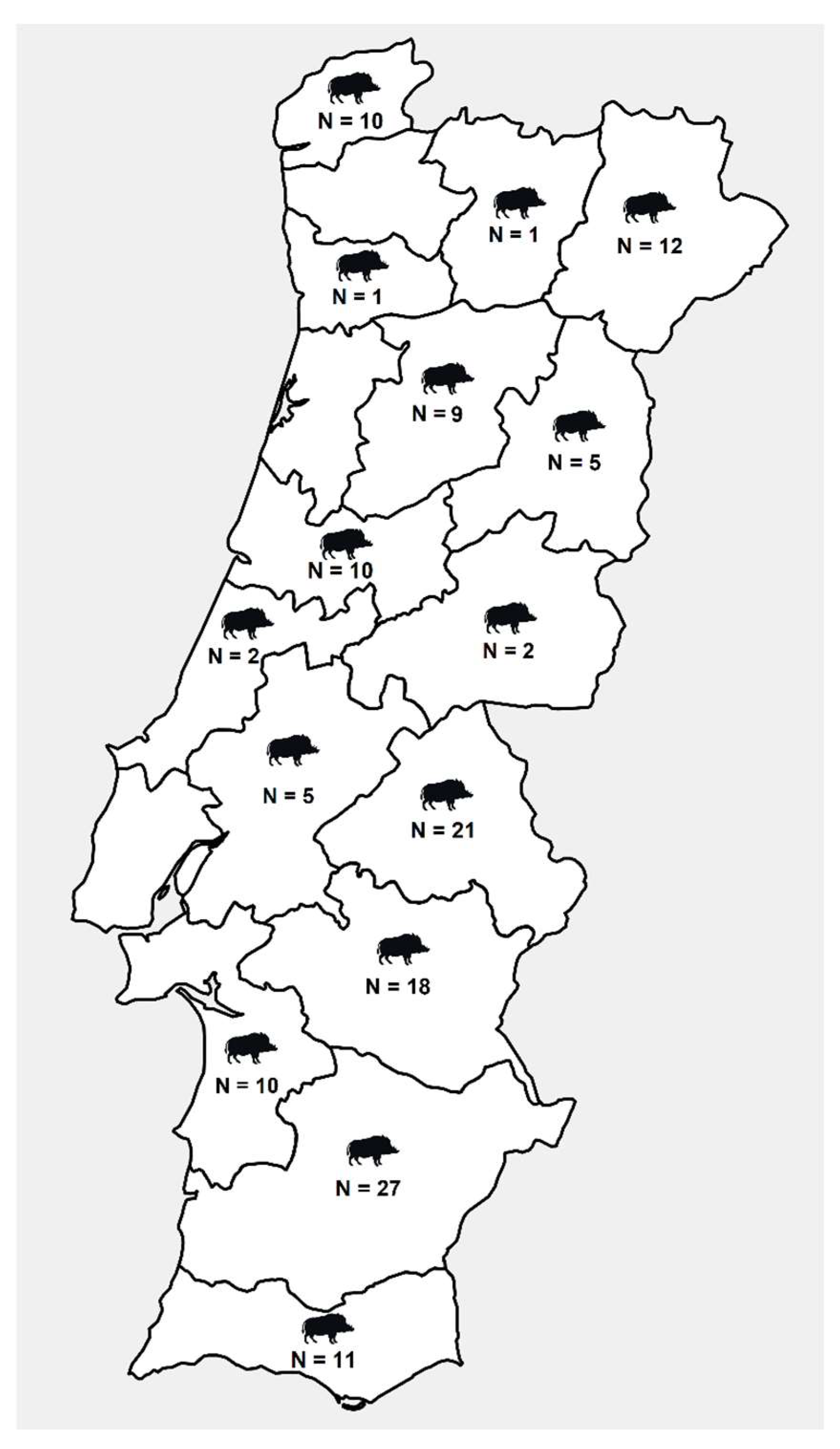
2.1. Sample Collection
2.2. DNA and RNA Extraction
2.3. Molecular Detection of Cryptosporidium spp.
2.4. Molecular Detection of Balantioides coli
2.5. Molecular Detection of Giardia duodenalis
2.6. Molecular Detection of Blastocystis sp.
2.7. Molecular Detection and Characterization of Enterocytozoon bieneusi
2.8. Molecular Detection of HEV
2.9. General Procedures
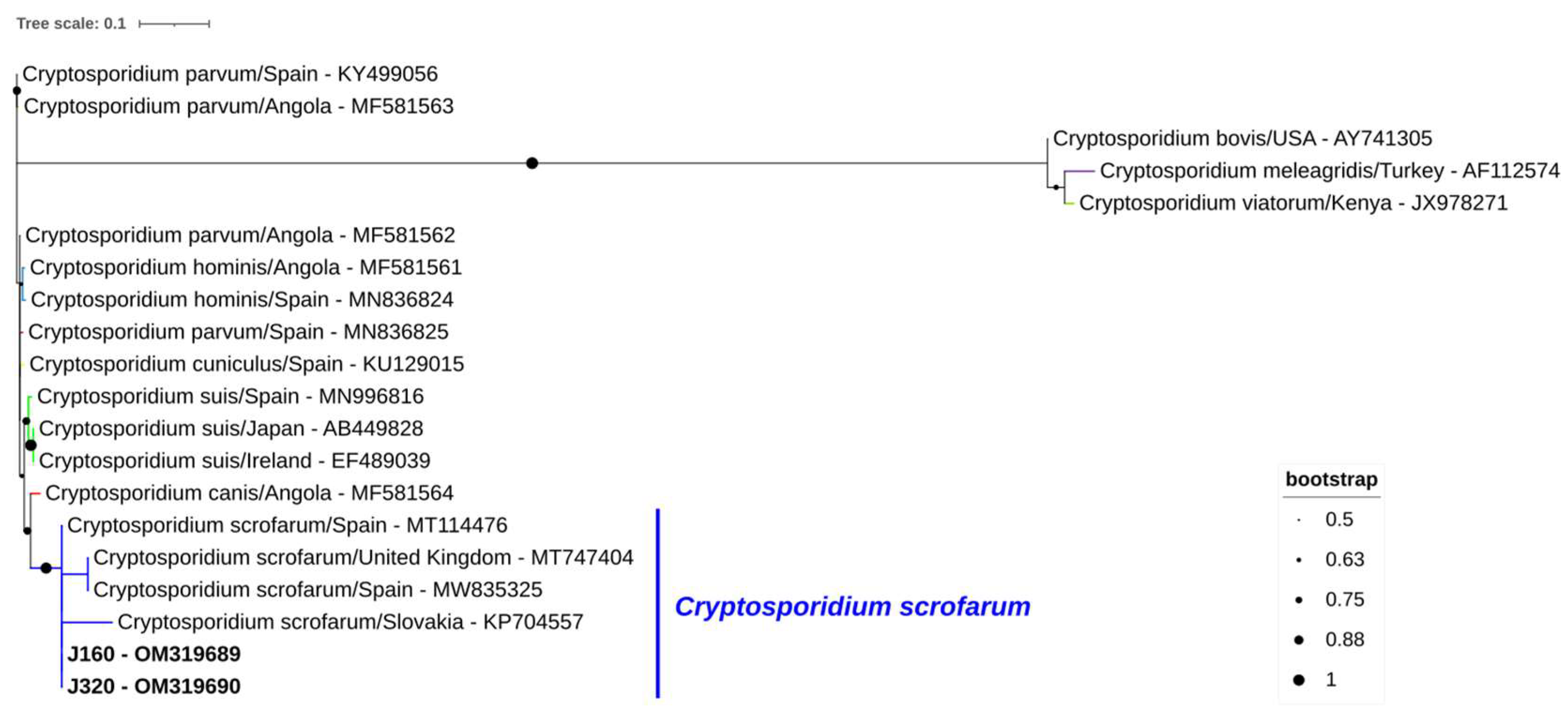
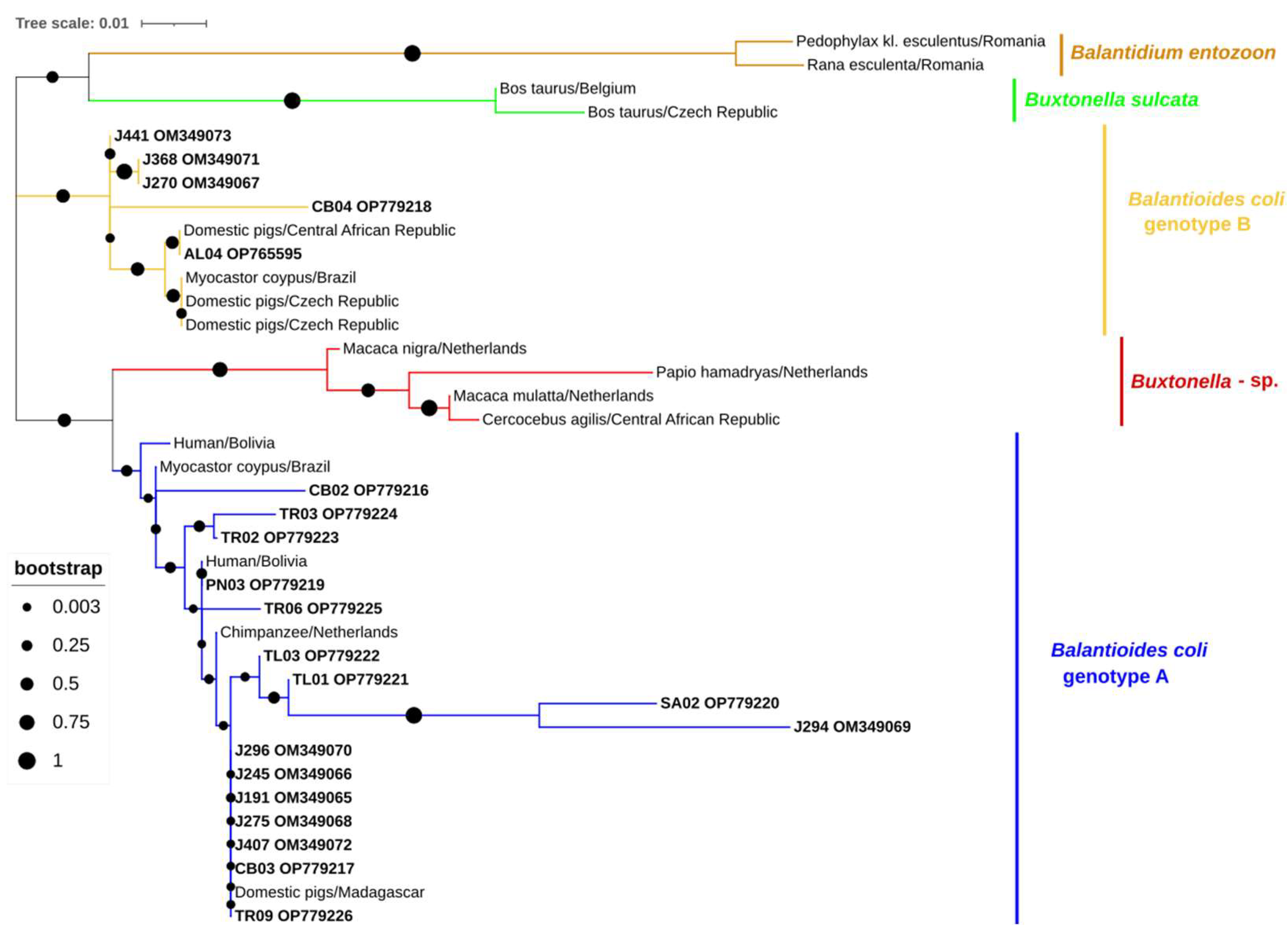
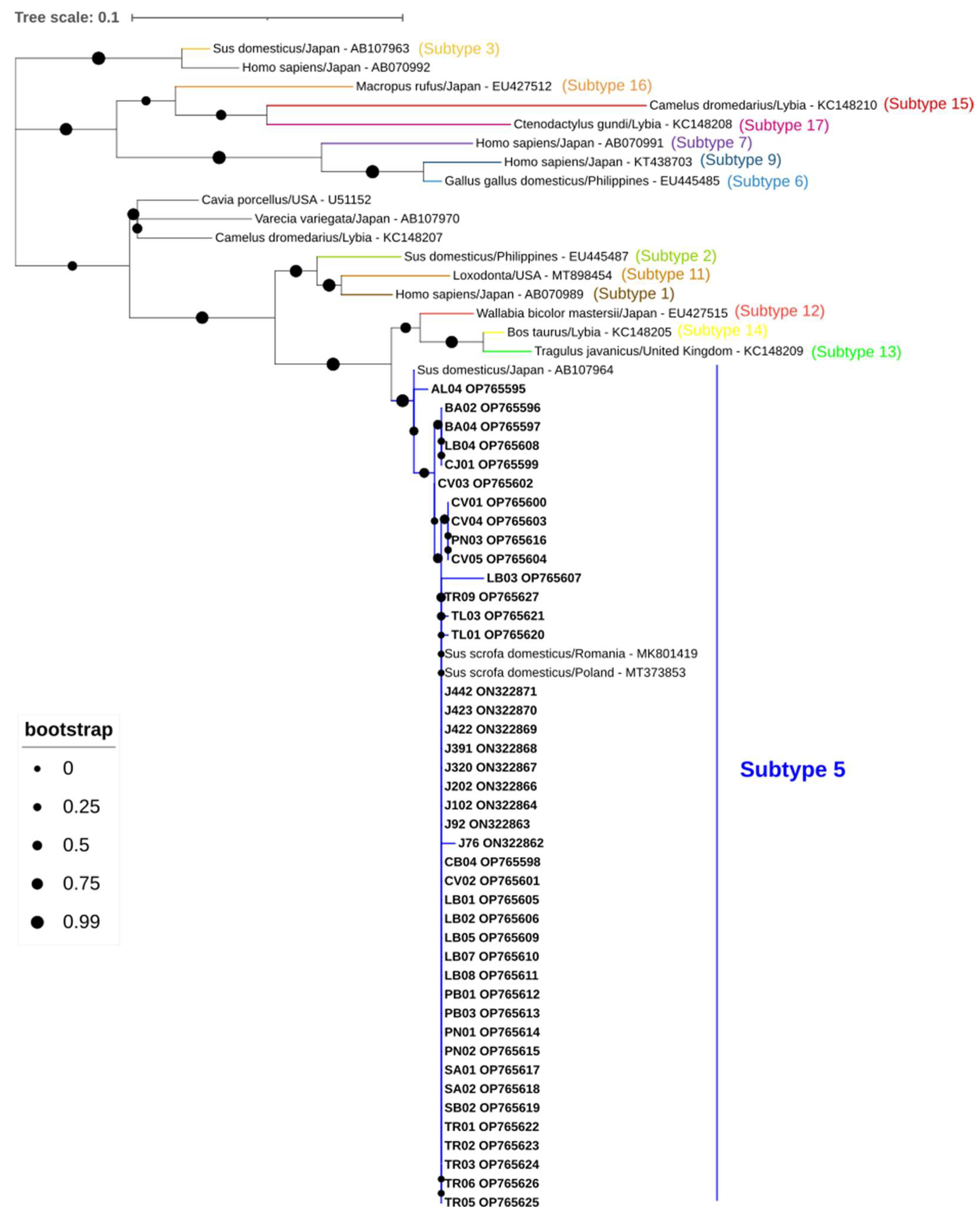
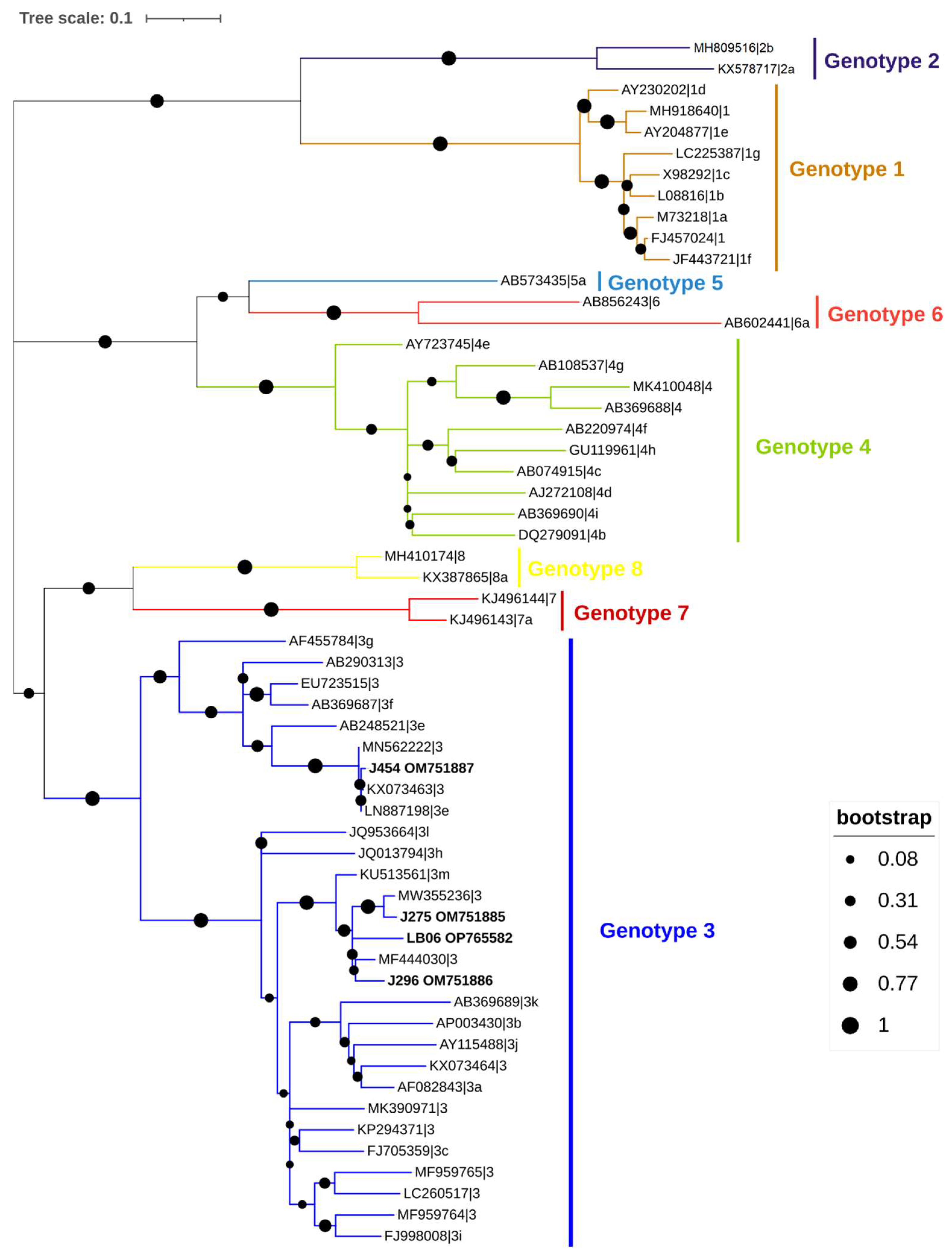
2.10. Sequencing and Phylogenetic Analysis
2.11. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rivero-Juarez, A.; Dashti, A.; López-López, P.; Muadica, A.S.; Risalde, M.D.L.A.; Risalde, M.D.L.A.; Köster, P.C.; Machuca, I.; Bailo, B.; De Mingo, M.H.; et al. Protist enteroparasites in wild boar (Sus scrofa ferus) and black Iberian pig (Sus scrofa domesticus) in southern Spain: A protective effect on hepatitis e acquisition? Parasit. Vectors. 2020, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- VanderWaal, K.; Deen, J. Global trends in infectious diseases of swine. Proc. Natl. Acad. Sci. USA 2018, 115, 11495–11500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dashti, A.; Rivero-Juarez, A.; Santín, M.; López-López, P.; Caballero-Gómez, J.; Frías-Casas, M.; Köster, P.C.; Bailo, B.; Calero-Bernal, R.; Briz, V.; et al. Enterocytozoon bieneusi (Microsporidia): Identification of novel genotypes and evidence of transmission between sympatric wild boars (Sus scrofa ferus) and Iberian pigs (Sus scrofa domesticus) in Southern Spain. Transbound. Emerg. Dis. 2020, 67, 2869–2880. [Google Scholar] [CrossRef] [PubMed]
- Lhotská, Z.; Jirků, M.; Hložková, O.; Brožová, K.; Jirsová, D.; Stensvold, C.R.; Kolísko, M.; Jirků Pomajbíková, K. A study on the prevalence and subtype diversity of the intestinal protist Blastocystis sp. in a gut-healthy human population in the Czech Republic. Front. Cell. Infect. Microbiol. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Feng, Y.; Xiao, L. Diagnosis and molecular typing of Enterocytozoon bieneusi: The significant role of domestic animals in transmission of human microsporidiosis. Res. Vet. Sci. 2020, 133, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Robertson, L.J.; Lalle, M.; Paulsen, P. Why we need a European focus on foodborne parasites. Exp. Parasitol. 2020, 214, 107900. [Google Scholar] [CrossRef]
- Schuster, F.L.; Ramirez-Avila, L. Current world status of Balantidium coli. Clin. Microbiol. Rev. 2008, 21, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Armson, A.; Yang, R.; Thompson, J.; Johnson, J.; Reid, S.; Ryan, U.M. Giardia genotypes in pigs in Western Australia: Prevalence and association with diarrhea. Exp. Parasitol. 2009, 121, 381–383. [Google Scholar] [CrossRef] [Green Version]
- Ponce-Gordo, F.; García-Rodríguez, J.J. Balantioides coli. Res. Vet. Sci. 2021, 135, 424–431. [Google Scholar] [CrossRef]
- Wang, P.; Li, S.; Zou, Y.; Du, Z.-C.; Song, D.-P.; Wang, P.; Chen, X.-Q. The infection and molecular characterization of Cryptosporidium spp. in diarrheic pigs in southern China. Microb. Pathog. 2022, 165, 105459. [Google Scholar] [CrossRef]
- Ballweber, L.R.; Xiao, L.; Bowman, D.D.; Kahn, G.; Cama, V.A. Giardiasis in dogs and cats: Update on epidemiology and public health significance. Trends. Parasitol. 2010, 26, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Ben Ayed, L.; Yang, W.; Widmer, G.; Cama, V.; Ortega, Y.; Xiao, L. Survey and genetic characterization of wastewater in Tunisia for Cryptosporidium spp.; Giardia duodenalis, Enterocytozoon bieneusi, Cyclospora cayetanensis and Eimeria spp. J. Water Health 2012, 10, 431–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cama, V.A.; Pearson, J.; Cabrera, L.; Pacheco, L.; Gilman, R.; Meyer, S.; Ortega, Y.; Xiao, L. Transmission of Enterocytozoon bieneusi between a child and guinea pigs. J. Clin. Microbiol. 2007, 45, 2708–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalmers, R.M.; Elwin, K.; Thomas, A.L.; Guy, E.C.; Mason, B. Long-term Cryptosporidium typing reveals the aetiology and species-specific epidemiology of human cryptosporidiosis in England and Wales, 2000 to 2003. Euro Surveill. 2009, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Wang, X.; Yang, R.; Zhao, W.; Li, N.; Guo, Y.; Xiao, L.; Feng, Y. Molecular characterization of the waterborne pathogens Cryptosporidium spp.; Giardia duodenalis, Enterocytozoon bieneusi, Cyclospora cayetanensis and Eimeria spp. in wastewater and sewage in Guangzhou, China. Parasit. Vectors 2021, 14, 1–10. [Google Scholar] [CrossRef]
- Galván, A.L.; Magnet, A.; Izquierdo, F.; Fenoy, S.; Rueda, C.; Adillo, C.F.V.; Henriques-Gil, N.; del Aguila, C. Molecular characterization of human-pathogenic microsporidia and Cyclospora cayetanensis isolated from various water sources in Spain: A year-long longitudinal study. Appl. Environ. Microbiol. 2013, 79, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Gumbo, T.; Sarbah, S.; Gangaidzo, I.T.; Ortega, Y.; Sterling, C.R.; Carville, A.; Tzipori, S.; Wiest, P.M. Intestinal parasites in patients with diarrhea and human immunodeficiency virus infection in Zimbabwe. Aids 1999, 13, 819–821. [Google Scholar] [CrossRef]
- Leelayoova, S.; Subrungruang, I.; Rangsin, R.; Chavalitshewinkoon-Petmitr, P.; Worapong, J.; Naaglor, T.; Mungthin, M. Transmission of Enterocytozoon bieneusi genotype a in a Thai orphanage. Am. J. Trop. Med. Hyg. 2005, 73, 104–107. [Google Scholar] [CrossRef]
- Michlmayr, D.; Alves de Sousa, L.; Müller, L.; Jokelainen, P.; Ethelberg, S.; Vestergaard, L.S.; Schjørring, S.; Mikkelsen, S.; Jensen, C.W.; Rasmussen, L.D.; et al. Incubation period, spore shedding duration, and symptoms of Enterocytozoon bieneusi genotype C infection in a foodborne outbreak in Denmark, 2020. Clin. Infect. Dis. 2021, 75, 468–475. [Google Scholar] [CrossRef]
- Sheikh, S.; Asghari, A.; Sadraei, J.; Pirestani, M.; Zare, M. Blastocystis sp. subtype 9: As the first reported subtype in patients with schizophrenia in Iran. SN Compr. Clin. Med. 2020, 2, 633–639. [Google Scholar] [CrossRef]
- Zhang, Y.; Koehler, A.V.; Wang, T.; Gasser, R.B. Enterocytozoon bieneusi of animals—With an ‘Australian twist’. Adv. Parasitol. 2021, 111, 1–73. [Google Scholar] [PubMed]
- Santin, M. Cryptosporidium and Giardia in ruminants. Vet. Clin. N. Am. Food. Anim. Pract. 2020, 36, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Schubnell, F.; von Ah, S.; Graage, R.; Sydler, T.; Sidler, X.; Hadorn, D.; Basso, W. Occurrence, clinical involvement and zoonotic potential of endoparasites infecting Swiss pigs. Parasitol. Int. 2016, 65, 618–624. [Google Scholar] [CrossRef]
- Ježková, J.; Limpouchová, Z.; Prediger, J.; Holubová, N.; Sak, B.; Konečný, R.; Květoňová, D.; Hlásková, L.; Rost, M.; McEvoy, J.; et al. Cryptosporidium myocastoris n. sp. (Apicomplexa: Cryptosporidiidae), the Species Adapted to the Nutria (Myocastor coypus). Microorganisms 2021, 9, 813. [Google Scholar] [CrossRef]
- Zahedi, A.; Bolland, S.J.; Oskam, C.L.; Ryan, U. Cryptosporidium abrahamseni n. sp. (Apicomplexa: Cryptosporidiiae) from red-eye tetra (Moenkhausia sanctaefilomenae). Exp. Parasitol. 2021, 223, 108089. [Google Scholar] [CrossRef]
- De Felice, L.A.; Moré, G.; Cappuccio, J.; Venturini, M.C.; Unzaga, J.M. Molecular characterization of Cryptosporidium spp. from domestic pigs in Argentina. Vet. Parasitol. Reg. Stud. Rep. 2020, 22, 100473. [Google Scholar] [CrossRef] [PubMed]
- Němejc, K.; Sak, B.; Květoňová, D.; Hanzal, V.; Janiszewski, P.; Forejtek, P.; Rajský, D.; Ravaszová, P.; McEvoy, J.; Kváč, M. Cryptosporidium suis and Cryptosporidium scrofarum in Eurasian wild boars (Sus scrofa) in Central Europe. Vet. Parasitol. 2013, 197, 504–508. [Google Scholar] [CrossRef] [Green Version]
- Petersen, H.H.; Jianmin, W.; Katakam, K.K.; Mejer, H.; Thamsborg, S.M.; Dalsgaard, A.; Olsen, A.; Enemark, H.L. Cryptosporidium and Giardia in Danish organic pig farms: Seasonal and age-related variation in prevalence, infection intensity and species/genotypes. Vet. Parasitol. 2015, 214, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Pettersson, E.; Ahola, H.; Frössling, J.; Wallgren, P.; Troell, K. Detection and molecular characterisation of Cryptosporidium spp. In Swedish pigs. Acta Vet. Scand. 2020, 62, 1–7. [Google Scholar] [CrossRef]
- Wang, W.; Gong, Q.L.; Zeng, A.; Li, M.H.; Zhao, Q.; Ni, H.B. Prevalence of Cryptosporidium in pigs in China: A systematic review and meta-analysis. Transbound. Emerg. Dis. 2020, 68, 1400–1413. [Google Scholar] [CrossRef]
- Solaymani-Mohammadi, S.; Petri, W.A. Zoonotic implications of the swine-transmitted protozoal infections. Vet. Parasitol. 2006, 140, 189–203. [Google Scholar] [CrossRef]
- Ponce-Gordo, F.; Jimenez-Ruiz, E.; Martínez-Díaz, R.A. Tentative identification of the species of Balantidium from ostriches (Struthio camelus) as Balantidium coli-like by analysis of polymorphic DNA. Vet. Parasitol. 2008, 157, 41–49. [Google Scholar] [CrossRef]
- Ryan, U.; Zahedi, A. Molecular epidemiology of giardiasis from a veterinary perspective. Adv. Parasitol. 2019, 106, 209–254. [Google Scholar] [PubMed]
- Sprong, H.; Cacciò, S.M.; Van Der Giessen, J.W.B. Identification of zoonotic genotypes of Giardia duodenalis. PLoS Negl. Trop. Dis. 2009, 3, e558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minetti, C.; Taweenan, W.; Hogg, R.; Featherstone, C.; Randle, N.; Latham, S.M.; Wastling, J.M. Occurrence and diversity of Giardia duodenalis assemblages in livestock in the UK. Transbound. Emerg. Dis. 2014, 61, e60–e67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaoyu, F.; Xiao, L. Zoonotic potential and molecular epidemiology of Giardia species and giardiasis. Clin. Microbiol. Rev. 2011, 24, 110–140. [Google Scholar]
- Sharifi, Y.; Abbasi, F.; Shahabi, S.; Zaraei, A.; Mikaeili, F.; Sarkari, B. Comparative genotyping of Blastocystis infecting cattle and human in the south of Iran. Comp. Immunol. Microbiol. Infect. Dis. 2020, 72, 101529. [Google Scholar] [CrossRef]
- Khademvatan, S.; Masjedizadeh, R.; Yousefi-Razin, E.; Mahbodfar, H.; Rahim, F.; Yousefi, E.; Foroutan, M. PCR-based molecular characterization of Blastocystis hominis subtypes in southwest of Iran. J. Infect. Public Health. 2018, 11, 43–47. [Google Scholar] [CrossRef]
- Maloney, J.G.; da Cunha, M.J.R.; Molokin, A.; Cury, M.C.; Santin, M. Next-generation sequencing reveals wide genetic diversity of Blastocystis subtypes in chickens including potentially zoonotic subtypes. Parasitol. Res. 2021, 120, 2219–2231. [Google Scholar] [CrossRef]
- Shams, M.; Shamsi, L.; Sadrebazzaz, A.; Asghari, A.; Badali, R.; Omidian, M.; Hassanipour, S. A systematic review and meta-analysis on the global prevalence and subtypes distribution of Blastocystis sp. infection in cattle: A zoonotic concern. Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101650. [Google Scholar] [CrossRef]
- Baek, S.; Maloney, J.G.; Molokin, A.; George, N.S.; Cortés Vecino, J.A.; Santin, M. Diversity of Blastocystis subtypes in horses in Colombia and identification of two new subtypes. Microorganisms 2022, 10, 1693. [Google Scholar] [CrossRef] [PubMed]
- Maloney, J.G.; Molokin, A.; da Cunha, M.J.R.; Cury, M.C.; Santin, M. Blastocystis subtype distribution in domestic and captive wild bird species from Brazil using next generation amplicon sequencing. Parasite Epidemiol. Control. 2020, 9, e00138. [Google Scholar] [CrossRef]
- Stensvold, C.R.; Clark, C.G. Pre-empting Pandora’s Box: Blastocystis subtypes revisited. Trends. Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, J.D.; Sánchez, A.; Hernández, C.; Flórez, C.; Bernal, M.C.; Giraldo, J.C.; Reyes, P.; López, M.C.; García, L.; Cooper, P.J.; et al. Geographic distribution of human Blastocystis subtypes in South America. Infect. Genet. Evol. 2016, 41, 32–35. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, S.; Dong, H.; Wang, R.; Zhang, L.; Ge, Y. Prevalence and molecular characteristics of Blastocystis sp. from Peafowl (Pavo cristatus) in China. J. Parasitol. 2021, 107, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Qiao, H.; Wang, H.; Li, S.; Zhai, P.; Huang, J.; Guo, Y. Molecular prevalence and subtypes of Blastocystis sp. in primates in northern China. Transbound. Emerg. Dis. 2020, 67, 2789–2796. [Google Scholar] [CrossRef]
- Jinatham, V.; Maxamhud, S.; Popluechai, S.; Tsaousis, A.D.; Gentekaki, E. Blastocystis One Health approach in a rural community of Northern Thailand: Prevalence, subtypes and novel transmission routes. Front. Microbiol. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Khaled, S.; Gantois, N.; Ly, A.T.; Senghor, S.; Even, G.; Dautel, E.; Dejager, R.; Sawant, M.; Baydoun, M.; Benamrouz-Vanneste, S.; et al. Prevalence and subtype distribution of Blastocystis sp. in senegalese school children. Microorganisms 2020, 8, 1408. [Google Scholar] [CrossRef]
- AbuOdeh, R.; Ezzedine, S.; Madkour, M.; Stensvold, C.R.; Samie, A.; Nasrallah, G.; AlAbsi, E.; ElBakri, A. Molecular subtyping of Blastocystis from diverse animals in the United Arab Emirates. Protist 2019, 170, 125679. [Google Scholar] [CrossRef] [PubMed]
- Asghari, A.; Sadraei, J.; Pirestani, M.; Mohammadpour, I. First molecular identification and subtype distribution of Blastocystis sp. isolated from hooded crows (Corvus cornix) and pigeons (Columba livia) in Tehran Province, Iran. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 25–30. [Google Scholar] [CrossRef]
- Mohammadpour, I.; Bozorg-Ghalati, F.; Gazzonis, A.L.; Manfredi, M.T.; Motazedian, M.H.; Mohammadpour, N. First molecular subtyping and phylogeny of Blastocystis sp. isolated from domestic and synanthropic animals (dogs, cats and brown rats) in southern Iran. Parasit. Vectors 2020, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Parkar, U.; Traub, R.J.; Vitali, S.; Elliot, A.; Levecke, B.; Robertson, I.; Geurden, T.; Steele, J.; Drake, B.; Thompson, R.C.A. Molecular characterization of Blastocystis isolates from zoo animals and their animal-keepers. Vet. Parasitol. 2010, 169, 8–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauff-Adedotun, A.A.; Mohd Zain, S.N.; Farah Haziqah, M.T. Current status of Blastocystis sp. in animals from Southeast Asia: A review. Parasitol. Res. 2020, 119, 3559–3570. [Google Scholar] [CrossRef] [PubMed]
- Santín, M.; Gómez-Muñoz, M.T.; Solano-Aguilar, G.; Fayer, R. Development of a new PCR protocol to detect and subtype Blastocystis spp. from humans and animals. Parasitol. Res. 2011, 109, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Kotler, D.P.; Orenstein, J.M. Clinical syndromes associated with microsporidiosis. Adv. Parasitol. 1998, 40, 321–349. [Google Scholar] [CrossRef]
- Li, W.; Feng, Y.; Xiao, L. Enterocytozoon bieneusi. Trends. Parasitol. 2022, 38, 95–96. [Google Scholar] [CrossRef]
- Li, W.; Xiao, L. Ecological and public health significance of Enterocytozoon bieneusi. One Health 2021, 12, 100209. [Google Scholar] [CrossRef]
- Li, W.; Feng, Y.; Santin, M. Host specificity of Enterocytozoon bieneusi and public health implications. Trends. Parasitol. 2019, 35, 436–451. [Google Scholar] [CrossRef]
- Zhou, H.H.; Zheng, X.L.; Ma, T.M.; Qi, M.; Zhou, J.G.; Liu, H.J.; Lu, G.; Zhao, W. Molecular detection of Enterocytozoon bieneusi in farm-raised pigs in Hainan Province, China: Infection rates, genotype distributions, and zoonotic potential. Parasite 2020, 27, 12. [Google Scholar] [CrossRef] [Green Version]
- Raji, Y.E.; Toung, O.P.; Taib, N.M.; Sekawi, Z.B. Hepatitis E Virus: An emerging enigmatic and underestimated pathogen. Saudi J. Biol. Sci. 2022, 1, 499–512. [Google Scholar] [CrossRef]
- Hoofnagle, J.H.; Nelson, K.E.; Purcell, R.H. Hepatitis E. N. Engl. J. Med. 2012, 367, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Purdy, M.A.; Drexler, J.F.; Meng, X.-J.; Norder, H.; Okamoto, H.; Van der Poel, W.H.M.; Reuter, G.; de Souza, W.M.; Ulrich, R.G.; Smith, D.B. ICTV Virus taxonomy profile: Hepeviridae 2022. J. Gen. Virol. 2022, 103, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Scotto, G.; Bulla, F.; Campanale, F.; Tartaglia, A.; Fazio, V. Hepatitis E. Infez. Med. 2013, 21, 175–188. [Google Scholar] [PubMed]
- Martino, C.; Rampacci, E.; Pierini, I.; Giammarioli, M.; Stefanetti, V.; Hyatt, D.R.; Ianni, A.; Di Paolo, G.; Coletti, M.; Passamonti, F. Detection of anti-HEV antibodies and RNA of HEV in pigs from a hyperendemic Italian region with high human seroprevalence. Eur. J. Public Health. 2021, 31, 68–72. [Google Scholar] [CrossRef]
- Spahr, C.; Knauf-Witzens, T.; Vahlenkamp, T.; Ulrich, R.G.; Johne, R. Hepatitis E virus and related viruses in wild, domestic and zoo animals: A review. Zoonoses Public Health 2018, 65, 11–29. [Google Scholar] [CrossRef]
- Meng, X.J.; Lindsay, D.S. Wild boars as sources for infectious diseases in livestock and humans. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2697–2707. [Google Scholar] [CrossRef] [Green Version]
- Salines, M.; Andraud, M.; Rose, N. From the epidemiology of hepatitis e virus (HEV) within the swine reservoir to public health risk mitigation strategies: A comprehensive review. Vet. Res. 2017, 48, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Meng, X.J. Hepatitis E virus: Host tropism and zoonotic infection. Curr. Opin. Microbiol. 2021, 59, 8–15. [Google Scholar] [CrossRef]
- Subissi, L.; Peeters, M.; Lamoral, S.; Klamer, S.; Suin, V.; Van Gucht, S. Subtype-specific differences in the risk of hospitalisation among patients infected with hepatitis E virus genotype 3 in Belgium, 2010–2018. Epidemiol. Infect. 2019, 147, 5–8. [Google Scholar] [CrossRef]
- Berto, A.; Mesquita, J.R.; Hakze-van der Honing, R.; Nascimento, M.S.J.; van der Poel, W.H.M. Detection and characterization of Hepatitis E Virus in domestic pigs of different ages in Portugal. Zoonoses Public Health 2012, 59, 477–481. [Google Scholar] [CrossRef]
- Mesquita, J.R.; Oliveira, R.M.S.; Coelho, C.; Vieira-Pinto, M.; Nascimento, M.S.J. Hepatitis E virus in sylvatic and captive wild boar from Portugal. Transbound. Emerg. Dis. 2016, 63, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, M.S.J.; Pereira, S.S.; Teixeira, J.; Abreu-Silva, J.; Oliveira, R.M.S.; Myrmel, M.; Stene-Johansen, K.; Øverbø, J.; Gonçalves, G.; Mesquita, J.R. A nationwide serosurvey of hepatitis e virus antibodies in the general population of Portugal. Eur. J. Public Health. 2018, 28, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Dashti, A.; Rivero-Juárez, A.; Santín, M.; George, N.S.; Köster, P.C.; López-López, P.; Risalde, M.A.; García-Bocanegra, I.; Gómez-Villamandos, J.C.; Caballero-Gómez, J.; et al. Diarrhoea-causing enteric protist species in intensively and extensively raised pigs (Sus scrofa domesticus) in Southern Spain. Part I: Prevalence and genetic diversity. Transbound. Emerg. Dis. 2022, 69, e1051–e1064. [Google Scholar] [CrossRef]
- Bilenko, N.; Levy, A.; Dagan, R.; Deckelbaum, R.J.; El-On, Y.; Fraser, D. Does co-infection with Giardia lamblia modulate the clinical characteristics of enteric infections in young children? Eur. J. Epidemiol. 2004, 19, 877–883. [Google Scholar] [CrossRef]
- Rivero-Juárez, A.; Dashti, A.; Santín, M.; Köster, P.C.; López-López, P.; Risalde, M.A.; García-Bocanegra, I.; Gómez-Villamandos, J.C.; Caballero-Gómez, J.; Frías, M.; et al. Diarrhoea-causing enteric protist species in intensively and extensively raised pigs (Sus scrofa domesticus) in Southern Spain. Part II: Association with Hepatitis E virus susceptibility. Transbound. Emerg. Dis. 2022, 69, e1172–e1178. [Google Scholar] [CrossRef] [PubMed]
- Tiangtip, R.; Jongwutiwes, S. Molecular analysis of Cryptosporidium species isolated from HIV-infected patients in Thailand. Trop. Med. Int. Health. 2002, 7, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Ponce-Gordo, F.; Fonseca-Salamanca, F.; Martínez-Díaz, R.A. Genetic heterogeneity in internal transcribed spacer genes of Balantidium coli (Litostomatea, Ciliophora). Protist 2011, 162, 774–794. [Google Scholar] [CrossRef]
- Verweij, J.J.; Schinkel, J.; Laeijendecker, D.; Van Rooyen, M.A.A.; Van Lieshout, L.; Polderman, A.M. Real-time PCR for the detection of Giardia lamblia. Mol. Cell. Probes. 2003, 17, 223–225. [Google Scholar] [CrossRef]
- Scicluna, S.M.; Tawari, B.; Clark, C.G. DNA barcoding of Blastocystis. Protist 2006, 157, 77–85. [Google Scholar] [CrossRef]
- Buckholt, M.A.; Lee, J.H.; Tzipori, S. Prevalence of Enterocytozoon bieneusi in swine: An 18-month survey at a slaughterhouse in Massachusetts. Appl. Environ. Microbiol. 2002, 68, 2595–2599. [Google Scholar] [CrossRef] [Green Version]
- Johne, R.; Plenge-Bönig, A.; Hess, M.; Ulrich, R.G.; Reetz, J.; Schielke, A. Detection of a novel hepatitis E-like virus in faeces of wild rats using a nested broad-spectrum RT-PCR. J. Gen. Virol. 2010, 91, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [PubMed] [Green Version]
- Gomes-de-Sá, S.; Santos-Silva, S.; Moreira, A.S.; Barradas, P.F.; Amorim, I.; Cardoso, L.; Mesquita, J.R. Assessment of the circulation of Dirofilaria immitis in dogs from northern Portugal through combined analysis of antigens, DNA and parasite forms in blood. Acta Trop. 2022; 106799, in Press. [Google Scholar]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors-occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef]
- Mega, J.; Santos-Silva, S.; Loureiro, A.; Palmeira, J.D.; Torres, R.T.; Rivero-Juarez, A.; Carmena, D.; Mesquita, J. Balantioides coli fecal excretion in hunted wild cervids (Cervus elaphus and Dama dama) from Portugal. Pathogens 2022, 11, 1242. [Google Scholar] [CrossRef]
- Russini, V.; Di Filippo, M.M.; Fanelli, R.; Polidori, M.; Berrilli, F.; Di Cave, D.; Novelletto, A.; Calderini, P. Characterization of prevalence and genetic subtypes of Blastocystis sp. in wild and domestic Suidae of central Italy aided by amplicon NGS. Vet. Parasitol. Reg. Stud. Rep. 2020, 22, 100472. [Google Scholar] [CrossRef]
- Beck, R.; Sprong, H.; Lucinger, S.; Pozio, E.; Cacciò, S.M. A large survey of Croatian wild mammals for Giardia duodenalis reveals a low prevalence and limited zoonotic potential. Vector Borne Zoonotic Dis. 2011, 11, 1049–1055. [Google Scholar] [CrossRef]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- Casares-Jimenez, M.; Lopez-Lopez, P.; Caballero-Gomez, J.; Frias, M.; Perez-Hernando, B.; Oluremi, A.S.; Risalde, M.A.; Ruiz-Caceres, I.; Opaleye, O.O.; Garcia-Bocanegra, I.; et al. Global molecular diversity of Hepatitis E virus in wild boar and domestic pig. One Health 2021, 13, 100304. [Google Scholar] [CrossRef]
- Lapa, D.; Capobianchi, M.R.; Garbuglia, A.R. Epidemiology of hepatitis E virus in European countries. Int. J. Mol. Sci. 2015, 16, 25711–25743. [Google Scholar] [CrossRef] [PubMed]
- Caballero-Gómez, J.; Jiménez-Ruiz, S.; Lopez-Lopez, P.; Vicente, J.; Risalde, M.A.; Cano-Terriza, D.; Frias, M.; Barasona, J.A.; Rivero, A.; García-Bocanegra, I.; et al. Emergent subtype of hepatitis E virus genotype 3 in wild boar in Spain. Transbound. Emerg. Dis. 2019, 66, 1803–1808. [Google Scholar] [CrossRef] [PubMed]
- Cancela, F.; Panzera, Y.; Mainardi, V.; Gerona, S.; Ramos, N.; Pérez, R.; Arbiza, J.; Mirazo, S. Complete genome sequence of Hepatitis E Virus Genotype 3 obtained from a chronically infected individual in Uruguay. Microbiol. Resour. Announc. 2021, 10, 9–11. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cryptosporidium spp. | Balantioides coli | Giardia duodenalis | Blastocystis spp. | Enterocytozoon bieneusi | Hepatitis E Virus | |
|---|---|---|---|---|---|---|
| Positive samples | 2 | 21 | 0 | 42 | 0 | 4 |
| Prevalence | 2/144 (1.4%) | 21/144 (14.6%) | 0/144 (0%) | 42/144 (29.2%) | 0/144 (0%) | 4/144 (2.8%) |
| Fecal Presence | No. of Samples |
|---|---|
| Balantioides coli + Blastocystis spp. | 10 |
| Balantioides coli + Hepatitis E virus | 2 |
| Blastocystis spp. + Cryptosporidium spp. | 1 |
| Blastocystis spp. | 42 |
| Balantioides coli | 21 |
| Hepatitis E virus | 4 |
| Cryptosporidium spp. | 2 |
| Giardia duodenalis | 0 |
| Enterocytozoon bieneusi | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos-Silva, S.; Moraes, D.F.d.S.D.; López-López, P.; Palmeira, J.D.; Torres, R.T.; São José Nascimento, M.; Dashti, A.; Carmena, D.; Rivero-Juarez, A.; Mesquita, J.R. Survey of Zoonotic Diarrheagenic Protist and Hepatitis E Virus in Wild Boar (Sus scrofa) of Portugal. Animals 2023, 13, 256. https://doi.org/10.3390/ani13020256
Santos-Silva S, Moraes DFdSD, López-López P, Palmeira JD, Torres RT, São José Nascimento M, Dashti A, Carmena D, Rivero-Juarez A, Mesquita JR. Survey of Zoonotic Diarrheagenic Protist and Hepatitis E Virus in Wild Boar (Sus scrofa) of Portugal. Animals. 2023; 13(2):256. https://doi.org/10.3390/ani13020256
Chicago/Turabian StyleSantos-Silva, Sérgio, Danny Franciele da Silva Dias Moraes, Pedro López-López, Josman D. Palmeira, Rita T. Torres, Maria São José Nascimento, Alejandro Dashti, David Carmena, Antonio Rivero-Juarez, and João R. Mesquita. 2023. "Survey of Zoonotic Diarrheagenic Protist and Hepatitis E Virus in Wild Boar (Sus scrofa) of Portugal" Animals 13, no. 2: 256. https://doi.org/10.3390/ani13020256