

Effect of Sugar Beet Pulp and Anionic Salts on Metabolic Status and Mineral Homeostasis during the Peri-Parturient Period of Dairy Sheep
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Feeding Regimes
2.2. Blood Sampling
2.3. Urine Sampling
2.4. Biochemical Analyses
2.5. Ethical Statement
2.6. Statistical Analysis
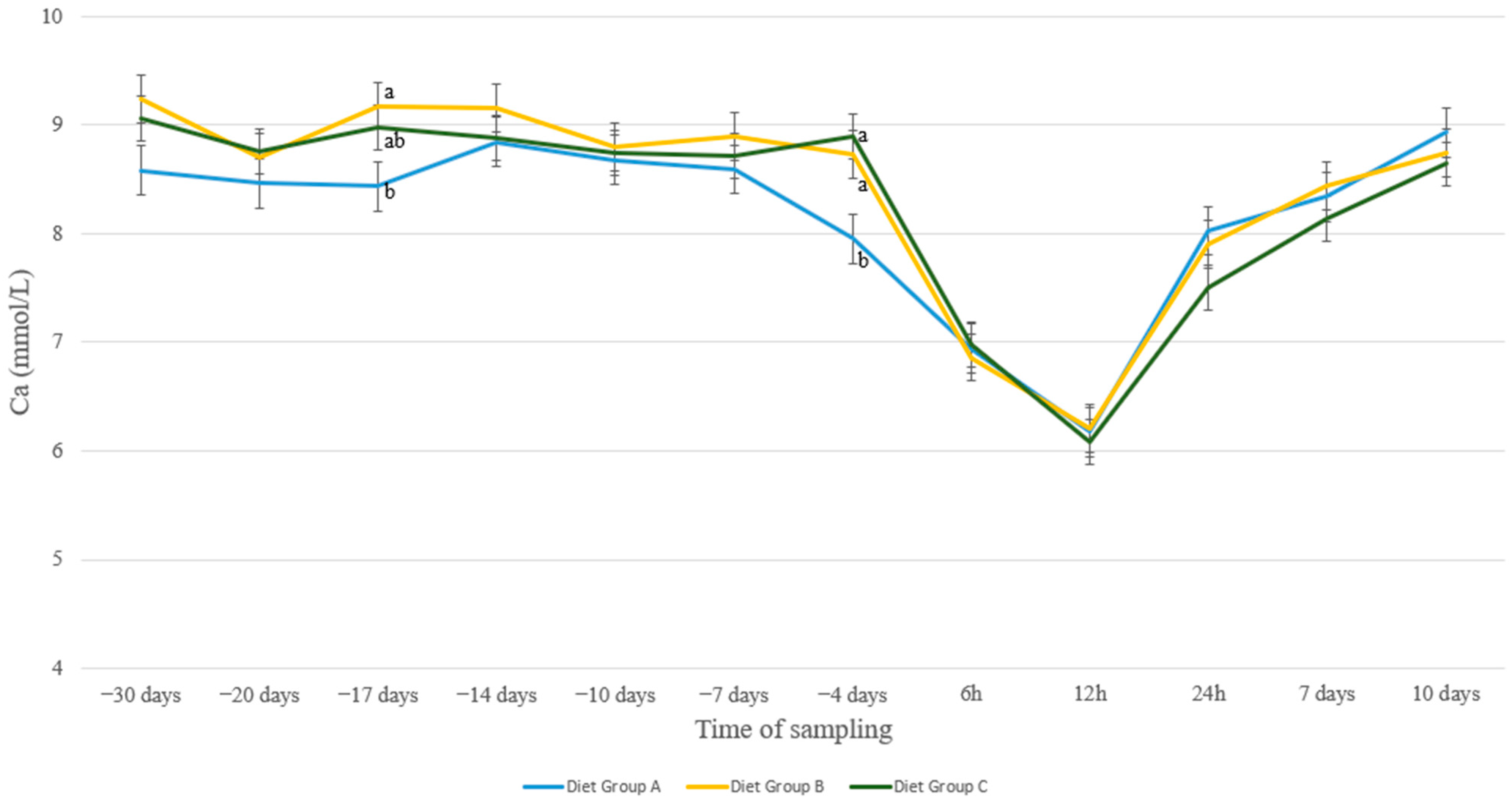
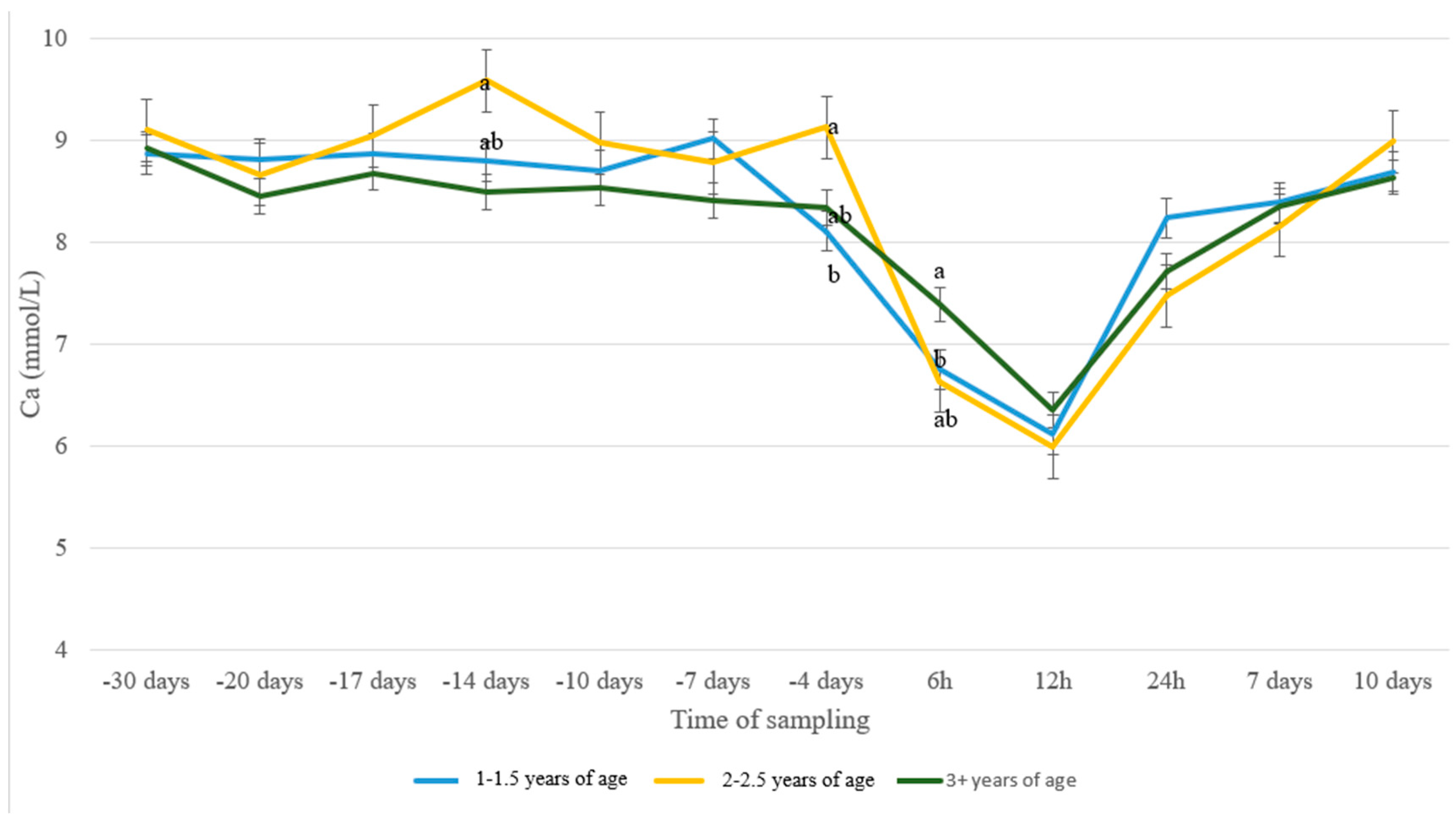
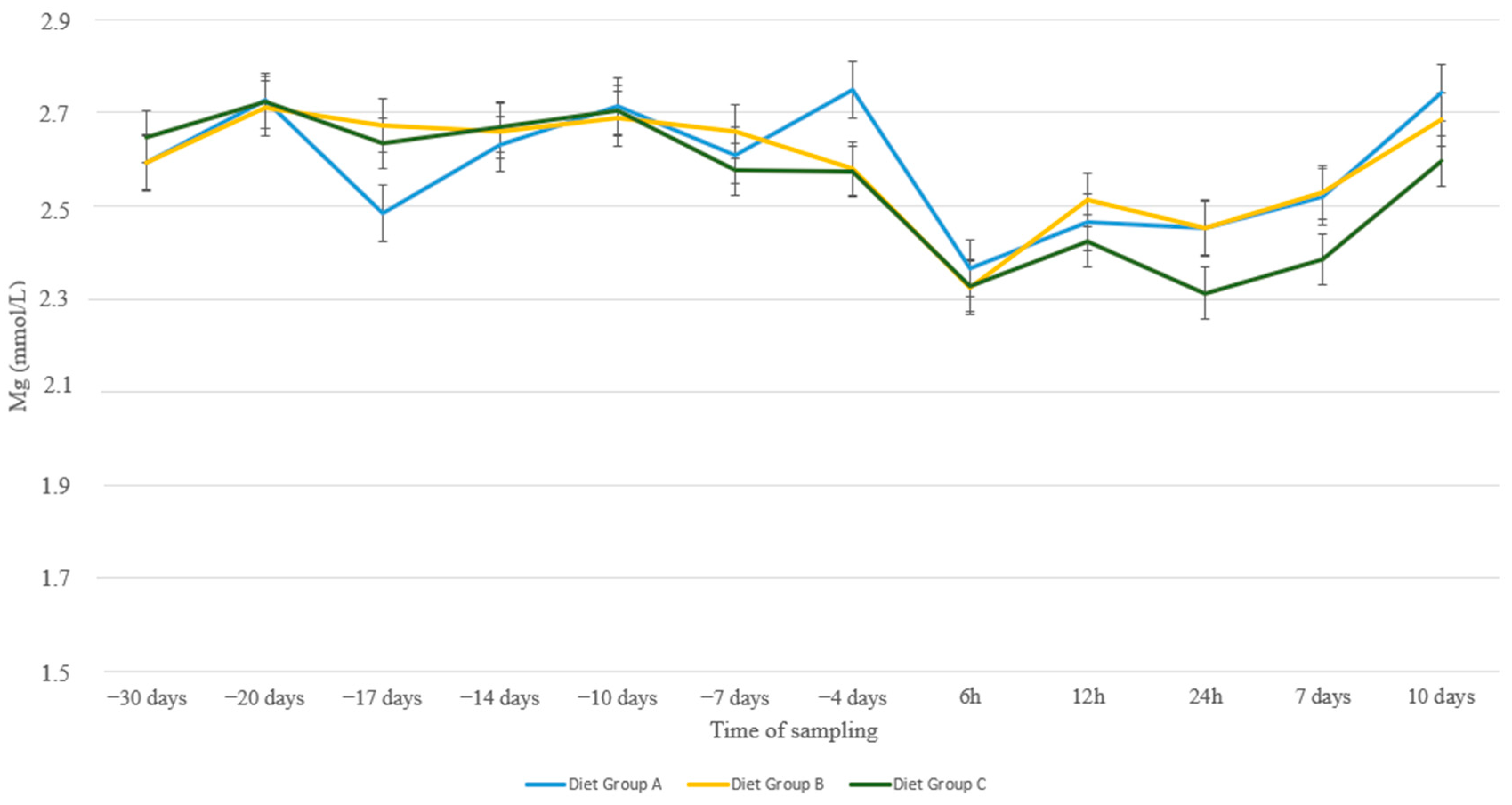
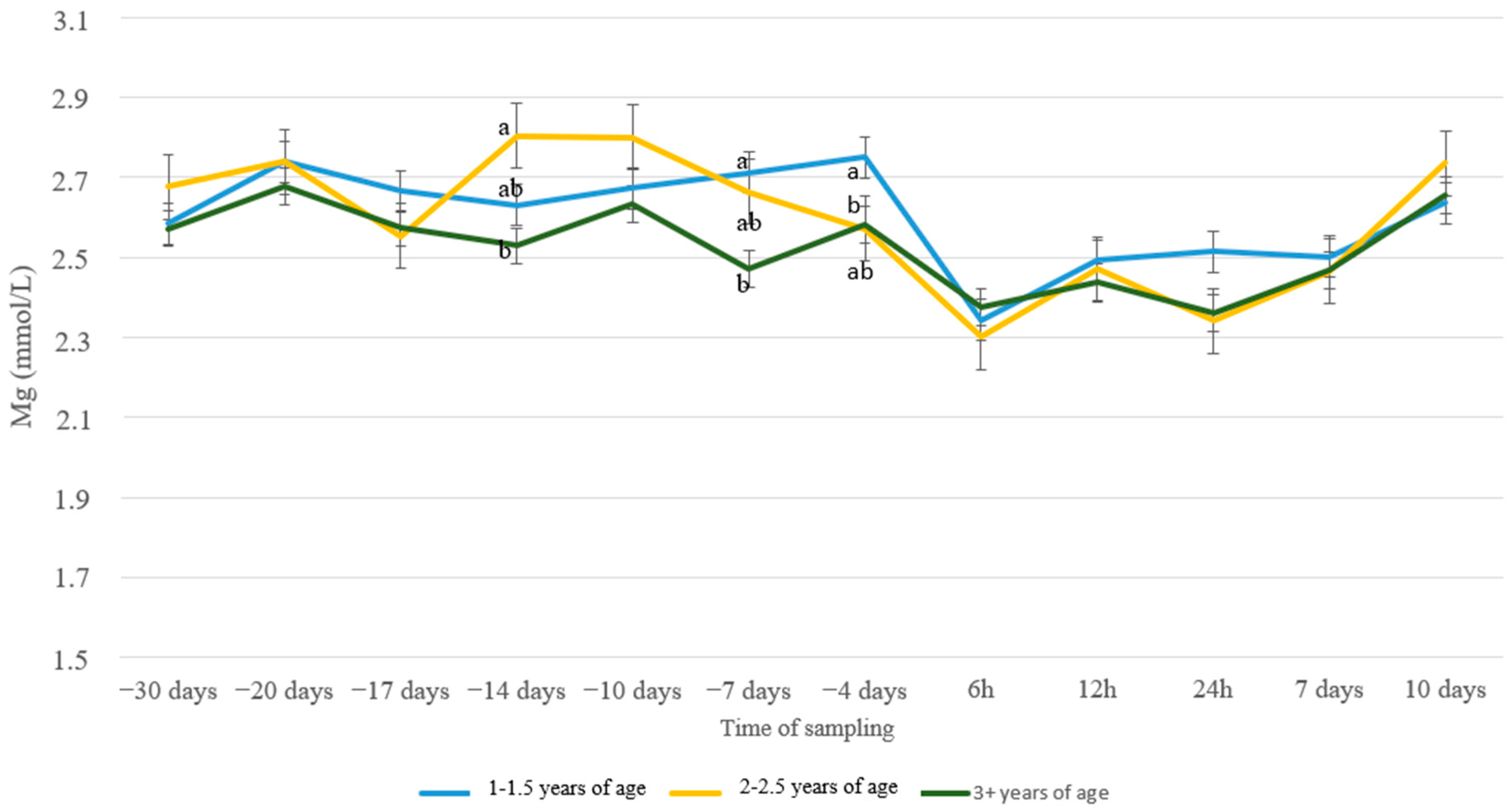
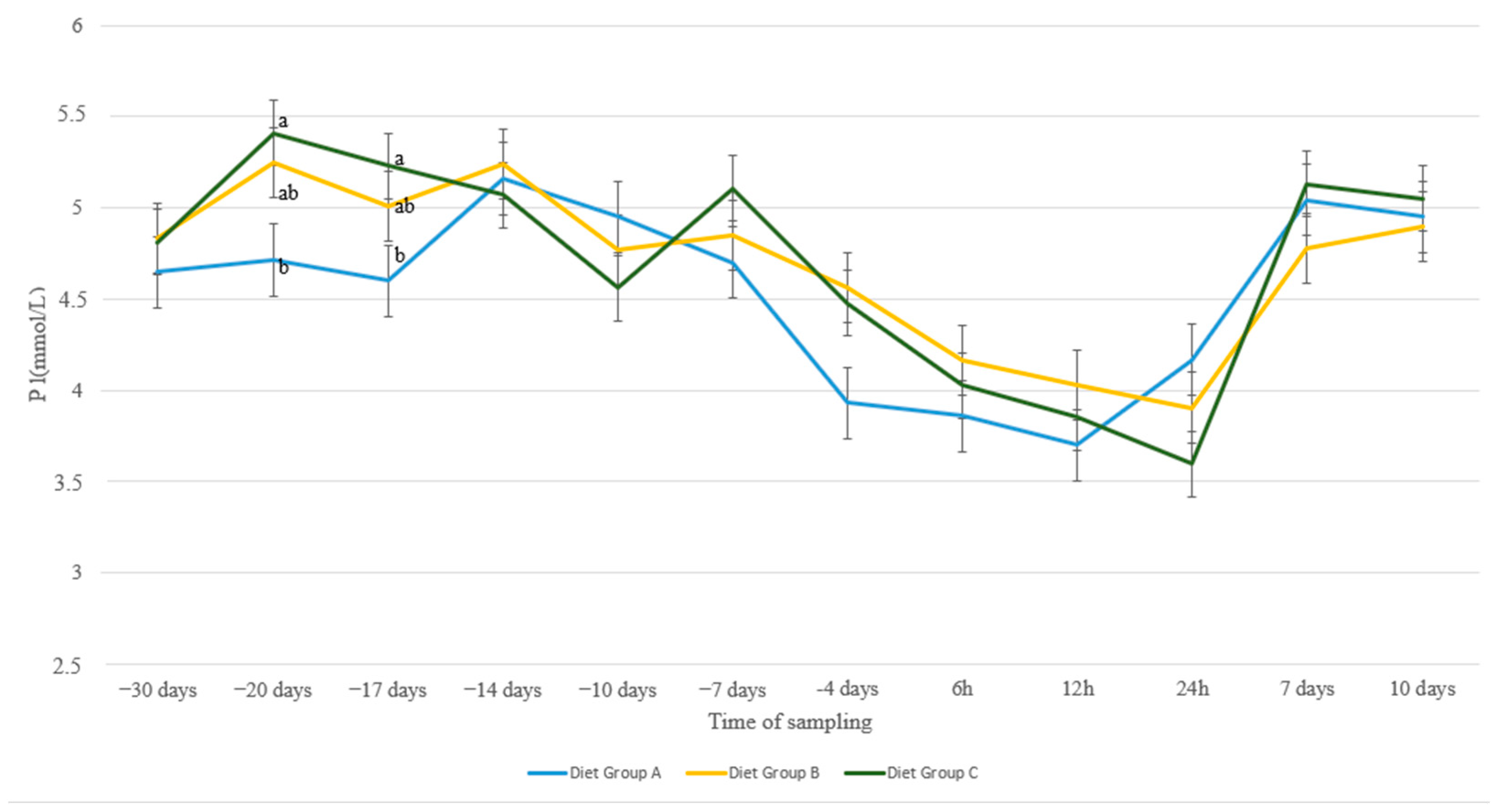
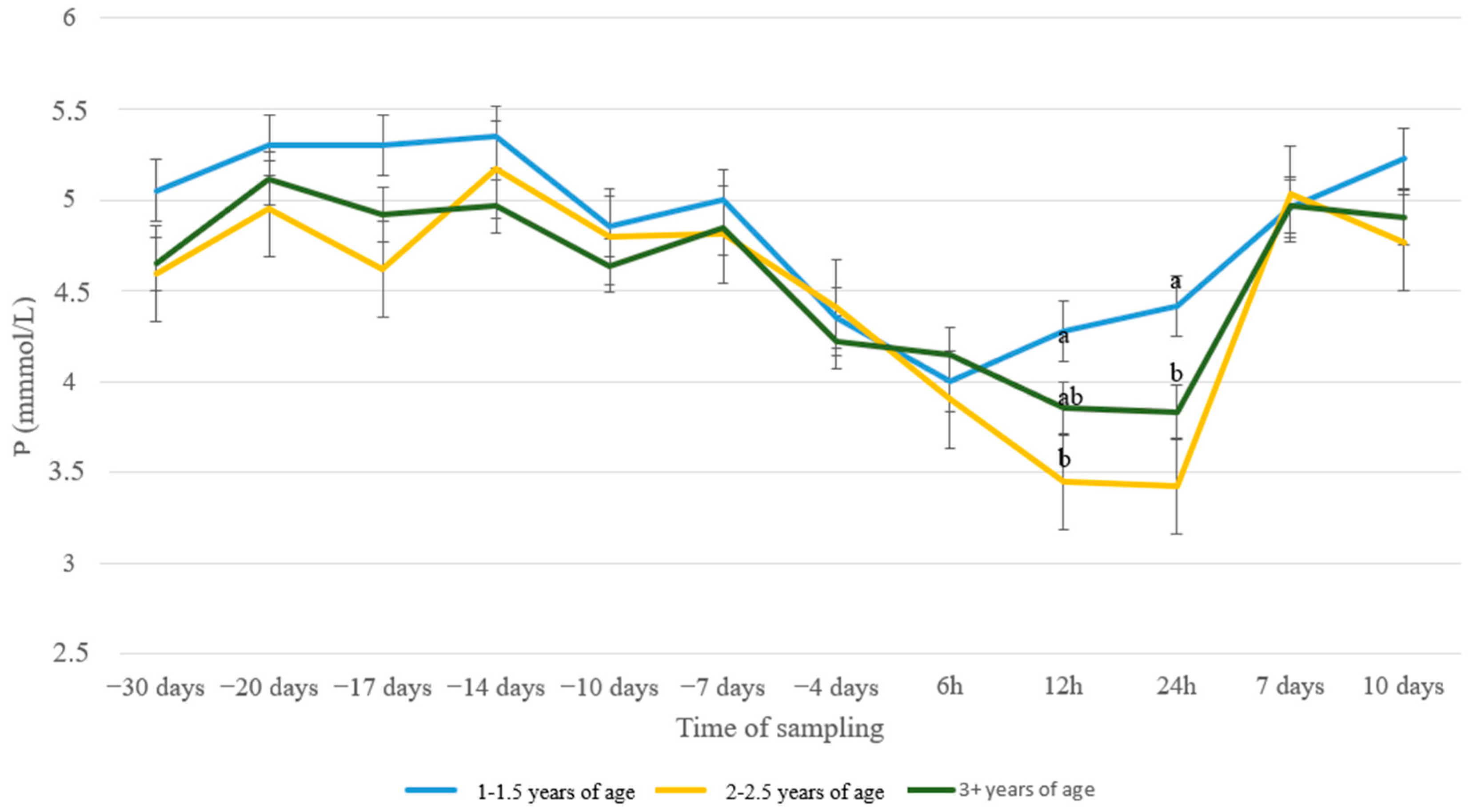
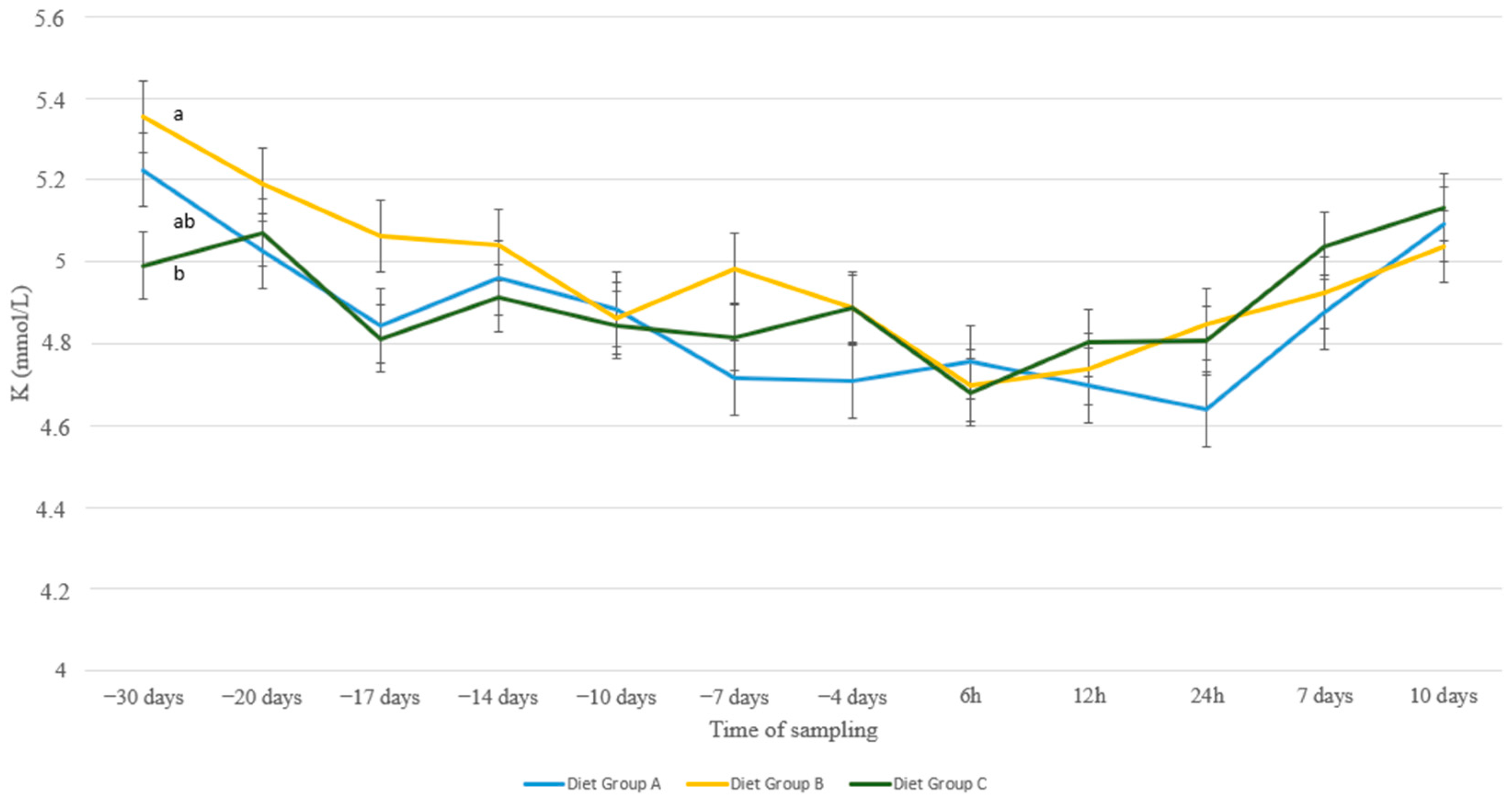
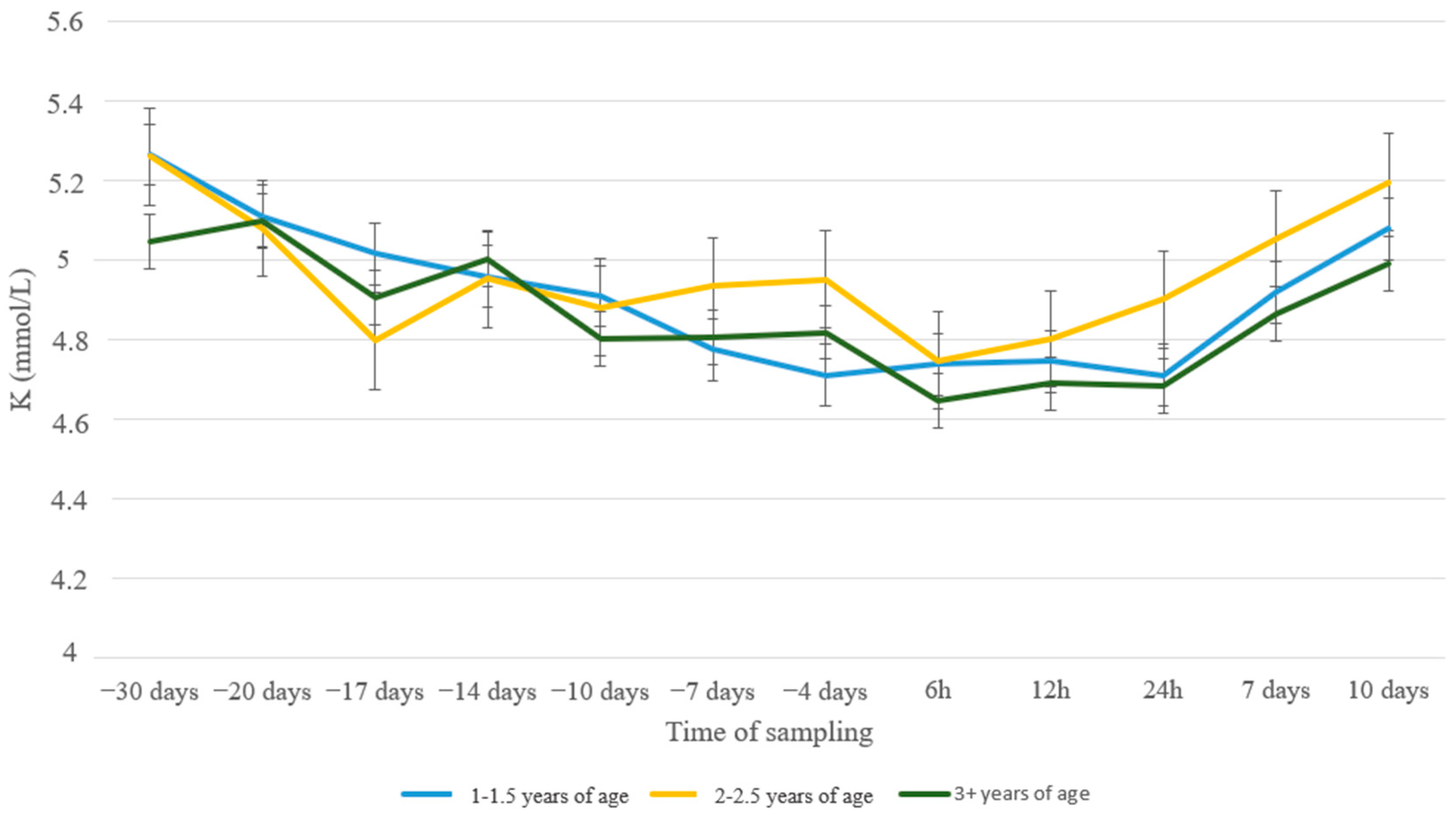
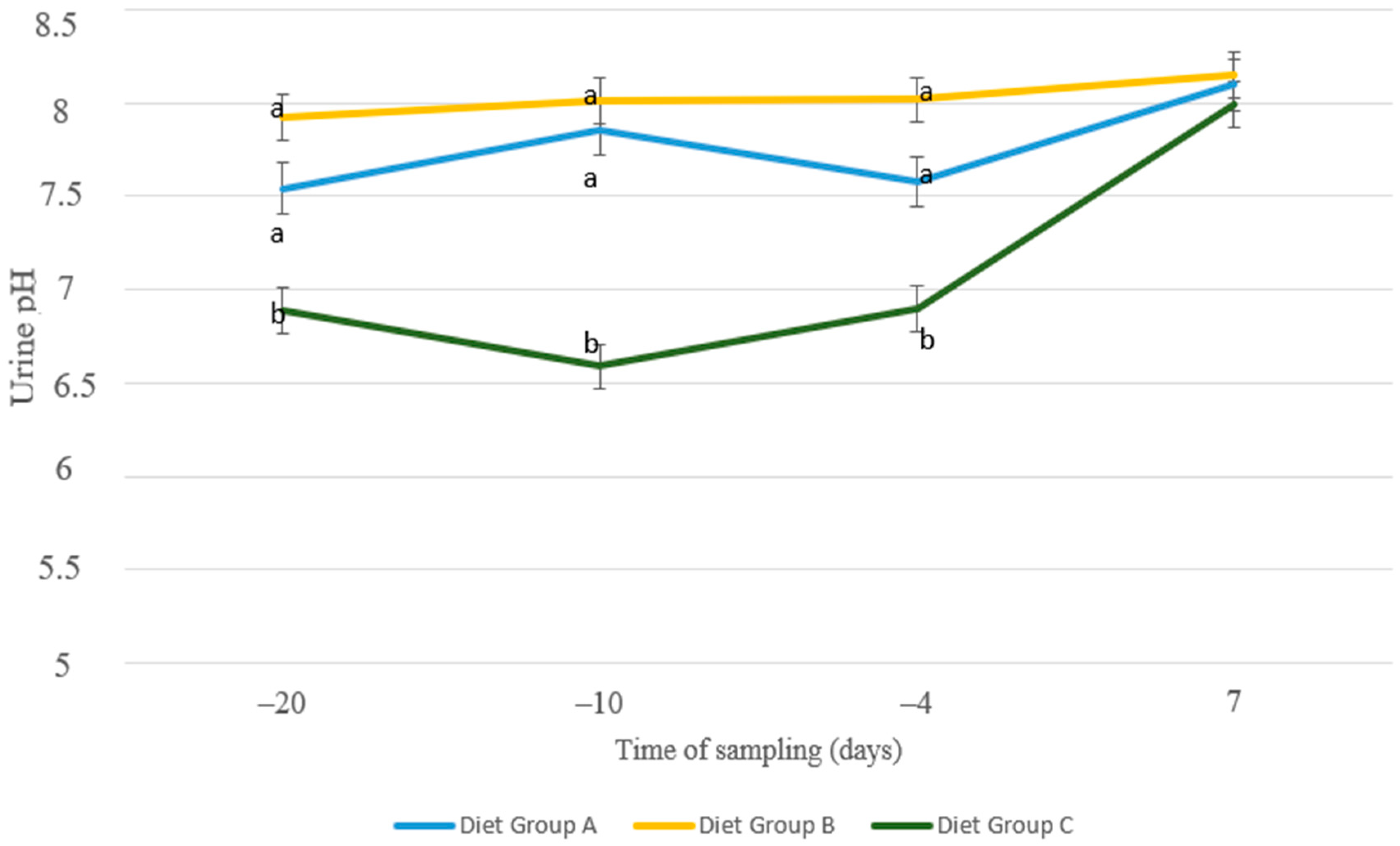
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci. 1995, 73, 2804. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, A.; Balikci, E.; Gurdogan, F. Serum Mineral Levels at Pregnancy and Postpartum in Single and Twin Pregnant Sheep. Biol. Trace Element Res. 2005, 107, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Ospina, P.A.; Nydam, D.V.; Stokol, T.; Overton, T.R. Association between the proportion of sampled transition cows with increased nonesterified fatty acids and β-hydroxybutyrate and disease incidence, pregnancy rate, and milk production at the herd level. J. Dairy Sci. 2010, 93, 3595–3601. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Contreras, G.A.; Aitken, S.L. Metabolic factors affecting the inflammatory response of periparturient dairy cows. Anim. Health. Res. Rev. 2009, 10, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P. Physiologic factors to consider in order to prevent periparturient disease in the dairy cow, with special emphasis on milk fever. In Proceedings of the 3rd Western Dairy Management Conference, Las Vegas, NV, USA, 8–10 April 1999. [Google Scholar]
- Hayirli, A.; Grummer, R.R.; Nordheim, E.V.; Crump, P.M. Models for Predicting Dry Matter Intake of Holsteins During the Prefresh Transition Period. J. Dairy Sci. 2003, 86, 1771–1779. [Google Scholar] [CrossRef] [Green Version]
- Duehlmeier, R.; Fluegge, I.; Schwert, B.; Ganter, M. Insulin Sensitivity during Late Gestation in Ewes Affected by Pregnancy Toxemia and in Ewes with High and Low Susceptibility to this Disorder. J. Vet. Intern. Med. 2013, 27, 359–366. [Google Scholar] [CrossRef]
- Karagiannis, I.; Brozos, C.; Kiossis, E.; Boscos, C.; Fthenakis, G.; Panousis, N. Associations of periparturient β-hydroxybutyric acid and non-esterified fatty acids blood serum concentrations with milk yield, milk composition and milk somatic cells count of intensively managed Chios dairy ewes. J. Hell. Vet. Med. Soc. 2019, 69, 1195. [Google Scholar] [CrossRef] [Green Version]
- Karagiannis, I.; Panousis, N.; Kiossis, E.; Tsakmakidis, I.; Lafi, S.; Arsenos, G.; Boscos, C.; Brozos, C. Associations of pre-lambing body condition score and serum β-hydroxybutyric acid and non-esterified fatty acids concentrations with periparturient health of Chios dairy ewes. Small Rumin. Res. 2014, 120, 164–173. [Google Scholar] [CrossRef]
- Brozos, C.; Mavrogianni, V.S.; Fthenakis, G.C. Treatment and Control of Peri-Parturient Metabolic Diseases: Pregnancy Toxemia, Hypocalcemia, Hypomagnesemia. Vet. Clin. North Am. Food Anim. Pract. 2011, 27, 105–113. [Google Scholar] [CrossRef]
- Panousis, N.; Brozos, C.; Karagiannis, I.; Giadinis, N.D.; Lafi, S.; Kritsepi-Konstantinou, M. Evaluation of Precision Xceed® meter for on-site monitoring of blood β-hydroxybutyric acid and glucose concentrations in dairy sheep. Res. Vet. Sci. 2012, 93, 435–439. [Google Scholar] [CrossRef]
- Daniel, R.C.W. Motility of the Rumen and Abomasum During Hypocalcaemia. Can. J. Comp. Med. 1983, 47, 276–280. [Google Scholar]
- Goff, J.P. The monitoring, prevention, and treatment of milk fever and subclinical hypocalcemia in dairy cows. Vet. J. 2008, 176, 50–57. [Google Scholar] [CrossRef]
- Martinez, N.; Risco, C.A.; Lima, F.S.; Bisinotto, R.S.; Greco, L.F.; Ribeiro, E.S.; Maunsell, F.; Galvão, K.; Santos, J.E.P. Evaluation of peripartal calcium status, energetic profile, and neutrophil function in dairy cows at low or high risk of developing uterine disease. J. Dairy Sci. 2012, 95, 7158–7172. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, A.N.; Sleiman, F.T. Beet Pulp as a Grain Replacement for Dairy Cows and Sheep. J. Dairy Sci. 1971, 54, 89–94. [Google Scholar] [CrossRef]
- Mustafa, M.M.M. Effect of Dried Olive Oil By-Product Supplementation to Ration on The Performance of Local Ewes and Their Lambs. Isot. Radiat. Res. 2011, 43, 77–93. [Google Scholar]
- Bendary, M.M.; Mahmoud, A.M.; Koriet, I.S.; Abd-El-Raouf, E.M.; Awadalla, S.A. Nutritional studies on using sugar beet tops in animal feeding. 4- Fattening Friesian calves on different forms on sugar beet tops. J. Agric. Sci. Mansoura Univ. 1992, 17, 2871–2880. [Google Scholar]
- Dickie, C.W.; Hamann, M.H.; Carroll, W.D.; Chow, F.H. Oxalate (Rumex venosus) poisoning in cattle. J. Am. Vet. Med. Assoc. 1978, 173, 73–74. [Google Scholar]
- El-Khodery, S.; El-Boshy, M.; Gaafar, K.; Elmashad, A. Hypocalcemia in Ossimi Sheep Associated with Feeding on Beet Tops (Beta vulgaris). Turk. J. Vet. Anim. Sci. 2008, 32, 199–205. [Google Scholar]
- Goff, J.P.; Horst, R. Physiological Changes at Parturition and Their Relationship to Metabolic Disorders. J. Dairy Sci. 1997, 80, 1260–1268. [Google Scholar] [CrossRef]
- Grunberg, W.; Constable, P.; Schroder, U.; Staufenbiel, R.; Morin, D.; Rohn, M. Phosphorus Homeostasis in Dairy Cows with Abomasal Displacement or Abomasal Volvulus. J. Vet. Intern. Med. 2005, 19, 894–898. [Google Scholar] [CrossRef]
- Horst, R.L.; Goff, J.P.; Reinhardt, T.A. Calcium and Vitamin D Metabolism in the Dairy Cow. J. Dairy Sci. 1994, 77, 1936–1951. [Google Scholar] [CrossRef] [PubMed]
- Masters, D.G.; Hancock, S.; Refshauge, G.; Robertson, S.M.; McGrath, S.; Bhanugopan, M.; Friend, M.A.; Thomson, A.N. Mineral supplements improve the calcium status of pregnant ewes grazing vegetative cereals. Anim. Prod. Sci. 2019, 59, 1299–1309. [Google Scholar] [CrossRef]
- Kohestani, M.G.; Yansari, A.T.; Rezaei, M. Effects of partial replacement of barley with sugar beet pulp on pre-and post-partum performance of Zel ewes. S. Afr. J. Anim. Sci. 2011, 41, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Oetzel, G.R. Management of dry cows for the prevention of milk fever and other mineral disorders. Vet. Clin. N. Am. Food A 2000, 16, 369–386. [Google Scholar] [CrossRef]
- Constable, P.D.; Megahed, A.A.; Hiew, M.W.H. Measurement of urine pH and net acid excretion and their association with urine calcium excretion in periparturient dairy cows. J. Dairy Sci. 2019, 102, 11370–11383. [Google Scholar] [CrossRef]
- Borsberry, S.; Hobson, H. Periparturient diseases and their effect on reproductive performance of five dairy herds. Vet. Rec. 1989, 124, 217–219. [Google Scholar] [CrossRef]
- Curtis, C.R.; Erb, H.N.; Sniffen, C.J.; Smith, R.D.; Powers, P.A.; Smith, M.C.; White, M.E.; Hillman, R.B.; Pearson, E.J. Association of parturient hypocalcemia with eight periparturient disorders in Holstein cows. J. Am. Vet. Med. Assoc. 1983, 183, 559–561. [Google Scholar]
- Goff, J.P. Macromineral disorders of the transition cow. Vet. Clin. Food Anim. Pract. 2004, 20, 471–494. [Google Scholar] [CrossRef]
- Martens, H.; Leonhard- Marek, S.; Rontgen, M.; Stumpff, F. Magnesium homeostasis in cattle: Absorption and excertion. Nutr. Res. Rev. 2018, 31, 114–130. [Google Scholar] [CrossRef]
- Charbonneau, E.; Pellerin, D.; Oetzel, G.R. Impact of Lowering Dietary Cation-Anion Difference in Nonlactating Dairy Cows: A Meta-Analysis. J. Dairy Sci. 2006, 89, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Lean, I.J.; Santos, J.E.P.; Block, E.; Golder, H.M. Effects of prepartum dietary cation-anion difference intake on production and health of dairy cows: A meta-analysis. J. Dairy Sci. 2019, 102, 2103–2133. [Google Scholar] [CrossRef] [Green Version]
- Goff, J.; Horst, R. Role of acid-base physiology on the pathogenesis of parturient hypocalcaemia (milk fever)—The DCAD theory in principal and practice. Acta Vet. Scand. Suppl. 2003, 97, 51–56. [Google Scholar]
- Goff, J.P.; Liesegang, A.; Horst, R.L. Diet-induced pseudohypoparathyroidism: Ahypocalcemia and milk fever risk factor. J. Dairy Sci. 2014, 97, 1520–1528. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, I. Recent advances in physiological calcium homeostasis. Clin. Chem. Lab. Med. 2006, 44, 237–273. [Google Scholar] [CrossRef]
- Constable, P.D.; Gelfert, C.C.; Fürll, M.; Staufenbiel, R.; Stämpfli, H.R. Application of strong ion difference theory to urine and the relationship between urine pH and net acid excretion in cattle. Am. J. Vet. Res. 2009, 70, 915–925. [Google Scholar] [CrossRef]
- Oetzel, G.R.; Olson, J.D.; Curtis, C.R.; Fettman, M.J. Ammonium Chloride and Ammonium Sulfate for Prevention of Parturient Paresis in Dairy Cows. J. Dairy Sci. 1988, 71, 3302–3309. [Google Scholar] [CrossRef]
- Roubies, N.; Panousis, N.; Fytianou, A.; Katsoulos, P.D.; Giadinis, N.; Karatzias, H. Effects of Age and Reproductive Stage on Certain Serum Biochemical Parameters of Chios Sheep Under Greek Rearing Conditions. J. Vet. Med. Ser. A 2006, 53, 277–281. [Google Scholar] [CrossRef]
- DeGaris, P.J.; Lean, I.J. Milk fever in dairy cows: A review of pathophysiology and control principles. Vet. J. 2008, 176, 58–69. [Google Scholar] [CrossRef]
- Grummer, R.R. Impact of changes in organic nutrient metabolism on feeding the transition dairy cow. J. Anim. Sci. 1995, 73, 2820. [Google Scholar] [CrossRef]
- Chapinal, N.; LeBlanc, S.J.; Carson, M.E.; Leslie, K.E.; Godden, S.; Capel, M.; Santos, J.E.P.; Overton, M.W.; Duffield, T.F. Herd-level association of serum metabolites in the transition period with disease, milk production, and early lactation reproductive performance. J. Dairy Sci. 2012, 95, 5676–5682. [Google Scholar] [CrossRef] [Green Version]
- Liesegang, A.; Risteli, J. Influence of different calcium concentrations in the diet on bone metabolism in growing dairy goats and sheep. J. Anim. Physiol. Anim. Nutr. 2005, 89, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Kimberling, C.V. Diseases of ewes. In Jensen and Swift’s Diseases of Sheep, 3rd ed.; Jensen, R., Swift, B.L., Kimberling, C.V., Eds.; Lea & Febiger: Philadelphia, PA, USA, 1988; pp. 26–29. [Google Scholar]
- Radostits, O.M.; Gay, C.C.; Hinchcliff, K.W.; Constable, P.D. Parturient paresis. In Veterinary Medicine: A Textbook of the Diseases of Cattle, Sheep, Pigs, Goats, And Horses, 10th ed.; Radostits, O.M., Done, S.H., Eds.; Elsevier Saunders: New York, NY, USA, 2007; pp. 1626–1644. [Google Scholar]
- Woldemeskel, M.; Eneyew, M.; Kassa, T. Study on ovine hypocalcaemia in ewes in central Ethiopia. Rev. Med. Vet. 2012, 151, 345–350. [Google Scholar]
- Schonewille, J.T.; Everts, H.; Jittakhof, S.; Beynen, A. Quantitive prediction of magnesium absorption in dairy cows. J. Diary Sci. 2008, 91, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontenot, J.P.; Allen, V.G.; Bunce, G.E.; Goff, J.P. Factors Influencing Magnesium Absorption and Metabolism in Ruminants. J. Anim. Sci. 1989, 67, 3445. [Google Scholar] [CrossRef] [PubMed]
- Sykes, A.R. Deficiency of mineral macro-elements. In Diseases of Sheep, 4th ed.; Aitken, I., Ed.; Blackwell Publishing: Oxford, UK, 2007; p. ch 53. [Google Scholar] [CrossRef]
- Rayssiguier, Y. Hypomagnesemia resulting from adrenaline infusion in ewes: Its relation to lipolysis. Horm. Metab. Res. 1977, 9, 309–314. [Google Scholar] [CrossRef]
- Persson, J.; Luthman, J. The effects of insulin, glucose and catecholamines on some blood minerals in sheep. Acta Vet. Scand. 1974, 15, 519–532. [Google Scholar] [CrossRef]
- Azab, M.E.; Abdel-Maksoud, H.A. Changes in some hematological and biochemical parameters during prepartum and postpartum periods in female Baladi goats. Small Rumin. Res. 1999, 34, 77–85. [Google Scholar] [CrossRef]
- Goff, J.P. Pathophysiology of calcium and phosphorus disorders. Vet. Clin. N. Am. Food Anim. Pract. 2000, 16, 319–338. [Google Scholar] [CrossRef]
- Garces, M.A.; Evans, J.L. Calcium and Magnesium Absorption in Growing Cattle as Influenced by Age of Animal and Source of Dietary Nitrogen. J. Anim. Sci. 1971, 32, 789–793. [Google Scholar] [CrossRef]
- Hansard, S.L.; Comar, C.L.; Davis, G.K. Effects of Age Upon the Physiological Behavior of Calcium in Cattle. Am. J. Physiol. Leg. Content 1954, 177, 383–389. [Google Scholar] [CrossRef]
- Schonewille, J.T.; Van Klooster, A.T.; Wouterse, H.; Beynen, A.C. Hypocalcemia induced by intravenous administration of disodium ethylenediaminotetraacetate and its effects on excretion of calcium in urine of cows fed a high chloride diet. J. Dairy Sci. 1999, 82, 1317–1324. [Google Scholar] [CrossRef]
- Durak, M.H.; Altiner, A. Effect of energy deficiency during late pregnancy in Chios Ewes on free fatty acids, β hydroxybutyrate and urea metabolites. Turk. J. Vet. Anim. Sci. 2006, 30, 497–502. [Google Scholar]
- Harmeyer, J.; Schlumbohm, C. Pregnancy impairs ketone body disposal in late gestating ewes: Implications for onset of pregnancy toxemia. Res. Vet. Sci. 2006, 81, 254–264. [Google Scholar] [CrossRef]
- Duehlmeier, R.; Fluegge, I.; Schwert, B.; Prvizi, N.; Ganter, M. Metabolic adaptations to pregnancy and lactation in German Blackheaded Mutton and Finn sheep ewes affected by pregnancy toxemia and in ewes with high and low susceptibilities to this disorder. Small Rumin. Res. 2011, 96, 178–184. [Google Scholar] [CrossRef]
- Raoofi, A.; Jafarian, M.; Safi, S.; Vatankhah, M. Fluctuations in energy-related metabolites during the peri-parturition period in Lori-Bakhtiari ewes. Small Rumin. Res. 2013, 109, 64–68. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
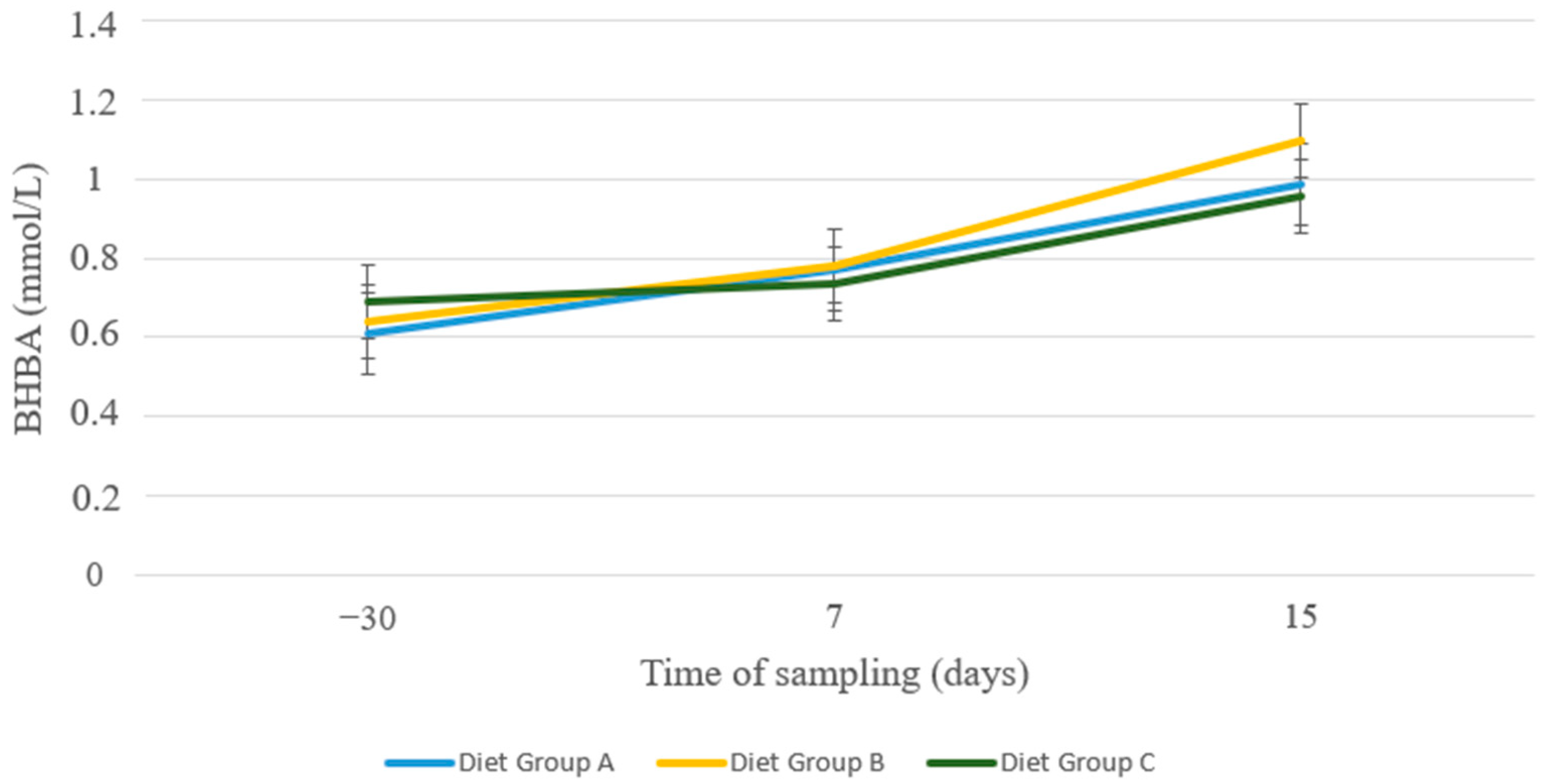{kind=link}
| Concentrate | Group A | Group B | Group C |
|---|---|---|---|
| Corn | 500 | 469 | 444 |
| Barley | 72 | - | - |
| Soya | 100 | 100 | 100 |
| Wheat bran | 290 | - | - |
| Sugar beet pulp | - | 400 | 400 |
| NH4Cl | - | - | 25 |
| CMV * | 2 | 2 | 2 |
| CaCo3 | 18.5 | - | - |
| CaHPO4 | 1.5 | 14 | 14 |
| NaCl | 10.5 | 7 | 7 |
| MgO | 5.5 | 8 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peleki, C.; Kiosis, E.; Polizopoulou, Z.S.; Tsousis, G.; Fthenakis, G.C.; Giadinis, N.D.; Brozos, C. Effect of Sugar Beet Pulp and Anionic Salts on Metabolic Status and Mineral Homeostasis during the Peri-Parturient Period of Dairy Sheep. Animals 2023, 13, 213. https://doi.org/10.3390/ani13020213
Peleki C, Kiosis E, Polizopoulou ZS, Tsousis G, Fthenakis GC, Giadinis ND, Brozos C. Effect of Sugar Beet Pulp and Anionic Salts on Metabolic Status and Mineral Homeostasis during the Peri-Parturient Period of Dairy Sheep. Animals. 2023; 13(2):213. https://doi.org/10.3390/ani13020213
Chicago/Turabian StylePeleki, Christina, Evangelos Kiosis, Zoe S. Polizopoulou, Georgios Tsousis, George C. Fthenakis, Nektarios D. Giadinis, and Christos Brozos. 2023. "Effect of Sugar Beet Pulp and Anionic Salts on Metabolic Status and Mineral Homeostasis during the Peri-Parturient Period of Dairy Sheep" Animals 13, no. 2: 213. https://doi.org/10.3390/ani13020213