
Effect of Chitosan on Ruminal Fermentation and Microbial Communities, Methane Emissions, and Productive Performance of Dairy Cattle
 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Diets
2.2. Measurements and Samplings
2.3. Handling and Laboratory Procedures
2.3.1. Feed
2.3.2. Milk
2.3.3. Volatile Fatty Acid Determinations
2.3.4. Blood
2.3.5. Purine Derivative Determinations
2.3.6. DNA Extraction and Sequencing of Rumen Samples
2.3.7. Bioinformatics
2.4. Calculations and Statistical Analysis
3. Results
3.1. VFA Profile
3.2. Milk Yield, Milk Composition and Methane Production
3.3. Blood Parameters and N Flux
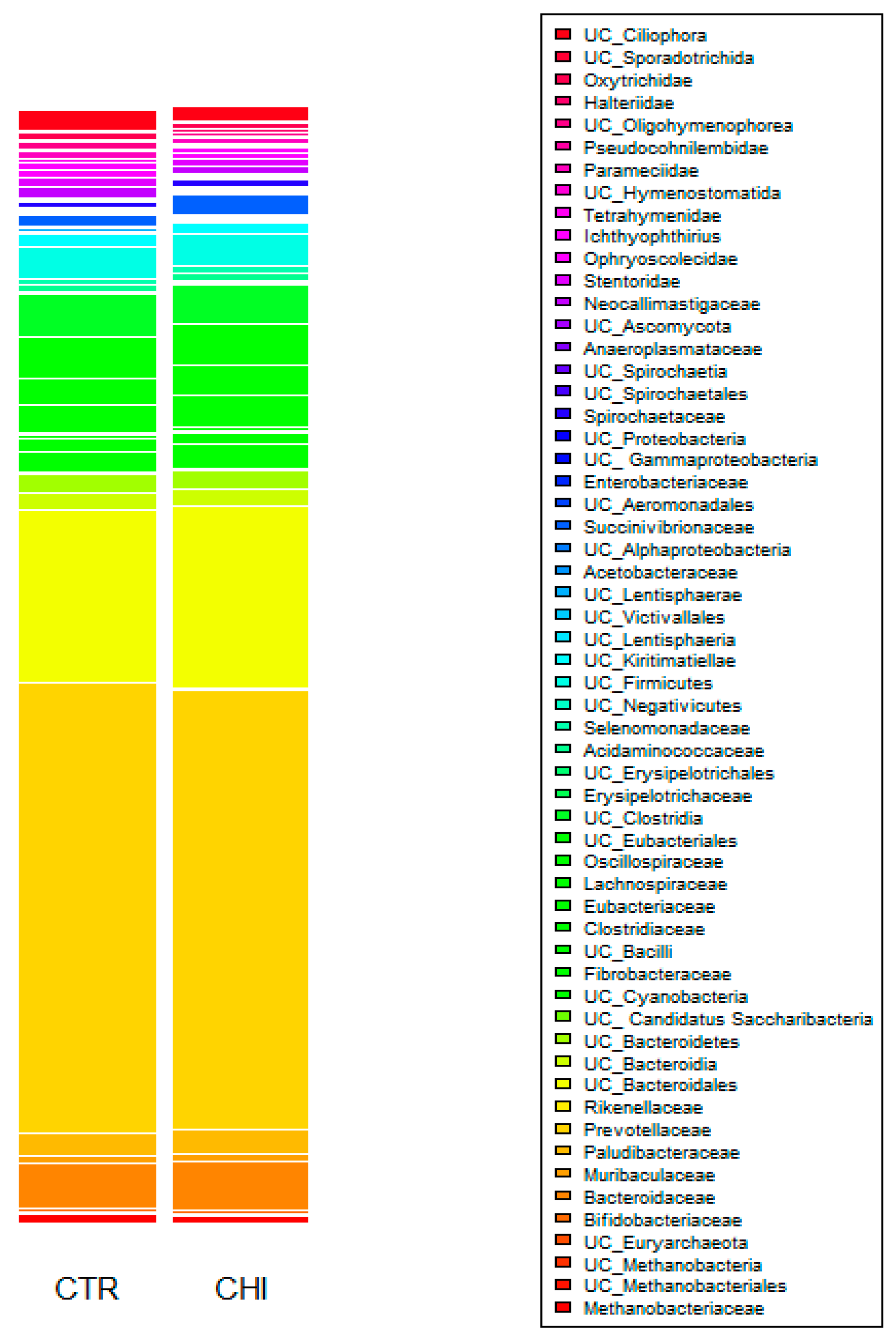
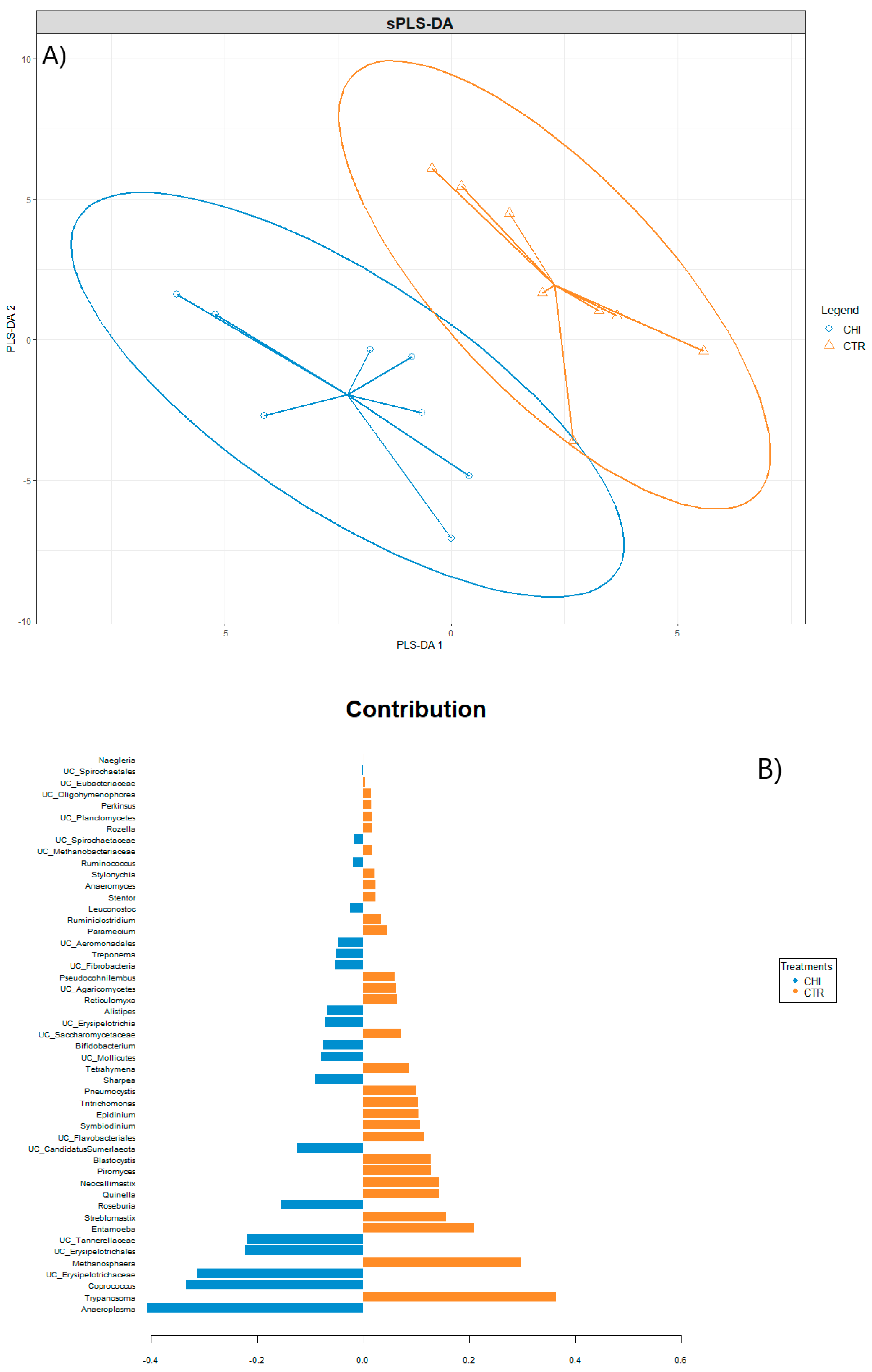
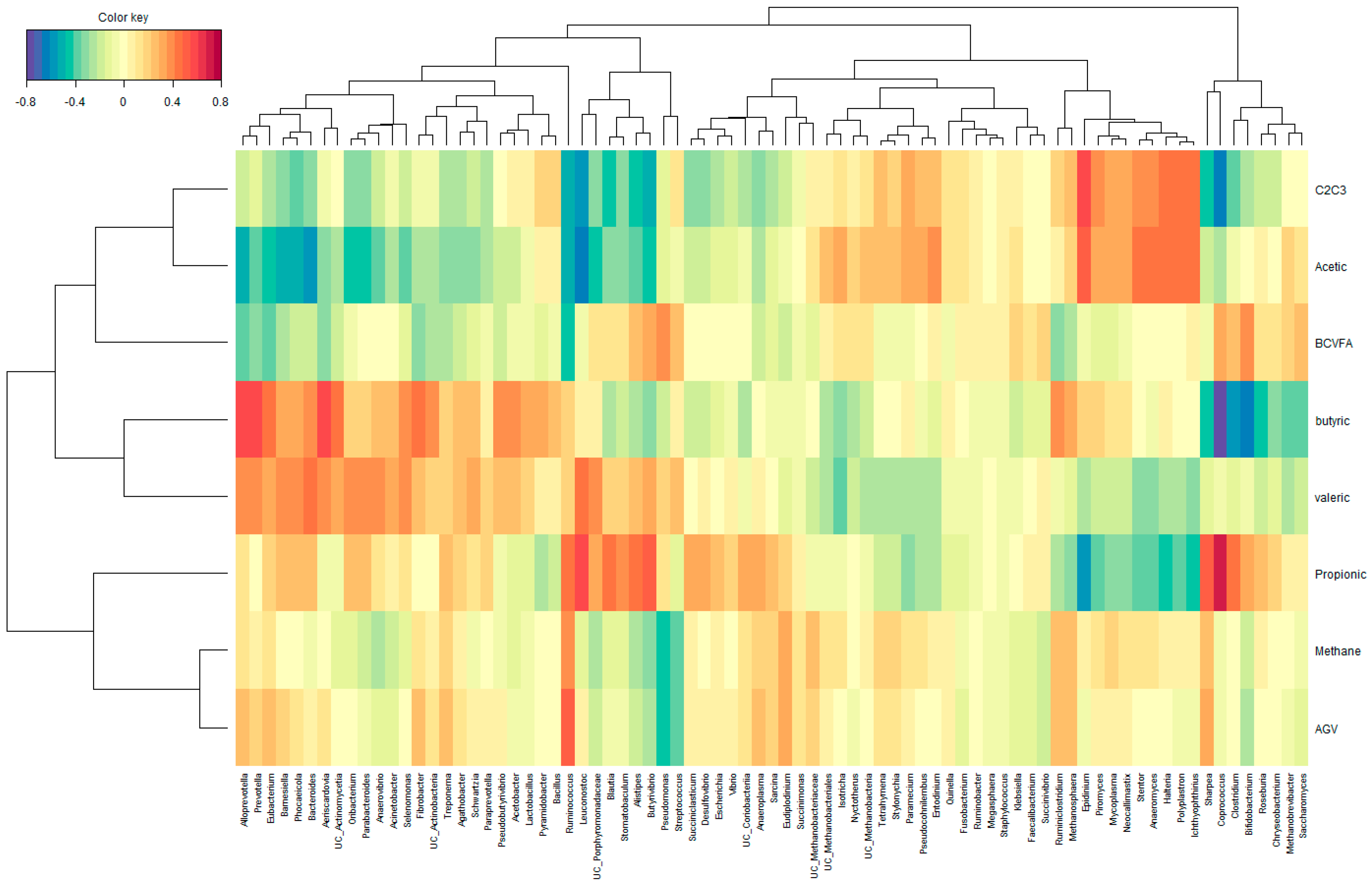
3.4. Ruminal Microbial Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McAllister, T.A.; Newbold, C.J. Redirecting methane to reduce methanogenesis. Aust. J. Exp. Agric. 2008, 48, 7–13. [Google Scholar] [CrossRef]
- Forster, P.; Storelvmo, T.; Armour, K.; Collins, W.; Dufresne, J.L.; Frame, D.; Lunt, D.; Mauritsen, T.; Palmer, M.; Watanabe, M.; et al. The Earth’s energy budget, climate feedbacks, and climate sensitivity. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; p. U13B-07. [Google Scholar]
- Janssen, P.H. Influence of hydrogen on rumen methane formation and fermentation balances through microbial growth kinetics and fermentation thermodynamics. Anim. Feed Sci. Technol. 2010, 160, 1–22. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef]
- Grossi, G.; Goglio, P.; Vitali, A.; Williams, A.G. Livestock and climate change: Analysis livestock on climate and mitigation strategies. Anim. Front. 2009, 9, 69–76. [Google Scholar] [CrossRef]
- Hungate, R.E.; Smith, W.; Bauchop, T.; Yu, I. Rabinowitz JC. Formate as an intermediate in the bovine rumen fermentation. J. Bacteriol. 1970, 102, 389–397. [Google Scholar] [CrossRef]
- Russell, J.B.; Wallace, R.J. Energy-yielding and energy-consuming reactions. In The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Springer: Dordrecht, The Netherlands, 1997; pp. 10–72. [Google Scholar] [CrossRef]
- Wedlock, D.N.; Pedersen, G.; Denis, M.; Buddle, B.M.; Dey, D.; Janssen, P.H. Development of a vaccine to mitigate greenhouse gas emissions in análisis: Vaccination of sheep with methanogen fractions induces antibodies that block methane production in vitro. N. Z. Vet. J. 2010, 58, 29–36. [Google Scholar] [CrossRef]
- Chaucheyras, F.; Fonty, G.; Bertin, G.; Gouet, P. In vitro H2 utilization by a ruminal acetogenic bacterium cultivated alone or in association with an archaea methanogen is stimulated by a probiotic strain of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1995, 61, 3466–3467. [Google Scholar] [CrossRef]
- Whitelaw, F.G.; Eadie, J.M.; Bruce, L.A.; Shand, W.J. Methane formation in faunated and ciliate-free cattle and its relationship with rumen volatile fatty acid proportions. Brit. J. Nutr. 1984, 62, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Itabashi, H.; Kobayashi, T.; Matsumoto, M. The effects of rumen ciliate protozoa on energy metabolism and some constituents in rumen fluid and blood plasma of goats. Jpn. J. Zootech. Sci. 1986, 55, 248–256. [Google Scholar]
- Van Nevel, C.J.; Demeyer, D.I. Control of rumen methanogenesis. Environ. Monit. Assess. 1996, 42, 73–97. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; McGinn, S.M.; Martinez, T.F.; McAllister, T.A. Use of condensed tannin extract from quebracho trees to reduce methane emissions from cattle. J. Anim. Sci. 2007, 85, 1990–1996. [Google Scholar] [CrossRef] [PubMed]
- Grainger, C.; Clarke, T.; Auldist, M.J.; Beauchemin, K.A.; McGinn, S.M.; Waghorn, G.C.; Eckard, R.J. Potential use of acacia mearnsii condensed tannins to reduce methane emissions and nitrogen excretion from grazing dairy cows. Can. J. Anim. Sci. 2009, 89, 241–251. [Google Scholar] [CrossRef]
- Goel, G.; Makkar, H.P.S. Methane mitigation from ruminants using tannins and saponins. Trop. Anim. Health Prod. 2012, 44, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Eugène, M.; Masse, D.; Chiquette, J.; Benchaar, C. Meta-analysis on the effects of lipid supplementation on methane production in lactating dairy cows. Can. J. Anim. Sci. 2008, 88, 331–334. [Google Scholar] [CrossRef]
- Martínez-Álvaro, M.; Auffret, M.D.; Stewart, R.D.; Dewhurst, R.J.; Duthie, C.A.; Rooke, J.A.; Roehe, R. Identification of complex rumen microbiome interaction within diverse functional niches as mechanisms affecting the variation of methane emissions in bovine. Front. Microbiol. 2020, 11, 659. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A. Chitin Enzymology; Alda Tecnografica: Ancona, Italy, 1993. [Google Scholar]
- Young, D.H.; Kohle, H.; Kauss, H. Effect of chitosan on membrane permeability of suspension-cultured Glycine max and Phaseolus vulgaris cell. Plant Physiol. 1982, 70, 1449–1454. [Google Scholar] [CrossRef]
- Matsuhashi, S.; Kume, T. Enhancement of the antimicrobial activity of chitosan by irradiation. J. Sci. Food Agric. 1999, 73, 237–241. [Google Scholar] [CrossRef]
- Goiri, I.; Garcia-Rodriguez, A.; Oregui, L.M. Effect of chitosans on in vitro rumen digestion and fermentation of maize silage. Anim. Feed Sci. Technol. 2009, 148, 276–287. [Google Scholar] [CrossRef]
- Goiri, I.; Oregui, L.M.; Garcia-Rodriguez, A. Use of chitosans to modulate ruminal fermentation of a 50:50 forage-to-concentrate diet in sheep. J. Anim. Sci. 2010, 88, 749–755. [Google Scholar] [CrossRef]
- Dias, A.O.C.; Goes, R.H.T.B.; Gandra, J.R.; Takiya, C.S.; Branco, A.F.; Jacaúna, A.G.; Oliveira, R.T.; Souza, C.J.S.; Vaz, M.S.M. Increasing doses of chitosan to grazing beef steers: Nutrient intake and digestibility, ruminal fermentation, and nitrogen utilization. Anim. Feed Sci. Technol. 2017, 225, 73–80. [Google Scholar] [CrossRef]
- De Paiva, P.G.; de Jesus, E.F.; Del Valle, T.A.; de Almeida, G.F.; Costa, A.G.B.V.B.; Consentini, C.E.C.; Zanferari, F.; Takiya, C.S.; da Silva Bueno, I.C.; Rennó, F.P. Effects of chitosan on ruminal fermentation, nutrient digestibility, and milk yield and composition of dairy cows. Anim. Prod. Sci. 2017, 57, 301. [Google Scholar] [CrossRef]
- Vendramini, T.H.A.; Takiya, C.S.; Silva, T.H.; Zanferari, F.; Rentas, M.F.; Bertoni, J.C.; Consentini, C.E.C.; Gardinal, R.; Acedo, T.S.; Rennó, F.P. Effects of a blend of essential oils, chitosan or monensin on nutrientintake and digestibility of lactating dairy cows. Anim. Feed Sci. Technol. 2016, 214, 12–21. [Google Scholar] [CrossRef]
- Belanche, A.; Pinloche, E.; Preskett, D.; Newbold, C.A. Effects and mode of action of chitosan and ivy fruit saponins on the microbiome, fermentation and methanogenesis in the rumen simulation technique. FEMS Microbiol. Ecol. 2016, 92, fiv160. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodriguez, A.; Arranz, J.; Mandaluniz, N.; Beltrán-de-Heredia, I.; Ruiz, R.; Goiri, I. Short-communication: Production performance and plasma metabolites of dairy ewes in early lactation as affect by chitosan. Span. J. Agric. Res. 2015, 13, e06SC04. [Google Scholar] [CrossRef]
- Del Valle, T.A.; de Paiva, P.G.; de Jesus, E.F.; de Almeida, G.F.; Zanferari, F.; Costa, A.G.B.V.B.; Bueno, I.C.S.; Rennó, F.P. Dietary chitosan improves nitrogen use and feed conversion in diets for mid-lactation dairy cows. Livest. Sci. 2017, 201, 22–29. [Google Scholar] [CrossRef]
- Zanferari, F.; Vendramini, T.H.A.; Rentas, M.F.; Gardinal, R.; Calomeni, G.D.; Mesquita, L.G.; Takiya, C.S.; Rennó, F.P. Effects of chitosan and whole raw soybeans on ruminal fermentation and bacterial populations, and milk fatty acid profile in dairy cows. J. Dairy Sci. 2018, 101, 10939–10952. [Google Scholar] [CrossRef]
- Goiri, I.; Oregui, L.M.; Garcia-Rodriguez, A. Dose-response effects of chitosans on in vitro rumen digestion and fermentation of mixtures differing in forage-to-concentrate ratios. Anim. Feed Sci. Technol. 2009, 151, 215–227. [Google Scholar] [CrossRef]
- Henry, D.D.; Ruiz-Moreno, M.; Ciriaco, F.M.; Kohmann, M.; Mercadante, V.R.; Lamb, G.C.; DiLorenzo, N. Effects of chitosan on nutrient digestibility, methane emissions, and in vitro fermentation in beef cattle. J. Anim. Sci. 2015, 93, 3539–3550. [Google Scholar] [CrossRef]
- Vyniard, J.R.; Faciola, A.P. Unraveling the pros and cons of various in vitro methodologies for ruminant nutrition: A review. Trans. Anim. Sci. 2022, 6, taxac130. [Google Scholar] [CrossRef]
- Tong, J.; Zhang, H.; Wang, J.; Liu, Y.; Mao, S.; Xiong, B.; Jiang, L. Effects of different molecular weights of chitosan on methane production and bacterial community structure in vitro. J. Integr. Agric. 2020, 19, 1644–1655. [Google Scholar] [CrossRef]
- López-Paredes, J.; Goiri, I.; Atxaerandio, R.; García-Rodríguez, A.; Ugarte, E.; Jiménez-Montero, J.A.; González-Recio, O. Mitigation of greenhouse gases in dairy cattle via genetic selection: 1. Genetic parameters of direct methane using noninvasive methods and proxies of methane. J. Dairy Sci. 2020, 103, 7199–7209. [Google Scholar] [CrossRef]
- Rey, J.; Atxaerandio, R.; Ruiz, R.; Ugarte, E.; González-Recio, O.; Garcia-Rodríguez, A.; Goiri, I. Comparison between non-invasive methane measurement techniques in cattle. Animals 2019, 9, 563. [Google Scholar] [CrossRef]
- AOAC. Association of Official Analytical Chemists. In Official Methods of Analysis; AOAC International: Rockville, MD, USA, 1996. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.B.; Van Soest, P. The detergent system of analisys. In The Analysis of Dietary Fibre in Food; James, W.P.T., Theander, O., Eds.; Marcel Dekker: New York, NY, USA, 1981; pp. 123–158. [Google Scholar]
- MAPA. Métodos Oficiales del MAPA; BOE: Madrid, Spain, 1995; Volume 52, p. 7167. [Google Scholar]
- Karkalas, J. An improved enzymic method for the determination of native and modified starch. J. Sci. Food Agric. 1985, 36, 1019–1027. [Google Scholar] [CrossRef]
- Reynal, S.M.; Broderick, G.A. A new HPLC purine assay for quantifying microbial. J. Dairy Sci. 2009, 92, 1177–1181. [Google Scholar] [CrossRef]
- Balcells, J.; Guada, J.A.; Peira, J.M.; Parker, D.S. Simultaneous determination of allantoin and oxypurines in biological fluids by high-performance liquid chromatography. J. Chromatogr. B Biomed. Appl. 1992, 575, 153–157. [Google Scholar] [CrossRef]
- Tamames, J.; Puente-Sánchez, F. SqueezeMeta, a highly portable, fully automatic metagenomic analysis pipeline. Front. Microbiol. 2019, 9, 3349. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Aitchison, J. The statistical analysis of compositional data. J. R. Stat. Soc. Ser. B 1982, 44, 139–177. Available online: https://www.jstor.org/stable/2345821 (accessed on 7 June 2023). [CrossRef]
- Greenacre, M. Compositional Data Analysis in Practice; Chapman & Hall/CRC: New York, NY, USA, 2018. [Google Scholar]
- Sorg, D.; Difford, G.F.; Mühlbach, S.; Kuhla, B.; Swalve, H.; Lassen, J.; Strabel, S.; Pszczola, M. Comparison of a laser methane detector with the GreenFeed and two breath analysers for on-farm measurements of methane emissions from dairy cows. Comput. Electron. Agric. 2018, 153, 285–294. [Google Scholar] [CrossRef]
- Broderick, G.A.; Merchen, N.R. Markers for quantifying microbial protein synthesis in the rumen. J. Dairy Sci. 1992, 75, 2618–2632. [Google Scholar] [CrossRef] [PubMed]
- Valadares, R.F.D.; Broderick, G.A.; Valadares Filho, S.C.; Clayton, M.K. Effect of replacing alfalfa silage with high moisture corn on ruminal protein synthesis estimated from excretion of total purine derivatives. J. Dairy Sci. 1999, 82, 2686–2696. [Google Scholar] [CrossRef] [PubMed]
- SAS. SAS Enterprise’s Guide; SAS: Cary, NC, USA, 2017. [Google Scholar]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Kim-Anh, L.C.; Rohart, F.; Gonzalez, I.S. Dejean with Key Contributors Benoit Gautier, Francois Bartolo, Contributions from Pierre Monget, Jeff Coquery, FangZou Yao and Benoit Liquet. mixOmics: Omics Data Integration Project. R Package Version 6.1.1. 2016. Available online: https://CRAN.R-project.org/package=mixOmics (accessed on 7 June 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 7 June 2023).
- Jouany, J.P.; Morgavi, D.P. Use of natural products as alternatives to antibiotic feed additives in ruminant production. Animal 2007, 1, 1443–1466. [Google Scholar] [CrossRef] [PubMed]
- Patra, A.K.; Saxena, J. Dietary phytochemicals as rumen modifiers: A review of the effects on microbial populations. Antonie Van Leeuwenhoek 2009, 96, 363–375. [Google Scholar] [CrossRef]
- Yuan, X.; Zheng, J.; Jiao, S.; Cheng, G.; Feng, C.; Du, Y.; Liu, H. A review on the preparation of chitosan oligosaccharides and application to human health, animal husbandry and agricultural production. Carbohydr. Polym. 2009, 220, 60–70. [Google Scholar] [CrossRef]
- Irawan, A.; Hidayat, C.; Jayanegara, A.; Ratriyanto, A. Essential oils as growth-promoting additives on performance, nutrient digestibility, cecal microbes, and serum metabolites of broiler chickens: A meta-analysis. Anim. Biosci. 2021, 34, 1499–1513. [Google Scholar] [CrossRef]
- Wina, E.; Muetzel, S.; Becker, K. The impact of saponins or saponin-containing plant materials on ruminant production—A review. J. Agric. Food Chem. 2005, 53, 8093–8105. [Google Scholar] [CrossRef]
- Franz, C.M.; Baser, K.H.C.; Hahn-Ramssl, I. Herbs and aromatic plants as feed additives: Aspects of composition, safety, and registration rules. In Feed Additives: Aromatic Plants and Herbs in Animal Nutrition and Health; Florou-Paneri, P., Christaki, E., Giannenas, I., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 35–56. [Google Scholar] [CrossRef]
- Kong, Y.; He, M.; McAlister, T.; Seviour, R.; Forster, R. Quantitative fluorescence in situ hybridization of microbial communities in the rumens of cattle fed different diets. Appl. Environ. Microbiol. 2010, 76, 6933–6938. [Google Scholar] [CrossRef]
- Zhan, J.; Liu, M.; Su, X.; Zhan, K.; Zhang, C.; Zhao, G. Effects of alfalfa flavonoids on the production performance, immune system, and ruminal fermentation of dairy cows. Asian-Australas. J. Anim. Sci. 2017, 30, 1416–1424. [Google Scholar] [CrossRef]
- Jiménez-Ocampo, R.; Valencia-Salazar, S.; Pinzón-Díaz, C.E.; Herrera-Torres, E.; Aguilar-Pérez, C.; Arango, J.; Ku-Vera, J.C. The role of chitosan as a possible agent for enteric methane mitigation in ruminants. Animals 2019, 9, 942. [Google Scholar] [CrossRef] [PubMed]
- Seankamsorn, A.; Cherdthong, A.; So, S.; Wanapat, M. Influence of chitosan sources on intake, digestibility, rumen fermentation, and milk production in tropical lactating dairy cows. Trop. Anim. Health Prod. 2021, 53, 241. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, mode of action, and in vivo activity of chitosan and its micro-and nanoparticles as antimicrobial agents: A review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.P.C.; Venturelli, B.C.; Santos, M.C.B.; Gardinal, R.; Cônsolo, N.R.B.; Calomeni, G.B.; Freitas, J.E.; Barletta, R.V.; Gandra, J.R.; Paiva, P.G. Chitosan affects total nutrient digestion and ruminal fermentationin Nellore steers. Anim. Feed Sci. Technol. 2015, 206, 114–118. [Google Scholar] [CrossRef]
- Harahap, R.P.; Setiawan, D.; Nahrowi, S.; Suharti, T.; Obitsu, A. Jayanegara. Enteric methane emissions and rumen fermentation profile treated by dietary chitosan: A meta-analysis of in vitro experiments. Trop. Anim. Sci. J. 2020, 43, 233–239. [Google Scholar] [CrossRef]
- Shah, A.M.; Qazi, I.H.; Matra, M.; Wanapat, M. Role of Chitin and chitosan in ruminant diets and their impact on digestibility, microbiota and performance of ruminants. Fermentation 2022, 8, 549. [Google Scholar] [CrossRef]
- Chung, Y.C.; Su, Y.P.; Chen, C.C.; Jia, G.; Wang, H.L.; Wu, J.C.; Lin, J.G. Relationship between antibacterial activity of chitosan and surface characteristics of cell wall. Acta Pharmacol. Sin. 2004, 25, 932–936. [Google Scholar]
- Joblin, K.N.; Naylor, G.E. Ruminal mycoplasma interactions with ruminal fungi and bacteria. S. Afr. J. Anim. Sci. 1999, 29, 115–116. [Google Scholar]
- Söllinger, A.; Tveit, A.T.; Poulsen, M.; Noel, S.J.; Bengtsson, M.; Bernhardt, J.; Frydendahl Hellwing, A.L.; Lund, P.; Riedel, K.; Schleper, C.; et al. Holistic assessment of rumen microbiome dynamics through quantitative metatranscriptomics reveals multifunctional redundancy during key steps of anaerobic feed degradation. mSystems 2018, 3, e00038-18. [Google Scholar] [CrossRef]
- Belanche, A.; Ramos-Morales, E.; Newbold, C.J. Screening of natural additives from crustaceans, diatoms, seaweeds and plant extracts to manipulate rumen fermentation. J. Sci. Food Agric. 2015, 96, 3069–3078. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.Y.; Liao, Y.T.; Tseng, Y.K.; Deng, F.S.; Lin, C.H. A potential antifungal effect of chitosan against Candida albicans is mediated via the inhibition of SAGA complex component expression and the subsequent alteration of cell surface integrity. Front. Microbiol. 2019, 10, 602. [Google Scholar] [CrossRef]
- Ing, L.Y.; Zin, N.M.; Sarwar, A.; Katas, H. Antifungal activity of chitosan nanoparticles and correlation with their physical properties. Int. J. Biomater. 2012, 2012, 632698. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular mechanisms of chitosan interactions with fungi and plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef]
- Marty, R.J.; Demeyer, D.I. The effect of inhibitors of methane production on fermentation pattern and stoichiometry in vitro using rumen contents from sheep given molasses. Br. J. Nutr. 1973, 30, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Ungerfeld, E.M. Metabolic hydrogen flows in rumen fermentation: Principles and possibilities of interventions. Front. Microbiol. 2020, 11, 589. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Ocampo, R.; Montoya-Flores, M.D.; Herrera-Torre, E.; Pámanes-Carrasco, G.; Arceo-Castillo, J.I.; Valencia-Salazar, S.E.; Arango, J.; Aguilar-Pérez, C.F.; Ramírez-Avilés, L.; Piñeiro-Vázquez, A.T.; et al. Effect of chitosan and naringin on enteric methane emissions in crossbred heifers fed tropical grass. Animals 2021, 11, 1599. [Google Scholar] [CrossRef]
- López-García, A.; Saborío-Montero, A.; Gutiérrez-Rivas, M.; Atxaerandio, R.; Goiri, I.; García-Rodríguez, A.; González-Recio, O. Fungal and ciliate protozoa are the main rumen microbes associated with methane emissions in dairy cattle. GigaScience 2020, 11, giab088. [Google Scholar] [CrossRef]
- Saborío-Montero, A.; Gutierrez-Rivas, M.; Goiri, I.; Atxaerandio, R.; García-Rodriguez, A.; López-Paredes, J.; González-Recio, O. Rumen eukaryotes are the main phenotypic risk factors for larger methane emissions in dairy cattle. Livest. Sci. 2020, 263, 105023. [Google Scholar] [CrossRef]
- Danielsson, R.; Dicksved, J.; Sun, L. Methane production in dairy cows correlates with rumen methanogenic and bacterial community structure. Front. Microbiol. 2017, 8, 226. [Google Scholar] [CrossRef]
- Gandra, J.R.; Takiya, C.S.; de Oliveira, E.R.; de Paiva, P.G.; de Tonissi, R.H.; de Goes, B.; Gandra, E.R.S.; Araki, H.M.C. Nutrient digestion, microbial protein synthesis, and blood metabolites of Jersey heifers fed chitosan andwhole raw soybeans. R. Bras. Zootec. 2016, 45, 130–137. [Google Scholar] [CrossRef]
- Zheng, Y.G.; Zhang, B.Q.; Qi, J.Y.; Zhao, Y.L.; Guo, X.Y.; Shi, B.L.; Yan, S.M. Dietary supplementation of chitosan affects milk performance, markers of inflammatory response and antioxidant estatus in dairy cows. Anim. Feed Sci. Technol. 2021, 277, 114952. [Google Scholar] [CrossRef]
- Mima, S.; Miya, M.; Iwamoto, R.; Yoshikawa, S. Highly deacetylated chitosan and its properties. J. Appl. Polym. Sci. 1983, 28, 1909–1917. [Google Scholar] [CrossRef]
- Mingoti, R.D.; Freitas, J.E.; Gandra, J.R.; Gardinal, R.; Calomeni, G.D.; Barletta, R.V.; Vendramini, T.H.A.; Paiva, P.G.; Renno, F.P. Dose response of chitosan on nutrient digestibility, blood metabolites and lactation performance in Holstein dairy cows. Livest. Sci. 2016, 187, 35–39. [Google Scholar] [CrossRef]
- Pereira, T.L.; Fernandes, A.R.M.; Oliveira, E.R.; Cônsolo, N.R.B.; Marques, O.F.C.; Maciel, T.P.; Pordeus, N.M.; Barbosa, L.C.G.S.; Buarque, V.L.M.; Padilla, A.R.H.; et al. Serum metabolomic fingerprints of lambs fed chitosan and its association with performance and meat quality traits. Animal 2020, 14, 1987–1998. [Google Scholar] [CrossRef]
- Wan, J.; Jiang, F.; Xu, Q.; Chen, D.; Yu, B.; Huang, Z.; He, J. New insights into the role of chitosan oligosaccharide in enhancing growth performance, antioxidant capacity, immunity and intestinal development of weaned pigs. RSC Adv. 2017, 7, 9669–9679. [Google Scholar] [CrossRef]
- Cohick, W.S. Role of the insulin-like growth factors and their binding proteins in lactation. J. Dairy Sci. 1998, 81, 1769–1777. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| HF | BS | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| CTR | CHI | CTR | CHI | SEM | Breed | Treat | Breed × Treat | |
| Total VFA, mM | 62.9 | 66.6 | 69.8 | 68.4 | 5.04 | 0.418 | 0.823 | 0.628 |
| Individual VFA, mol/100 mol | ||||||||
| Acetic | 65.0 | 62.5 | 64.3 | 64.1 | 0.60 | 0.444 | 0.046 | 0.109 |
| Propionic | 16.7 | 18.8 | 16.6 | 18.2 | 0.57 | 0.533 | 0.008 | 0.674 |
| Butyric | 14.2 | 14.3 | 15.2 | 14.1 | 0.52 | 0.493 | 0.376 | 0.288 |
| Isobutyric | 0.956 | 0.832 | 0.872 | 0.784 | 0.0775 | 0.430 | 0.210 | 0.827 |
| Valeric | 1.28 | 1.25 | 1.29 | 1.26 | 0.054 | 0.877 | 0.635 | 0.934 |
| Isovaleric | 1.87 | 2.25 | 1.69 | 1.56 | 0.161 | 0.022 | 0.477 | 0.154 |
| BCVFA | 2.83 | 3.08 | 2.56 | 2.34 | 0.220 | 0.046 | 0.945 | 0.321 |
| C2/C3 | 3.93 | 3.34 | 3.92 | 3.54 | 0.159 | 0.569 | 0.013 | 0.551 |
| C2 + C4/C3 | 4.84 | 4.15 | 4.89 | 4.36 | 0.195 | 0.522 | 0.011 | 0.707 |
| HF | BS | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| CTR | CHI | CTR | CHI | SEM | Breed | Treat | Breed × Treat | |
| Yield, kg/d | ||||||||
| Milk | 25.8 | 26.2 | 26.0 | 27.3 | 1.52 | 0.718 | 0.562 | 0.756 |
| ECM | 26.7 | 24.0 | 28.3 | 29.3 | 2.25 | 0.160 | 0.709 | 0.406 |
| Fat | 1.22 | 0.757 | 1.13 | 1.16 | 0.1536 | 0.355 | 0.167 | 0.131 |
| Protein | 1.05 | 0.912 | 0.944 | 0.993 | 0.1048 | 0.921 | 0.664 | 0.365 |
| Lactose | 1.49 | 1.32 | 1.29 | 1.34 | 0.1233 | 0.539 | 0.627 | 0.358 |
| Composition, % | ||||||||
| Fat | 4.68 | 3.61 | 4.09 | 4.00 | 0.306 | 0.742 | 0.080 | 0.130 |
| Protein | 3.54 | 3.54 | 3.65 | 3.62 | 0.094 | 0.882 | 0.358 | 0.883 |
| Lactose | 4.83 | 4.73 | 4.73 | 4.90 | 0.069 | 0.588 | 0.640 | 0.056 |
| CH4 | ||||||||
| ppm | 1605 | 1770 | 2325 | 1924 | 329.2 | 0.271 | 0.727 | 0.461 |
| g/d | 214 | 206 | 290 | 234 | 45.3 | 0.337 | 0.498 | 0.642 |
| g/kg milk | 9.54 | 9.14 | 10.85 | 9.51 | 2.242 | 0.740 | 0.694 | 0.496 |
| HF | BS | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| CTR | CHI | CTR | CHI | SEM | Breed | Treat | Breed × Treat | |
| Blood parameters | ||||||||
| BUN, mmol/L | 2.52 | 2.39 | 2.71 | 2.81 | 0.249 | 0.267 | 0.960 | 0.672 |
| Glucose, mmol/L | 3.70 | 4.49 | 3.63 | 3.28 | 0.161 | 0.002 | 0.216 | 0.005 |
| IGF-1, ng/mL | 141 | 188 | 103 | 99 | 12.0 | <0.001 | 0.115 | 0.068 |
| N flux | ||||||||
| Purine derivative excretion (mmol/d) | 332 | 279 | 300 | 312 | 28.1 | 0.981 | 0.496 | 0.291 |
| N flux (g N/d) | 244 | 204 | 236 | 231 | 26.4 | 0.735 | 0.432 | 0.535 |
| HF | BS | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| CTR | CHI | CTR | CHI | SEM | Breed | Treat | Breed × Treat | |
| Observed | 681 | 636 | 652 | 699 | 63.6 | 0.743 | 0.984 | 0.397 |
| Chao1 | 773 | 730 | 757 | 781 | 28.9 | 0.460 | 0.699 | 0.181 |
| Shannon | 3.01 | 2.59 | 2.59 | 2.75 | 0.180 | 0.405 | 0.387 | 0.077 |
| InvSimpson | 6.85 | 4.91 | 5.16 | 5.77 | 0.956 | 0.606 | 0.411 | 0.129 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rey, J.; Díaz de Otálora, X.; Atxaerandio, R.; Mandaluniz, N.; García-Rodríguez, A.; González-Recio, O.; López-García, A.; Ruiz, R.; Goiri, I. Effect of Chitosan on Ruminal Fermentation and Microbial Communities, Methane Emissions, and Productive Performance of Dairy Cattle. Animals 2023, 13, 2861. https://doi.org/10.3390/ani13182861
Rey J, Díaz de Otálora X, Atxaerandio R, Mandaluniz N, García-Rodríguez A, González-Recio O, López-García A, Ruiz R, Goiri I. Effect of Chitosan on Ruminal Fermentation and Microbial Communities, Methane Emissions, and Productive Performance of Dairy Cattle. Animals. 2023; 13(18):2861. https://doi.org/10.3390/ani13182861
Chicago/Turabian StyleRey, Jagoba, Xabier Díaz de Otálora, Raquel Atxaerandio, Nerea Mandaluniz, Aser García-Rodríguez, Oscar González-Recio, Adrián López-García, Roberto Ruiz, and Idoia Goiri. 2023. "Effect of Chitosan on Ruminal Fermentation and Microbial Communities, Methane Emissions, and Productive Performance of Dairy Cattle" Animals 13, no. 18: 2861. https://doi.org/10.3390/ani13182861