
Exploring the Domestication Syndrome Hypothesis in Dogs: Pigmentation Does Not Predict Cortisol Levels

Abstract
:Simple Summary
Abstract
1. Introduction
1.1. The Domestication Syndrome Hypothesis
1.2. Dog Pigmentation Overview
1.3. Dogs and Stress
2. Materials and Methods
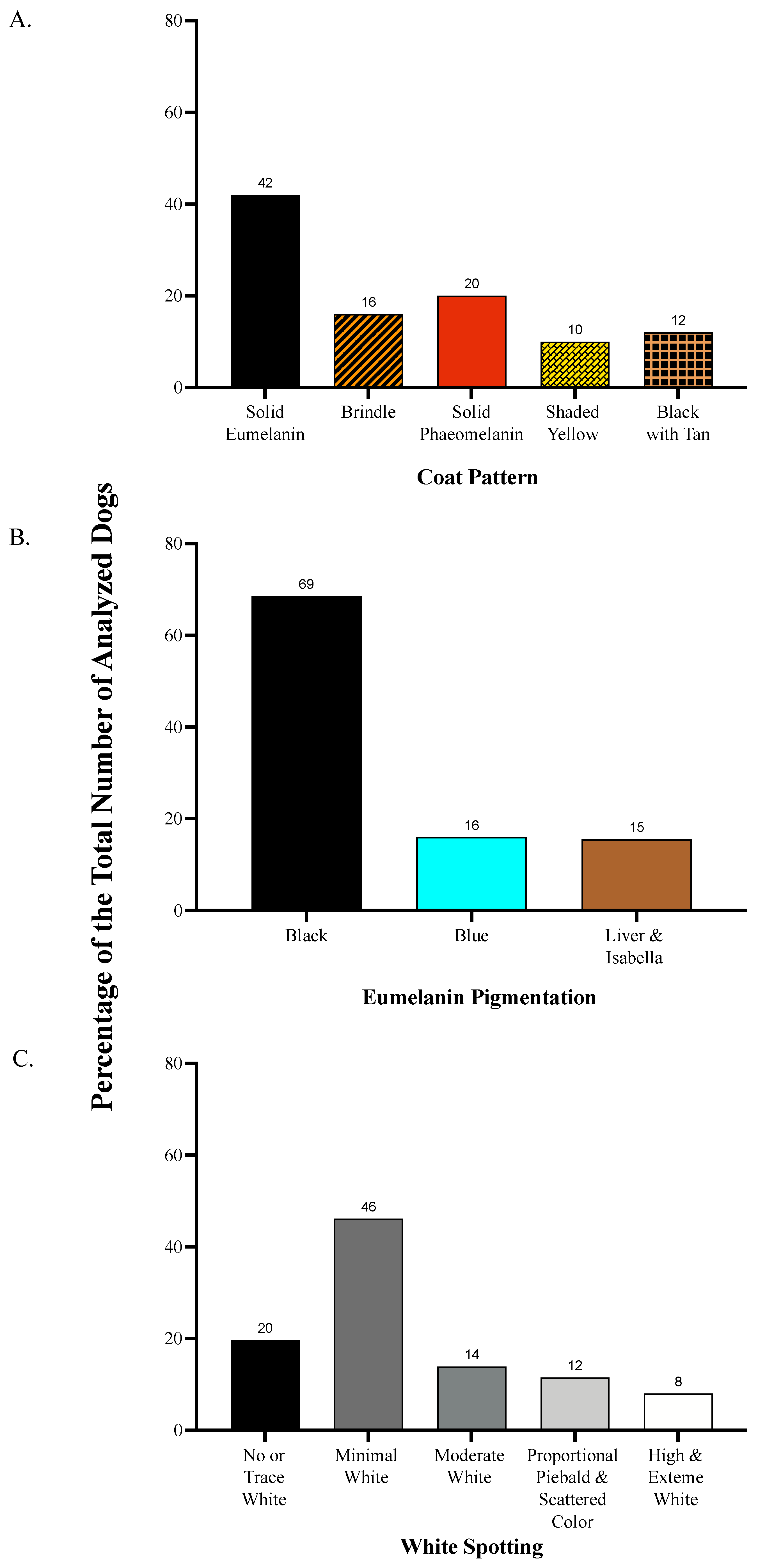
2.1. Visual Phenotyping
2.1.1. Coat Pattern
2.1.2. Eumelanin Color
2.1.3. White Spotting
2.2. Cortisol Collection
2.2.1. Brief Outing Intervention
2.2.2. Weeklong Fostering Intervention
2.3. Analysis
3. Results
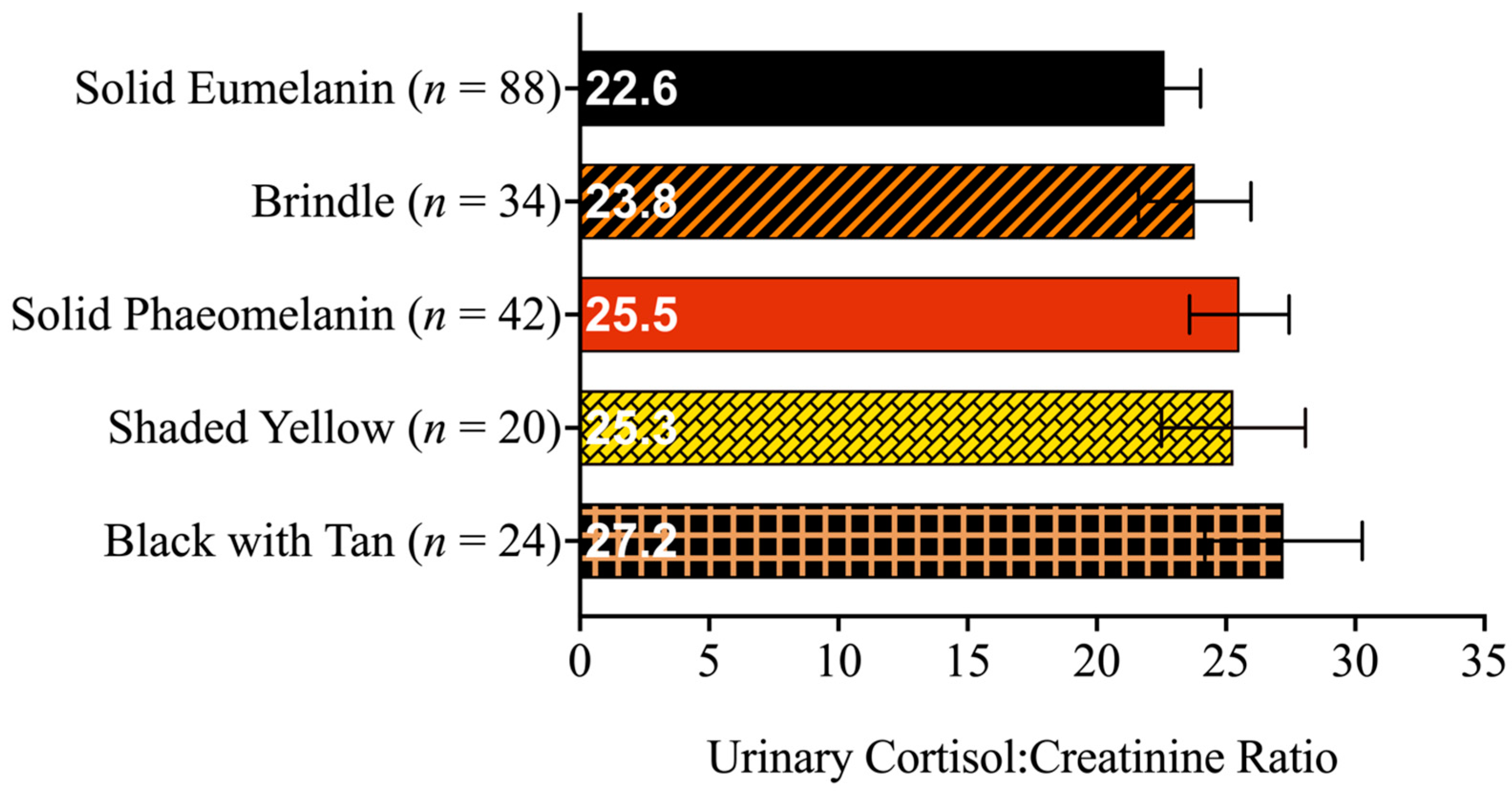
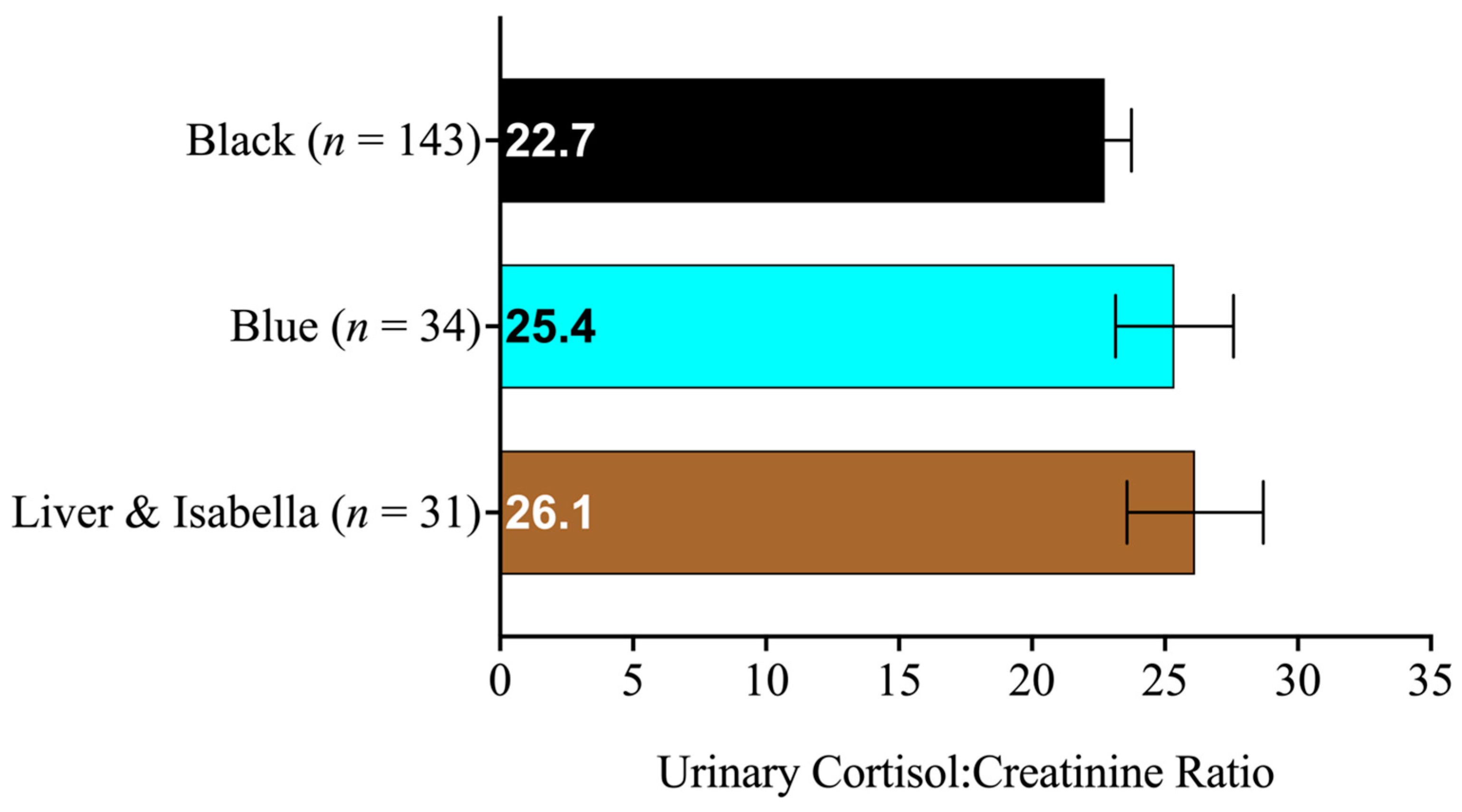
3.1. Descriptive Statistics
3.2. Linear Mixed Models
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darwin, C. The Variation of Animals and Plants under Domestication (Vol. 2). J. Murray. 1868. Available online: https://www.gutenberg.org/files/3332/3332-h/3332-h.htm (accessed on 10 February 2021).
- Zeder, M.A. The Domestication of Animals. J. Anthropol. Res. 2012, 68, 161–190. [Google Scholar] [CrossRef]
- Minton, J.E. Function of the hypothalamic-pituitary-adrenal axis and the sympathetic nervous system in models of acute stress in domestic farm animals2. J. Anim. Sci. 1994, 72, 1891–1898. [Google Scholar] [CrossRef] [PubMed]
- Newberry, R. Behavioral, emotional, & cognitive effects of domestication. In APA Handbook of Comparative Psychology: Vol. 1. Basic Concepts, Methods, Neural Substrate, and Behavior; American Psychological Association: Washington, DC, USA, 2017; Chapter 16; pp. 315–329. [Google Scholar] [CrossRef]
- Lord, K.A.; Larson, G.; Coppinger, R.P.; Karlsson, E.K. The History of Farm Foxes Undermines the Animal Domestication Syndrome. Trends Ecol. Evol. 2020, 35, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Villagra, M.R.; Geiger, M.; Schneider, R.A. The taming of the neural crest: A developmental perspective on the origins of morphological covariation in domesticated mammals. R. Soc. Open Sci. 2016, 3, 160107. [Google Scholar] [CrossRef]
- Trut, L.; Oskina, I.; Kharlamova, A. Animal evolution during domestication: The domesticated fox as a model. BioEssays News Rev. Mol. Cell. Dev. Biol. 2009, 31, 349–360. [Google Scholar] [CrossRef]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and Genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef]
- Brancalion, L.; Haase, B.; Wade, C.M. Canine coat pigmentation genetics: A review. Anim. Genet. 2022, 53, 3–34. [Google Scholar] [CrossRef]
- Linderholm, A.; Larson, G. The Role of Humans in Facilitating and Sustaining Coat Colour Variation in Domestic Animals. Semin. Cell Dev. Biol. 2013, 24, 587–593. [Google Scholar] [CrossRef]
- Rigg, R. Livestock Guarding Dogs: Their Current Use World Wide; IUCN/SSC Canid Specialist Group Occasional Paper; IUCN: Gland, Switzerland, 2001; Available online: http://www.canids.org/occasionalpapers/ (accessed on 21 July 2023).
- Saif, R.; Iftekhar, A.; Asif, F.; Alghanem, M.S. Dog Coat Colour Genetics: A Review. Adv. Life Sci. 2020, 7, 215–224. [Google Scholar]
- Grandin, T. The Way I See It: The Dangers of Overselection; Western Horseman: Fort Worth, TX, USA, 1998; pp. 120–124. Available online: https://mountainscholar.org/bitstream/handle/10217/4424/H105.pdf;sequence=1 (accessed on 10 February 2021).
- Grandin, T.; Deesing, M. Genetics and Animal Welfare. Available online: https://www.grandin.com/welfare/genetics.animal.welfare.html (accessed on 6 April 2021).
- Daniels, T.J.; Bekoff, M. Population and Social Biology of Free-Ranging Dogs, Canis familiaris. J. Mammal. 1989, 70, 754–762. [Google Scholar] [CrossRef]
- Schmutz, S.M.; Berryere, T.G. Genes affecting coat colour and pattern in domestic dogs: A review. Anim. Genet. 2007, 38, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Berryere, T.G.; Dreger, D.L. MITF and White Spotting in Dogs: A Population Study. J. Hered. 2009, 100, S66–S74. [Google Scholar] [CrossRef]
- Baranowska Körberg, I.; Sundström, E.; Meadows, J.R.S.; Rosengren Pielberg, G.; Gustafson, U.; Hedhammar, Å.; Karlsson, E.K.; Seddon, J.; Söderberg, A.; Vilà, C.; et al. A Simple Repeat Polymorphism in the MITF-M Promoter Is a Key Regulator of White Spotting in Dogs. PLoS ONE 2014, 9, e104363. [Google Scholar] [CrossRef] [PubMed]
- Beerda, B.; Schilder, M.B.H.; Van Hooff, J.A.R.A.M.; De Vries, H.W.; Mol, J.A. Chronic Stress in Dogs Subjected to Social and Spatial Restriction. I. Behavioral Responses. Physiol. Behav. 1999, 66, 233–242. [Google Scholar] [CrossRef]
- Scheifele, P.; Martin, D.; Clark, J.G.; Kemper, D.; Wells, J. Effect of kennel noise on hearing in dogs. Am. J. Vet. Res. 2012, 73, 482–489. [Google Scholar] [CrossRef]
- Hellhammer, D.H.; Wüst, S.; Kudielka, B.M. Salivary cortisol as a biomarker in stress research. Psychoneuroendocrinology 2009, 34, 163–171. [Google Scholar] [CrossRef]
- Beerda, B.; Schilder, M.B.H.; Janssen, N.S.C.R.M.; Mol, J.A. The Use of Saliva Cortisol, Urinary Cortisol, and Catecholamine Measurements for a Noninvasive Assessment of Stress Responses in Dogs. Horm. Behav. 1996, 30, 272–279. [Google Scholar] [CrossRef]
- Schatz, S.; Palme, R. Measurement of Faecal Cortisol Metabolites in Cats and Dogs: A Non-invasive Method for Evaluating Adrenocortical Function. Vet. Res. Commun. 2001, 25, 271–287. [Google Scholar] [CrossRef]
- Sandri, M.; Colussi, A.; Perrotta, M.G.; Stefanon, B. Salivary cortisol concentration in healthy dogs is affected by size, sex, and housing context. J. Vet. Behav. 2015, 10, 302–306. [Google Scholar] [CrossRef]
- Dudley, E.S.; Schiml, P.A.; Hennessy, M.B. Effects of repeated petting sessions on leukocyte counts, intestinal parasite prevalence, and plasma cortisol concentration of dogs housed in a county animal shelter. J. Am. Vet. Med. Assoc. 2015, 247, 1289–1298. [Google Scholar] [CrossRef]
- Gunter, L.M.; Feuerbacher, E.N.; Gilchrist, R.J.; Wynne, C.D.L. Evaluating the effects of a temporary fostering program on shelter dog welfare. PeerJ 2019, 7, e6620. [Google Scholar] [CrossRef] [PubMed]
- Gunter, L.M.; Gilchrist, R.J.; Blade, E.M.; Barber, R.T.; Feuerbacher, E.N.; Platzer, J.M.; Wynne, C.D.L. Investigating the Impact of Brief Outings on the Welfare of Dogs Living in US Shelters. Animals 2021, 11, 548. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, M.B.; Voith, V.L.; Mazzei, S.J.; Buttram, J.; Miller, D.D.; Linden, F. Behavior and cortisol levels of dogs in a public animal shelter, and an exploration of the ability of these measures to predict problem behavior after adoption. Appl. Anim. Behav. Sci. 2001, 73, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Houpt, K.A.; Willis, M.B. Genetics of behavior. In The Genetics of the Dog; Ruvinsky, A., Sampson, J., Eds.; CABI Pub: Wallingford, UK; New York, NY, USA, 2001; ISBN 978-0-85199-520-5. [Google Scholar]
- Pérez-Guisado, J.; Lopez-Rodríguez, R.; Muñoz-Serrano, A. Heritability of dominant–aggressive behaviour in English Cocker Spaniels. Appl. Anim. Behav. Sci. 2006, 100, 219–227. [Google Scholar] [CrossRef]
- Podberscek, A.L.; Serpell, J.A. The English Cocker Spaniel: Preliminary findings on aggressive behaviour. Appl. Anim. Behav. Sci. 1996, 47, 75–89. [Google Scholar] [CrossRef]
- van Rooy, D.; Wade, C.M. Association between coat colour and the behaviour of Australian Labrador retrievers. Canine Genet. Epidemiol. 2019, 6, 10. [Google Scholar] [CrossRef]
- Kim, Y.K.; Lee, S.S.; Oh, S.I.; Kim, J.S.; Suh, E.H.; Houpt, K.A.; Lee, H.C.; Lee, H.J.; Yeon, S.C. Behavioural reactivity of the Korean native Jindo dog varies with coat colour. Behav. Process. 2010, 84, 568–572. [Google Scholar] [CrossRef]
- Lofgren, S.E.; Wiener, P.; Blott, S.C.; Sanchez-Molano, E.; Woolliams, J.A.; Clements, D.N.; Haskell, M.J. Management and personality in Labrador Retriever dogs. Appl. Anim. Behav. Sci. 2014, 156, 44–53. [Google Scholar] [CrossRef]
- Roulin, A.; Ducrest, A.-L. Association between melanism, physiology and behaviour: A role for the melanocortin system. Eur. J. Pharmacol. 2011, 660, 226–233. [Google Scholar] [CrossRef]
- Reissmann, M.; Ludwig, A. Pleiotropic effects of coat colour-associated mutations in humans, mice and other mammals. Semin. Cell Dev. Biol. 2013, 24, 576–586. [Google Scholar] [CrossRef]
- Mills, D.; Karagiannis, C.; Zulch, H. Stress—its effects on health and behavior. Vet. Clin. N. Am. Small Anim. Pract. 2014, 44, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Gunter, L. The Influences of Weeklong Fostering and Co-Housing on Shelter Dog Welfare; [Manuscript in preparation]; Virginia Tech: Blacksburg, VA, USA, 2023. [Google Scholar]
- Bannasch, D.L.; Kaelin, C.B.; Letko, A.; Loechel, R.; Hug, P.; Jagannathan, V.; Henkel, J.; Roosje, P.; Hytönen, M.K.; Lohi, H.; et al. Dog colour patterns explained by modular promoters of ancient canid origin. Nat. Ecol. Evol. 2021, 5, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Morrill, K.; Hekman, J.; Li, X.; McClure, J.; Logan, B.; Goodman, L.; Gao, M.; Dong, Y.; Alonso, M.; Carmichael, E.; et al. Ancestry-inclusive dog genomics challenges popular breed stereotypes. Science 2022, 376, eabk0639. [Google Scholar] [CrossRef]
- Hansen Wheat, C.; van der Bijl, W.; Wheat, C.W. Morphology does not covary with predicted behavioral correlations of the domestication syndrome in dogs. Evol. Lett. 2020, 4, 189–199. [Google Scholar] [CrossRef]
- Hansen Wheat, C.; Fitzpatrick, J.L.; Rogell, B.; Temrin, H. Behavioural correlations of the domestication syndrome are decoupled in modern dog breeds. Nat. Commun. 2019, 10, 2422. [Google Scholar] [CrossRef]
- Padmabandu, M. Top 10 Most Common Breeds in Embark-Tested Dogs. Embark. 2022. Available online: https://embarkvet.com/resources/most-common-dog-breed-ancestry/ (accessed on 5 May 2023).
- Trut, L.N.; Plyusnina, I.Z.; Oskina, I.N. An experiment on fox domestication and debatable issues of evolution of the dog. Russ. J. Genet. 2004, 40, 644–655. [Google Scholar] [CrossRef]
- Protopopova, A.; Brandifino, M.; Wynne, C.D.L. Preference assessments and structured potential adopter-dog interactions increase adoptions. Appl. Anim. Behav. Sci. 2016, 176, 87–95. [Google Scholar] [CrossRef]
- Iyasere, O.S.; Durosaro, S.O.; Oyeniran, V.J.; Daramola, J.O. Is an increase of glucocorticoid concentrations related to the degree of arousal or valence experienced by an animal to a stimulus? Domest. Anim. Endocrinol. 2022, 81, 106752. [Google Scholar] [CrossRef]
- Novais, A.; Monteiro, S.; Roque, S.; Correia-Neves, M.; Sousa, N. How age, sex and genotype shape the stress response. Neurobiology 2017, Stress 6, 44–56. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pigmentation Variable | Brief Outing Intervention | Weeklong Fostering Intervention | ||||
|---|---|---|---|---|---|---|
| Before | During | After | Before | During | After | |
| M (SE) | M (SE) | M (SE) | M (SE) | M (SE) | M (SE) | |
| Coat Pattern | ||||||
| Solid Eumelanin | 28.3 (1.7) | 30.5 (1.8) | 27.9 (1.7) | 19.0 (2.6) | 11.9 (2.7) | 18.1 (2.6) |
| Brindle | 29.3 (2.8) | 28.9 (3.0) | 31.4 (2.8) | 19.6 (3.9) | 16.3 (4.1) | 17.2 (4.0) |
| Solid Pheomelanin | 32.2 (2.5) | 36.1 (2.7) | 36.2 (2.5) | 20.3 (3.4) | 11.7 (3.6) | 16.7 (3.4) |
| Shaded Yellow | 31.0 (4.8) | 39.4 (5.0) | 35.3 (4.8) | 16.8 (3.8) | 13.2 (4.0) | 16.0 (3.8) |
| Black with Tan | 38.4 (5.9) | 46.7 (6.2) | 34.1 (5.9) | 18.3 (3.1) | 9.7 (3.3) | 16.1 (3.1) |
| Eumelanin Pigmentation | ||||||
| Black | 28.5 (1.5) | 33.1 (1.6) | 29.6 (1.5) | 17.8 (1.6) | 11.3 (1.7) | 16.2 (1.6) |
| Blue | 30.1 (2.7) | 29.4 (2.9) | 31.5 (2.8) | 22.9 (4.0) | 16.0 (4.3) | 22.2 (4.1) |
| Liver or Isabella | 34.6 (2.7) | 34.6 (2.8) | 35.7 (2.7) | 22.2 (5.0) | 13.5 (5.2) | 16.3 (5.0) |
| White Spotting | ||||||
| No or Trace White | 35.3 (2.9) | 39.2 (3.0) | 35.0 (2.9) | 14.0 (2.9) | 10.0 (3.1) | 13.0 (3.0) |
| Minimal | 29.5 (1.7) | 31.2 (1.8) | 32.2 (1.7) | 19.7 (2.1) | 13.2 (2.2) | 19.0 (2.1) |
| Moderate | 26.7 (3.2) | 31.2 (3.3) | 26.2 (3.1) | 19.4 (3.9) | 12.6 (4.2) | 15.7 (4.0) |
| Proportional Piebald or Scattered Color | 27.5 (3.2) | 30.0 (3.4) | 25.9 (3.2) | 27.6 (4.6) | 12.2 (4.8) | 20.2 (4.6) |
| High or Extreme White | 25.9 (3.9) | 27.7 (4.1) | 28.5 (3.9) | 17.6 (5.3) | 12.2 (5.6) | 16.5 (5.3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Platzer, J.M.; Gunter, L.M.; Feuerbacher, E.N. Exploring the Domestication Syndrome Hypothesis in Dogs: Pigmentation Does Not Predict Cortisol Levels. Animals 2023, 13, 3095. https://doi.org/10.3390/ani13193095
Platzer JM, Gunter LM, Feuerbacher EN. Exploring the Domestication Syndrome Hypothesis in Dogs: Pigmentation Does Not Predict Cortisol Levels. Animals. 2023; 13(19):3095. https://doi.org/10.3390/ani13193095
Chicago/Turabian StylePlatzer, JoAnna M., Lisa M. Gunter, and Erica N. Feuerbacher. 2023. "Exploring the Domestication Syndrome Hypothesis in Dogs: Pigmentation Does Not Predict Cortisol Levels" Animals 13, no. 19: 3095. https://doi.org/10.3390/ani13193095