
Effect of Yeast Culture on Reproductive Performance, Gut Microbiota, and Milk Composition in Primiparous Sows
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Design
2.2. Sample Collection and Measurement
2.3. Chemical Analysis of Feed and Feces
2.4. SCFA Analysis
2.5. Microbial Analysis
2.6. Metabolite Extraction and Analysis
2.7. Statistical Analysis
3. Results
3.1. Production Performance of Primiparous Sows
3.2. The ATTD of Primiparous Sows during Lactation
3.3. The SCFA Concentration in Fecal Samples of Primiparous Sows
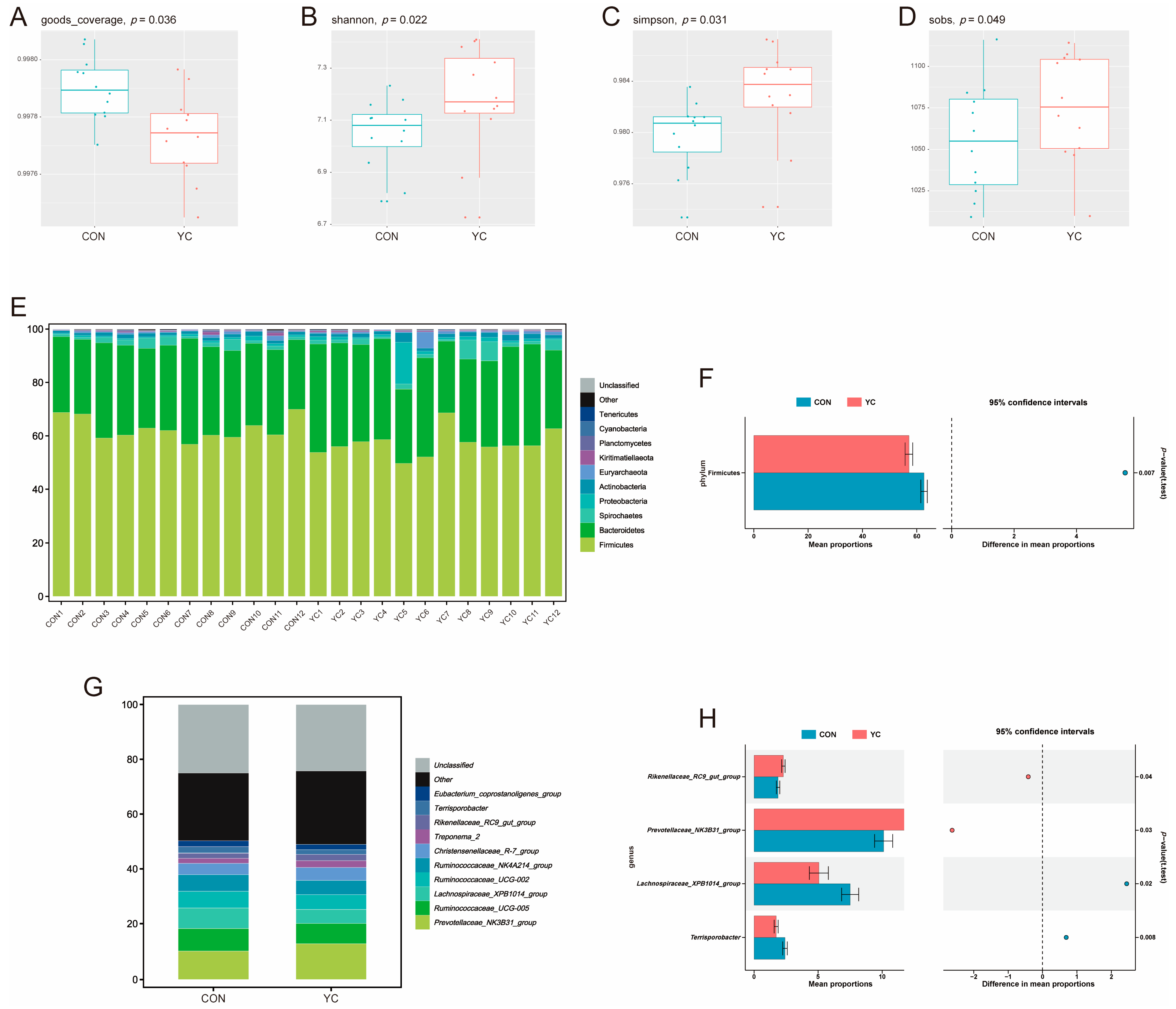
3.4. Fecal Microbiota Analysis of Primiparous Sows
3.5. Colostrum and Milk Composition
3.6. Metabolomic Analysis of Colostrum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, B.; Zhou, Y.F.; Xia, X.; Wang, C.; Wei, H.K.; Peng, J. Effects of Dietary Lysine Levels on Production Performance and Milk Composition of High-Producing Sows during Lactation. Animals 2020, 10, 12. [Google Scholar] [CrossRef]
- Costermans, N.G.J.; Teerds, K.J.; Kemp, B.; Keijer, J.; Soede, N.M. Physiological and metabolic aspects of follicular developmental competence as affected by lactational body condition loss. Mol. Reprod. Dev. 2022, 12, 491–502. [Google Scholar] [CrossRef]
- Kim, S.W.; Brandherm, M.; Freeland, M.; Newton, B.; Cook, D.; Yoon, I. Effects of yeast culture supplementation to gestation and lactation diets on growth of nursing piglets. Asian Australas. J. Anim. Sci. 2008, 21, 1011–1014. [Google Scholar] [CrossRef]
- Clowes, E.J.; Aherne, F.X.; Foxcroft, G.R.; Baracos, V.E. Selective protein loss in lactating sows is associated with reduced litter growth and ovarian function. J. Anim. Sci. 2003, 81, 753–764. [Google Scholar] [CrossRef]
- Schenkel, A.C.; Bernardi, M.L.; Bortolozzo, F.P.; Wentz, I. Body reserve mobilization during lactation in first parity sows and its effect on second litter size. Livest. Sci. 2010, 132, 165–172. [Google Scholar] [CrossRef]
- Wijesiriwardana, U.A.; Craig, J.R.; Cottrell, J.J.; Dunshea, F.R.; Pluske, J.R. Animal board invited review: Factors affecting the early growth and development of gilt progeny compared to sow progeny. Animal 2022, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.B.; Carroll, J.A.; Yoon, I.; Mateo, R.D.; Kim, S.W. Effects of supplementing Saccharomyces cerevisiae fermentation product in sow diets on performance of sows and nursing piglets. J. Anim. Sci. 2011, 89, 2462–2471. [Google Scholar] [CrossRef] [PubMed]
- Perricone, V.; Sandrini, S.; Irshad, N.; Savoini, G.; Comi, M.; Agazzi, A. Yeast-Derived Products: The Role of Hydrolyzed Yeast and Yeast Culture in Poultry Nutrition—A Review. Animals 2022, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.H.; Garrett, J.E. Effect of yeast culture (Saccharomyces cerevisiae) on adaptation of cows to postpartum diets and on lactational performance. J. Anim. Sci. 1999, 77, 988–999. [Google Scholar] [CrossRef]
- Geng, C.Y.; Ren, L.P.; Zhou, Z.M.; Chang, Y.; Meng, Q.X. Comparison of active dry yeast (Saccharomyces cerevisiae) and yeast culture for growth performance, carcass traits, meat quality and blood indexes in finishing bulls. Anim. Sci. J. 2016, 87, 982–988. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Q.B.; Zhou, P.; Li, Z.; Zhong, W.; Zhuo, Y.; Che, L.Q.; Xu, S.Y.; Fang, Z.F.; Jiang, X.M.; et al. Effects of yeast culture supplementation from late gestation to weaning on performance of lactating sows and growth of nursing piglets. Animal 2022, 16, 8. [Google Scholar] [CrossRef] [PubMed]
- NRC; National Research Council. The Nutrient Requirements of Swine; Eighth Revised Edition; National Academy Press: Washington, DC, USA, 2012.
- AOAC. Official Methods of Analysis, 18th ed.; AOAC Int.: Arlington, VA, USA, 2006. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Zhe, L.; Zhou, R.; Theil, P.K.; Krogh, U.; Yang, L.X.; Zhuo, Y.; Lin, Y.; Xu, S.Y.; Jiang, X.M.; Huang, L.J.; et al. The Impact of Enhancing Diet Quality or Dietary Supplementation of Flavor and Multi-Enzymes on Primiparous Lactating Sows. Animals 2022, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.W.; Luo, Z.; Cao, C.Y.; Sun, S.S.; Ma, Q.Q.; Li, Z.Y.; Shi, B.M.; Shan, A.S. Weaning Alters Intestinal Gene Expression Involved in Nutrient Metabolism by Shaping Gut Microbiota in Pigs. Front. Microbiol. 2020, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.K.; Fang, J.B.; Qi, X.J.; Lin, M.M.; Zhong, Y.P.; Sun, L.M.; Cui, W. Combined Analysis of the Fruit Metabolome and Transcriptome Reveals Candidate Genes Involved in Flavonoid Biosynthesis in Actinidia arguta. Int. J. Mol. Sci. 2018, 19, 17. [Google Scholar] [CrossRef]
- Theil, P.K.; Farmer, C.; Feyera, T. Review: Physiology and nutrition of late gestating and transition sows. J. Anim. Sci. 2022, 100, 15. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Lang, M.; Zhen, Y.G.; Chen, X.; Sun, Z.; Zhao, W.; Zhang, X.F.; Wang, T.; Qin, G.X. Effects of yeast culture supplementation and the ratio of non-structural carbohydrate to fat on growth performance, carcass traits and the fatty acid profile of the longissimus dorsi muscle in lambs. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1274–1282. [Google Scholar] [CrossRef]
- Dong, L.; Li, Y.; Zhang, Y.; Zhang, Y.; Ren, J.; Zheng, J.; Diao, J.; Ni, H.; Yin, Y.; Sun, R.; et al. Effects of organic zinc on production performance, meat quality, apparent nutrient digestibility and gut microbiota of broilers fed low-protein diets. Sci. Rep. 2023, 13, 10803. [Google Scholar] [CrossRef]
- Jacyno, E.; Pietruszka, A.; Biel, W.; Kolodziej-Skalska, A.; Matysiak, B.; Kawecka, M.; Sosnowska, A. Effect of sow age on the apparent total tract digestibility of nutrients in the diet. S. Afr. J. Anim. Sci. 2016, 46, 230–236. [Google Scholar] [CrossRef]
- Miller, M.B.; Hartsock, T.G.; Erez, B.; Douglass, L.; Alston-Mills, B. Effect of dietary calcium concentrations during gestation and lactation in the sow on milk composition and litter growth. J. Anim. Sci. 1994, 72, 1315–1319. [Google Scholar] [CrossRef]
- Conejos, J.R.V.; Acda, S.P.; Capitan, S.S.; Agbisit, E.M.; Merca, F.E. Mannan Oligosaccharides from Yeast (Saccharomyces cerevisiae) Cell Wall Improves Nutrient Digestibility and Intestinal Morphology of Growing Pigs Sus domesticus (Erxleben). Philipp. Agric. Sci. 2012, 95, 305–311. [Google Scholar]
- Kim, J.S.; Ingale, S.L.; Hosseindoust, A.R.; Lee, S.H.; Lee, J.H.; Chae, B.J. Effects of mannan level and β-mannanase supplementation on growth performance, apparent total tract digestibility and blood metabolites of growing pigs. Animal 2017, 11, 202–208. [Google Scholar] [CrossRef]
- Kornegay, E.T.; Rhein-Welker, D.; Lindemann, M.D.; Wood, C.M. Performance and nutrient digestibility in weanling pigs as influenced by yeast culture additions to starter diets containing dried whey or one of two fiber sources. J. Anim. Sci. 1995, 73, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Veum, T.L.; Reyes, J.; Ellersieck, M. Effect of supplemental yeast culture in sow gestation and lactation diets on apparent nutrient digestibilities and reproductive performance through one reproductive cycle. J. Anim. Sci. 1995, 73, 1741–1745. [Google Scholar] [CrossRef]
- Vasquez, R.; Oh, J.K.; Song, J.H.; Kang, D.K. Gut microbiome-produced metabolites in pigs: A review on their biological functions and the influence of probiotics. J. Anim. Sci. Technol. 2022, 64, 671–695. [Google Scholar] [CrossRef]
- Zietek, M.; Celewicz, Z.; Kikut, J.; Szczuko, M. Implications of SCFAs on the Parameters of the Lipid and Hepatic Profile in Pregnant Women. Nutrients 2021, 13, 13. [Google Scholar] [CrossRef]
- Sun, B.; Vatanen, T.; Jayasinghe, T.N.; McKenzie, E.; Murphy, R.; O’Sullivan, J.M. Desacetyl-alpha-MSH and alpha-MSH have sex specific interactions with diet to influence mouse gut morphology, metabolites and microbiota. Sci. Rep. 2020, 10, 11. [Google Scholar] [CrossRef]
- Rook, J.A.; Balch, C.C.; Johnson, V.W. Further observations on the effects of intraruminal infusions of volatile fatty acids and of lactic acid on the yield and composition of the milk of the cow. Br. J. Nutr. 1965, 19, 93–99. [Google Scholar] [CrossRef]
- Shen, Y.B.; Fellner, V.; Yoon, I.; Kim, S.W. Effects of dietary supplementation of Saccharomyces cerevisiae fermentation product to sows and their offspring on growth and meat quality. Transl. Anim. Sci. 2017, 1, 45–53. [Google Scholar] [CrossRef]
- Wei, X.X.; Li, L.X.; Yan, H.S.; Li, Q.H.; Gao, J.J.; Hao, R.R. Grape seed procyanidins improve intestinal health by modulating gut microbiota and enhancing intestinal antioxidant capacity in weaned piglets. Livest. Sci. 2022, 264, 13. [Google Scholar] [CrossRef]
- Wang, W.W.; Li, Z.I.; Han, Q.Q.; Guo, Y.M.; Zhang, B.; D’Inca, R. Dietary live yeast and mannan-oligosaccharide supplementation attenuate intestinal inflammation and barrier dysfunction induced by Escherichia coli in broilers. Br. J. Nutr. 2016, 116, 1878–1888. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.H.; Li, H.; Luo, J.Q.; Zhang, K.Y. Yeast-Derived beta-1,3-Glucan Substrate Significantly Increased the Diversity of Methanogens During In vitro Fermentation of Porcine Colonic Digesta. J. Integr. Agric. 2013, 12, 2229–2234. [Google Scholar] [CrossRef]
- Leser, T.D.; Molbak, L. Better living through microbial action: The benefits of the mammalian gastrointestinal microbiota on the host. Environ. Microbiol. 2009, 11, 2194–2206. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Cheng, X.; Zhen, W.R.; Zeng, D.; Qu, L.J.; Wang, Z.; Ning, Z.H. Yeast Culture Improves Egg Quality and Reproductive Performance of Aged Breeder Layers by Regulating Gut Microbes. Front. Microbiol. 2021, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.G.; Zhang, S.S.; Nie, Q.X.; He, H.J.; Tan, H.Z.; Geng, F.; Ji, H.H.; Hu, J.L.; Nie, S.P. Gut firmicutes: Relationship with dietary fiber and role in host homeostasis. Crit. Rev. Food Sci. Nutr. 2022, 16, 1–16. [Google Scholar] [CrossRef]
- Vander Wyst, K.B.; Ortega-Santos, C.P.; Toffoli, S.N.; Lahti, C.E.; Whisner, C.M. Diet, adiposity, and the gut microbiota from infancy to adolescence: A systematic review. Obes. Rev. 2021, 22, 21. [Google Scholar] [CrossRef]
- Precup, G.; Vodnar, D.C. Gut Prevotella as a possible biomarker of diet and its eubiotic versus dysbiotic roles: A comprehensive literature review. Br. J. Nutr. 2019, 122, 131–140. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 25. [Google Scholar] [CrossRef]
- Niu, J.K.; Liu, X.; Xu, J.Y.; Li, F.; Wang, J.C.; Zhang, X.X.; Yang, X.; Wang, L.; Ma, S.; Li, D.F.; et al. Effects of Silage Diet on Meat Quality through Shaping Gut Microbiota in Finishing Pigs. Microbiol. Spectr. 2022, 19, e02416-22. [Google Scholar] [CrossRef]
- Dias, A.L.G.; Freitas, J.A.; Micai, B.; Azevedo, R.A.; Greco, L.F.; Santos, J.E.P. Effects of supplementing yeast culture to diets differing in starch content on performance and feeding behavior of dairy cows. J. Dairy Sci. 2018, 101, 186–200. [Google Scholar] [CrossRef]
- Theil, P.K.; Lauridsen, C.; Quesnel, H. Neonatal piglet survival: Impact of sow nutrition around parturition on fetal glycogen deposition and production and composition of colostrum and transient milk. Animal 2014, 8, 1021–1030. [Google Scholar] [CrossRef]
- Quesnel, H.; Farmer, C.; Devillers, N. Colostrum intake: Influence on piglet performance and factors of variation. Livest. Sci. 2012, 146, 105–114. [Google Scholar] [CrossRef]
- Szyndler-Nedza, M.; Rozycki, M.; Eckert, R.; Mucha, A.; Koska, M.; Szulc, T. Relationships between chemical composition of colostrum and milk and rearing performance of piglets during a 21-day lactation. Ann. Anim. Sci. 2013, 13, 771–781. [Google Scholar] [CrossRef]
- Han, X.S.; Gao, Y.; Li, G.L.; Xiong, Y.C.; Zhao, C.Z.; Ruan, J.X.; Ma, Y.L.; Li, X.Y.; Li, C.C.; Zhao, S.H.; et al. Enhancing the antibacterial activities of sow milk via site-specific knock-in of a lactoferrin gene in pigs using CRISPR/Cas9 technology. Cell Biosci. 2020, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Spry, C.; Kirk, K.; Saliba, K.J. Coenzyme A biosynthesis: An antimicrobial drug target. FEMS Microbiol. Rev. 2008, 32, 56–106. [Google Scholar] [CrossRef] [PubMed]
- Stothers, S.C.; Schmidt, D.A.; Johnston, R.L.; Hoefer, J.A.; Luecke, R.W. The pantothenic acid requirement of the baby pig. J. Nutr. 1955, 57, 47–53. [Google Scholar] [CrossRef]
- Owen, B.D.; Bowland, J.P. The pantothenic acid content of the blood and milk of swine fed supplemental levels of the vitamin. J. Nutr. 1952, 48, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Christgen, S.L.; Becker, D.F. Role of Proline in Pathogen and Host Interactions. Antioxid. Redox Signal. 2019, 30, 683–709. [Google Scholar] [CrossRef]
- Tanner, J.J. Structural biology of proline catabolism. Amino Acids 2008, 35, 719–730. [Google Scholar] [CrossRef]
- Gu, C.S.; Mao, X.B.; Chen, D.W.; Yu, B.; Yang, Q. Isoleucine Plays an Important Role for Maintaining Immune Function. Curr. Protein Pept. Sci. 2019, 20, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Munro, H.N. Absorption and metabolism of amino acids with special emphasis on phenylalanine. J. Toxicol. Environ. Health 1976, 2, 189–206. [Google Scholar] [CrossRef]
- Giudetti, A.M.; Stanca, E.; Siculella, L.; Gnoni, G.V.; Damiano, F. Nutritional and Hormonal Regulation of Citrate and Carnitine/Acylcarnitine Transporters: Two Mitochondrial Carriers Involved in Fatty Acid Metabolism. Int. J. Mol. Sci. 2016, 17, 15. [Google Scholar] [CrossRef]
- Engel, P.; Dzapo, V. Influence of endogenous carnitine in blood plasma and lymphocytes on reproduction performance of sows. Tieraerztl. Prax. Ausg. Grosstiere Nutztiere 2010, 38, 278–284. [Google Scholar] [CrossRef]
- Alugongo Gibson, M.; Xiao, J.; Wu, Z.; Li, S.; Wang, Y.; Cao, Z. Review: Utilization of yeast of Saccharomyces cerevisiae origin in artificially raised calves. J. Anim. Sci. Biotechnol. 2017, 8, 549–560. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Items | Gestation | Lactation |
|---|---|---|
| Ingredients, % | ||
| Corn | 55.81 | 61.24 |
| Wheat bran | 9.23 | 5.00 |
| Soybean meal | 16.38 | 21.70 |
| Expanded soybean | 14.63 | − |
| Fish meal | − | 2.50 |
| Soybean oil | − | 5.00 |
| L-lysine-HCL | 0.04 | 0.27 |
| DL-methionine | − | 0.12 |
| Chloride choline | 0.15 | 0.15 |
| NaCL | 0.40 | 0.40 |
| Calcium carbonate | 0.16 | 0.17 |
| Dicalcium phosphate | 1.20 | 1.45 |
| Vitamin and mineral premix 1 | 2.00 | 2.00 |
| Total | 100.00 | 100.00 |
| Nutrient levels 2 | ||
| Net energy, MJ/kg | 9.88 | 9.91 |
| Crude protein | 16.89 | 16.65 |
| Calcium | 0.86 | 0.92 |
| Phosphorus | 0.65 | 0.63 |
| Lysine | 0.73 | 1.11 |
| Methionine | 0.66 | 0.61 |
| Items | CON | YC | p-Value |
|---|---|---|---|
| Litter size at birth, heads | 12.23 ± 2.16 | 12.45 ± 2.37 | 0.717 |
| Number of piglets born alive, heads | 11.93 ± 2.18 | 12.34 ± 2.33 | 0.487 |
| Litter birth weight, kg | 16.97 ± 3.29 | 18.47 ± 3.39 | 0.271 |
| Piglet birth weight, kg | 1.42 ± 0.04 | 1.49 ± 0.07 | 0.046 |
| Stillborn piglets, heads | 0.30 ± 0.53 | 0.10 ± 0.31 | 0.091 |
| Piglets < 1.0 kg, heads | 0.90 ± 0.96 | 0.52 ± 0.57 | 0.069 |
| Number of weaned piglets, heads | 11.03 ± 2.31 | 11.86 ± 2.13 | 0.158 |
| Litter weaning weight, kg | 87.18 ± 4.26 | 89.68 ± 6.58 | 0.648 |
| Piglet weaning weight, kg | 7.39 ± 0.19 | 7.53 ± 0.19 | 0.155 |
| ADFI during lactation 2, kg/d | 5.12 ± 0.23 | 5.10 ± 0.47 | 0.302 |
| Body weight at 1 d of lactation, kg | 138.4 ± 10.52 | 140.2 ± 9.44 | 0.776 |
| Body weight at 28 d of lactation, kg | 132.8 ± 9.08 | 134.8 ± 9.69 | 0.142 |
| Loss of body weight during lactation, kg | 6.30 ± 0.66 | 5.48 ± 0.60 | 0.755 |
| Backfat thickness at 1 d of lactation, mm | 18.12 ± 1.43 | 18.18 ± 1.22 | 0.395 |
| Backfat thickness at 28 d of lactation, mm | 17.84 ± 1.28 | 18.93 ± 1.04 | 0.031 |
| Loss of BF during lactation 3, mm | 0.30 ± 0.02 | 0.27 ± 0.03 | 0.255 |
| Days to return to estrus, d | 5.41 ± 0.91 | 4.90 ± 0.72 | 0.862 |
| Items (%) | CON | YC | p-Value |
|---|---|---|---|
| Crude ash | 76.24 ± 0.44 | 75.22 ± 0.59 | 0.388 |
| Crude fiber | 13.36 ± 0.63 | 12.51 ± 0.47 | 0.361 |
| Gross energy | 83.10 ± 0.22 | 84.20 ± 0.26 | 0.049 |
| Crude protein | 81.60 ± 0.22 | 82.31 ± 0.42 | 0.302 |
| Phosphorus | 40.53 ± 0.44 | 40.84 ± 0.60 | 0.054 |
| Calcium | 70.12 ± 0.55 | 73.44 ± 0.72 | 0.032 |
| Items | CON | YC | p-Value |
|---|---|---|---|
| Colostrum | |||
| Fat, % | 5.03 ± 0.16 | 5.19 ± 0.10 | 0.046 |
| Protein, % | 15.75 ± 0.33 | 15.92 ± 0.53 | 0.345 |
| Lactose, % | 3.47 ± 0.67 | 3.94 ± 0.77 | 0.027 |
| Dry matter, % | 28.04 ± 0.40 | 28.11 ± 0.77 | 0.804 |
| Milk | |||
| Fat, % | 5.02 ± 0.11 | 5.12 ± 0.63 | 0.055 |
| Protein, % | 5.72 ± 0.37 | 5.81 ± 0.25 | 0.444 |
| Lactose, % | 5.21 ± 0.71 | 5.39 ± 0.11 | 0.060 |
| Dry matter, % | 19.67 ± 0.51 | 20.04 ± 0.73 | 0.163 |
| Metabolite | Class | Ratio 1 | p-Value | VIP 2 | MZ 3 | RT 4 |
|---|---|---|---|---|---|---|
| Styrene oxide | Benzenoids | 0.05 | 7.70 × 10−5 | 1.96 | 121.08 | 1.41 |
| Pantothenic acid | Organic acids and derivatives | 60.50 | 1.03 × 10−6 | 2.33 | 220.13 | 3.18 |
| Ramiprilat | Organic acids and derivatives | 24.57 | 1.14 × 10−6 | 2.04 | 387.19 | 3.25 |
| Seryltryptophan | Organic acids and derivatives | 49.14 | 1.93 × 10−7 | 2.25 | 290.11 | 3.36 |
| Pyroglutamylvaline | Organic acids and derivatives | 14.82 | 6.94 × 10−7 | 1.90 | 227.10 | 3.34 |
| Proline | Organic acids and derivatives | 15.99 | 6.05 × 10−3 | 2.14 | 116.07 | 3.31 |
| Isoleucine | Organic acids and derivatives | 17.67 | 3.17 × 10−6 | 1.91 | 132.10 | 3.23 |
| Iminodiacetic acid | Organic acids and derivatives | 31.52 | 3.65 × 10−3 | 1.77 | 134.04 | 3.74 |
| Methionine | Organic acids and derivatives | 0.15 | 2.98 × 10−7 | 1.68 | 150.06 | 1.23 |
| Phenylalanine | Organic acids and derivatives | 8.55 | 4.39 × 10−5 | 1.69 | 166.09 | 3.54 |
| Pyridoxamine | Organoheterocyclic compounds | 6.63 | 2.36 × 10−5 | 1.61 | 169.10 | 3.15 |
| Caffeoylcholine | Unknown | 112.25 | 1.01 × 10−8 | 2.50 | 266.16 | 3.41 |
| Linsidomine cation | Unknown | 22.39 | 6.94 × 10−4 | 2.14 | 189.12 | 2.31 |
| Imazamox | Unknown | 57.85 | 4.80 × 10−6 | 2.32 | 304.15 | 3.19 |
| Pilocarpine | Alkaloids and derivatives | 102.54 | 1.52 × 10−3 | 2.34 | 226.15 | 1.86 |
| Acylcarnitine 4:0 | Lipids and lipid-like molecules | 100.27 | 2.84 × 10−8 | 2.52 | 232.17 | 3.33 |
| Acylcarnitine 5:0 | Lipids and lipid-like molecules | 76.79 | 2.16 × 10−8 | 2.46 | 246.19 | 3.45 |
| Daidzein | Phenylpropanoids and polyketides | 6.59 | 1.06 × 10−5 | 1.65 | 255.06 | 3.97 |
| Genistein | Phenylpropanoids and polyketides | 9.36 | 1.15 × 10−5 | 1.80 | 271.06 | 4.17 |
| Biochanin A | Phenylpropanoids and polyketides | 7.04 | 2.42 × 10−5 | 1.65 | 285.07 | 4.01 |
| Gerberinol | Phenylpropanoids and polyketides | 0.12 | 9.03 × 10−6 | 1.84 | 365.10 | 3.32 |
| Apigenin | Phenylpropanoids and polyketides | 9.24 | 3.32 × 10−5 | 1.78 | 269.04 | 4.18 |
| Hexaethylene glycol | Organic oxygen compounds | 11.15 | 1.42 × 10−5 | 1.56 | 300.19 | 3.31 |
| Heliannuol C | Organoheterocyclic compounds | 0.22 | 1.80 × 10−4 | 1.56 | 307.15 | 0.69 |
| Piperidine | Organoheterocyclic compounds | 9.14 | 2.04 × 10−5 | 1.61 | 86.10 | 1.76 |
| Xanthine | Organoheterocyclic compounds | 11.37 | 1.36 × 10−3 | 1.75 | 151.02 | 1.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Wu, Z.; Wang, Y.; Meng, Q.; Chen, P.; Li, J.; Shan, A. Effect of Yeast Culture on Reproductive Performance, Gut Microbiota, and Milk Composition in Primiparous Sows. Animals 2023, 13, 2954. https://doi.org/10.3390/ani13182954
Ma Z, Wu Z, Wang Y, Meng Q, Chen P, Li J, Shan A. Effect of Yeast Culture on Reproductive Performance, Gut Microbiota, and Milk Composition in Primiparous Sows. Animals. 2023; 13(18):2954. https://doi.org/10.3390/ani13182954
Chicago/Turabian StyleMa, Zhizhuo, Ze Wu, Yu Wang, Qingwei Meng, Peng Chen, Jianping Li, and Anshan Shan. 2023. "Effect of Yeast Culture on Reproductive Performance, Gut Microbiota, and Milk Composition in Primiparous Sows" Animals 13, no. 18: 2954. https://doi.org/10.3390/ani13182954