
In Vivo Efficacy of Purified Quillaja Saponin Extracts in Protecting against Piscirickettsia salmonis Infections in Atlantic Salmon (Salmo salar)
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Quillaja Saponin Extracts
2.2. In Vivo Efficacy of Quillaja Saponin Extracts against P. salmonis in Freshwater Conditions under Intraperitoneal Challenge
2.3. In Vivo Efficacy of Quillaja Saponin Extracts against P. salmonis in Atlantic Salmon Smolts, under a Cohabitation Challenge
2.4. RNA Extraction and Quantitative Analyses of Immune Gene Expression by RT-qPCR
2.5. Efficacy of PAQ-Xtract in the Prevention of P. salmonis Infections in Atlantic Salmon in Seawater under Pilot Commercial Conditions
2.6. Statistical Analyses
3. Results
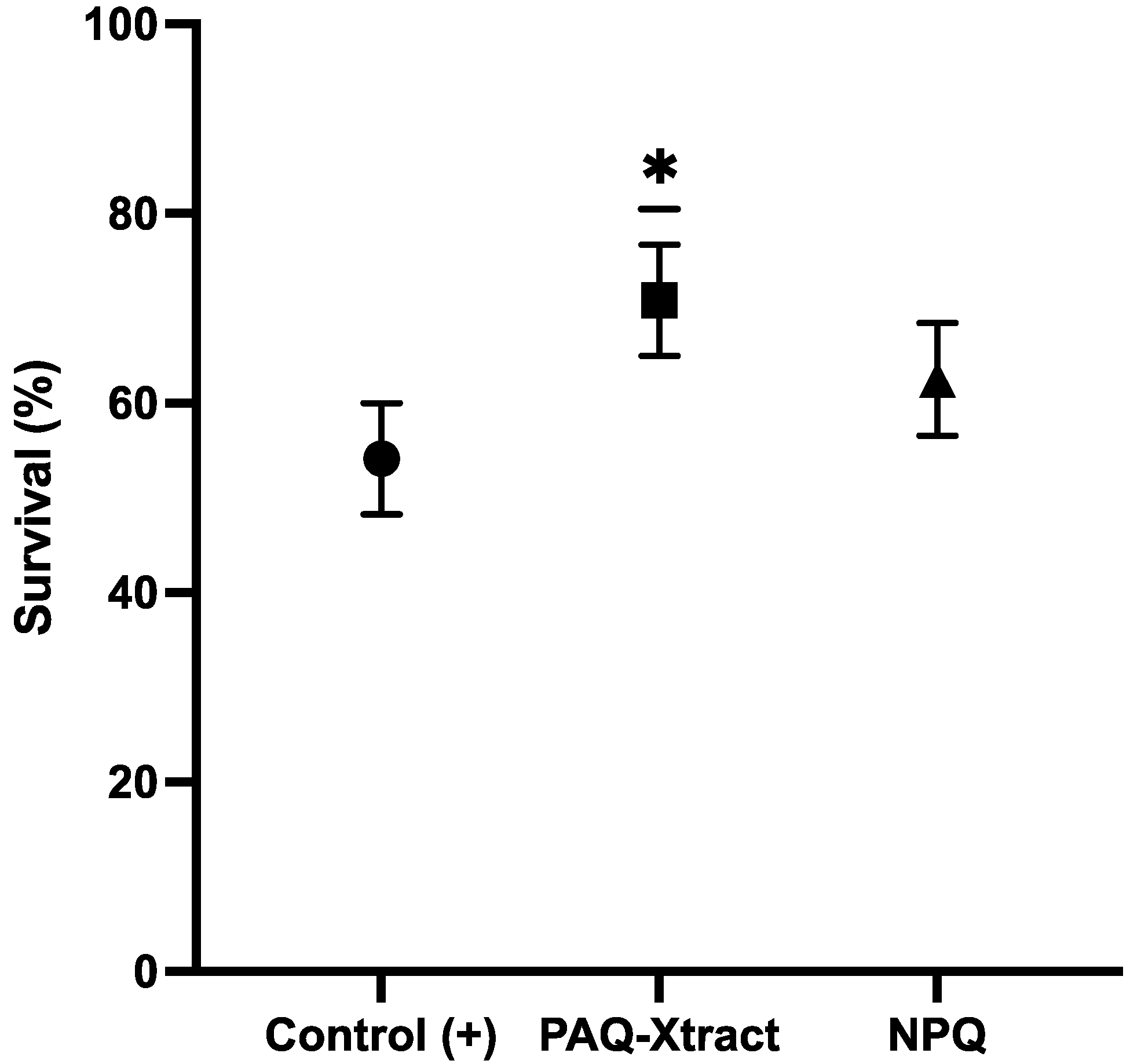
3.1. In Vivo Efficacy of Quillaja Saponin Extracts against P. salmonis in Freshwater Conditions under Intraperitoneal Challenge
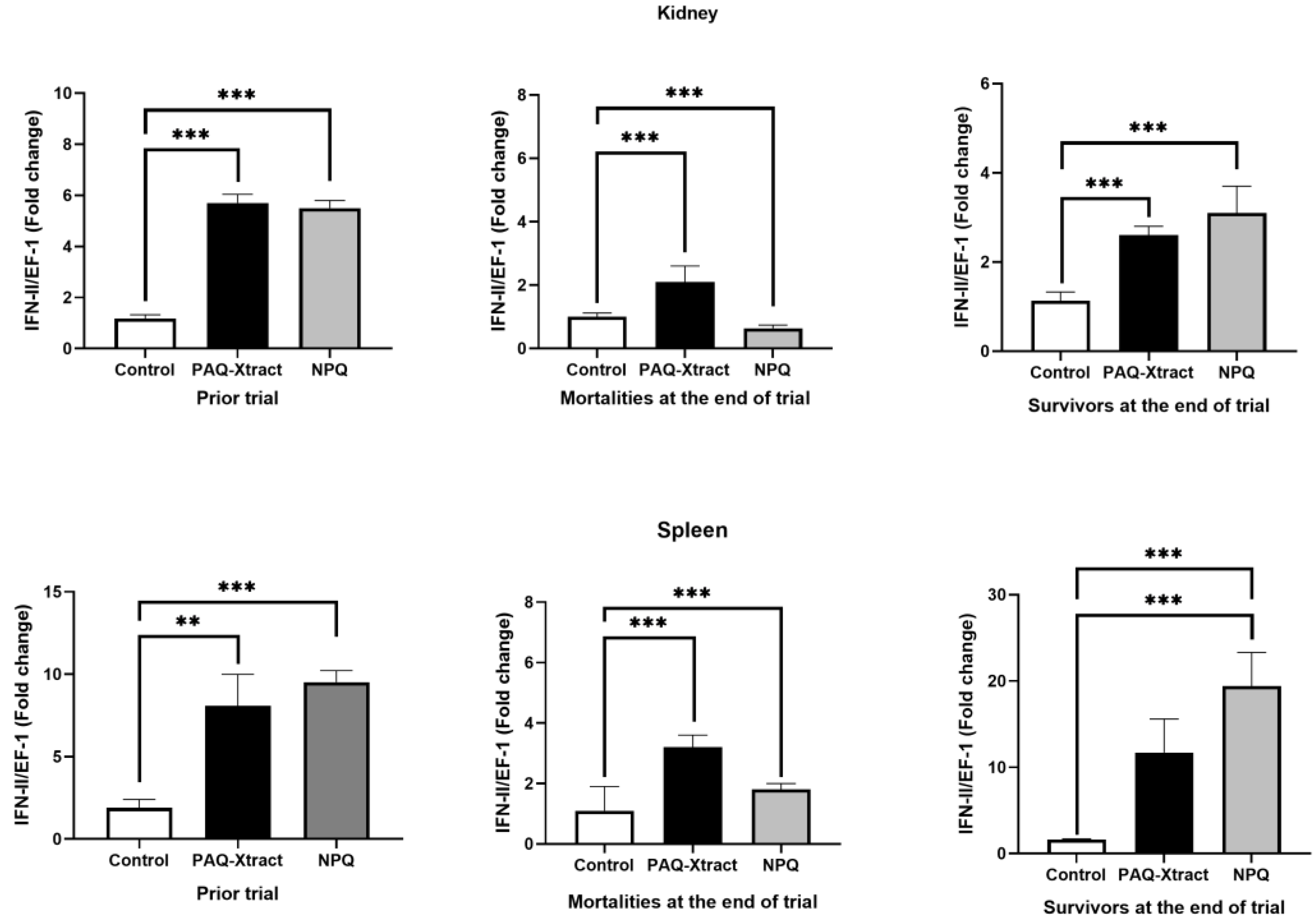
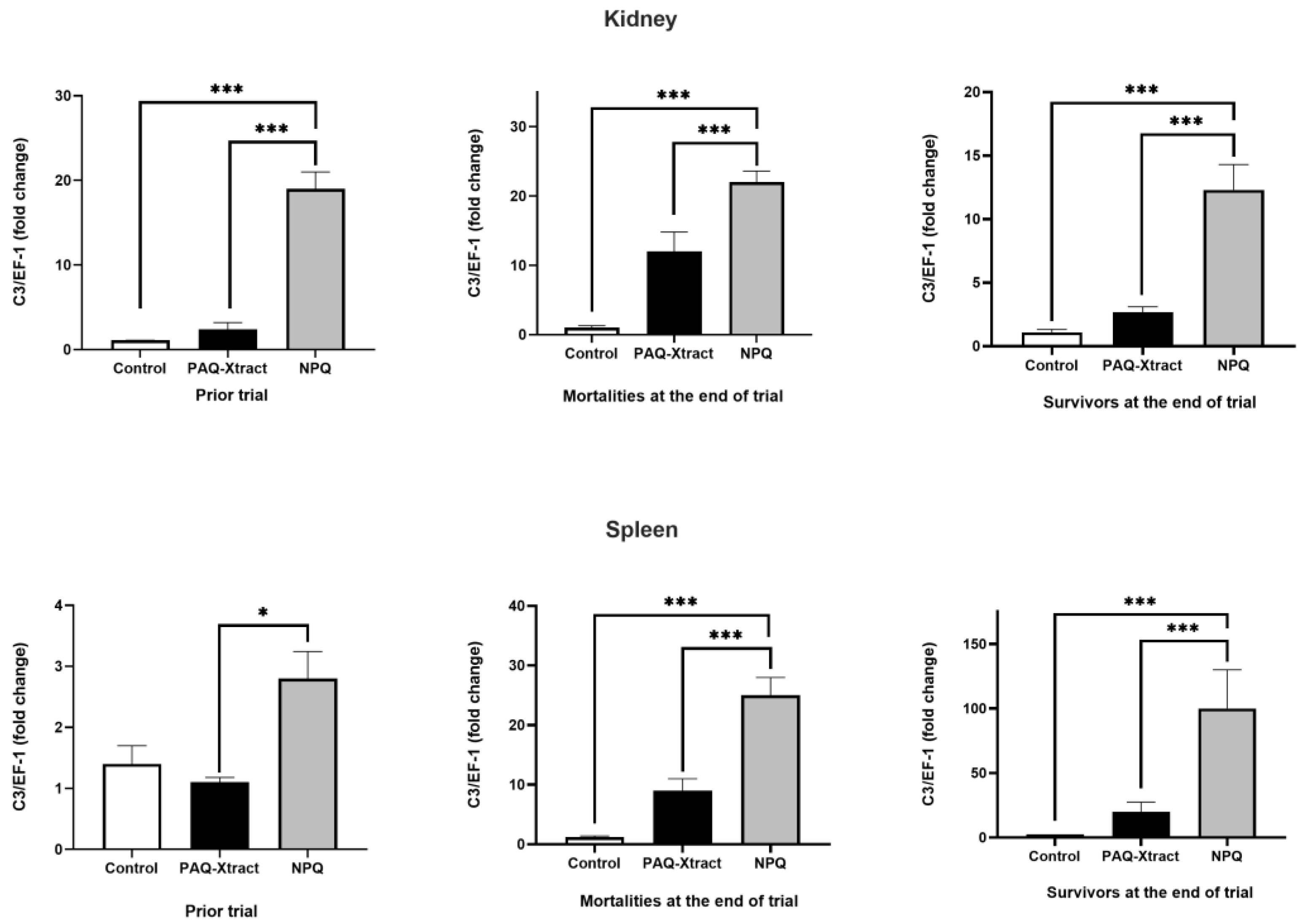
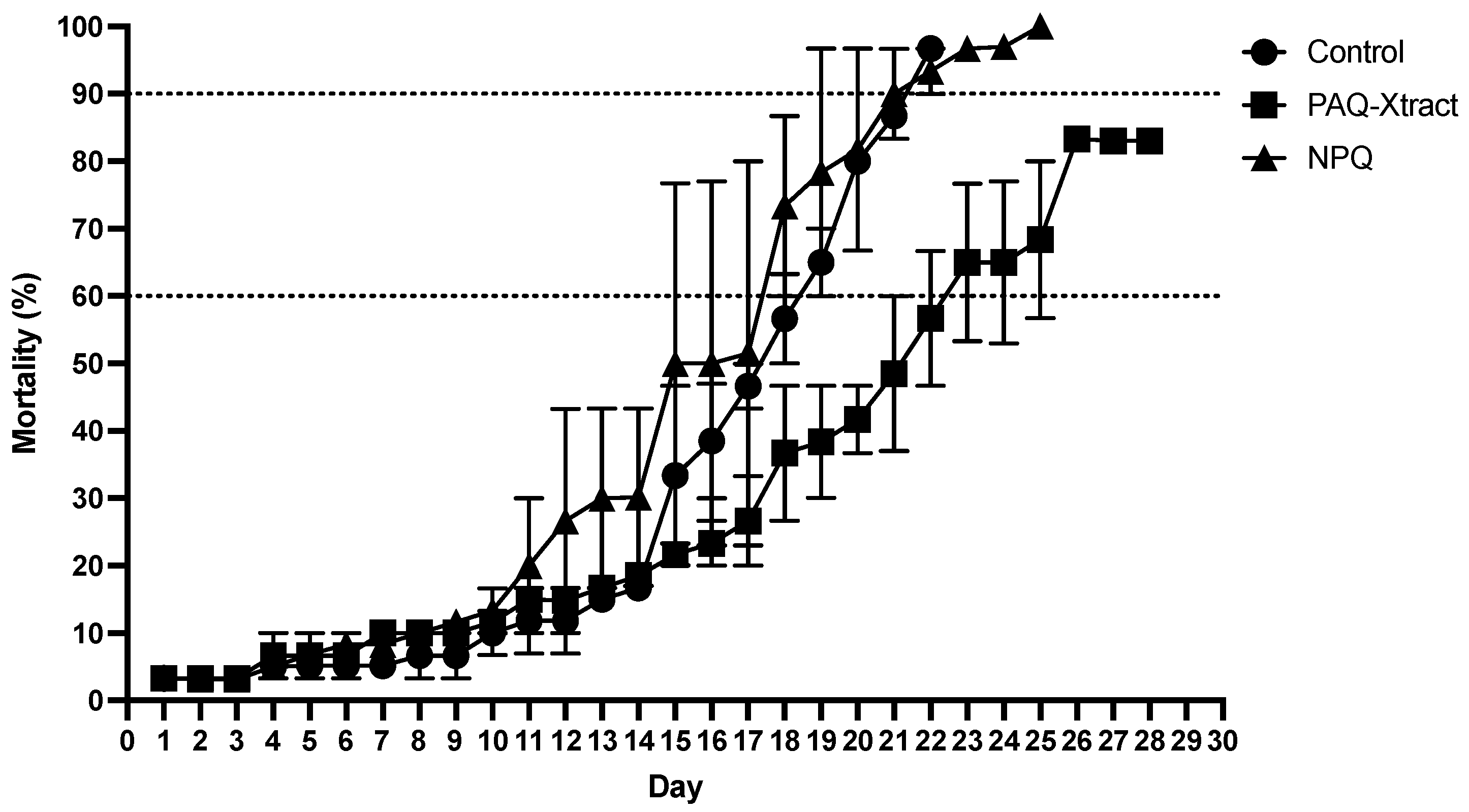
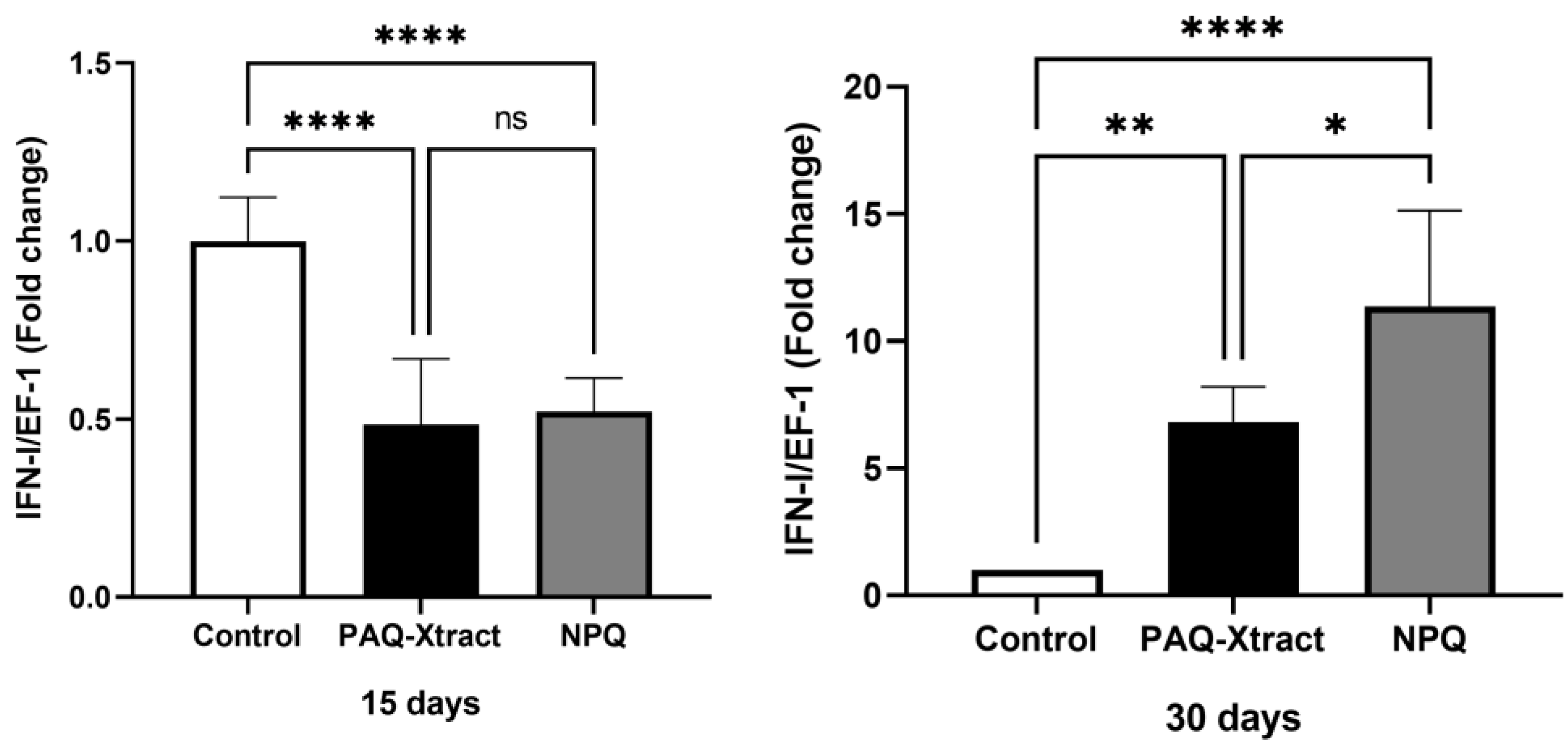
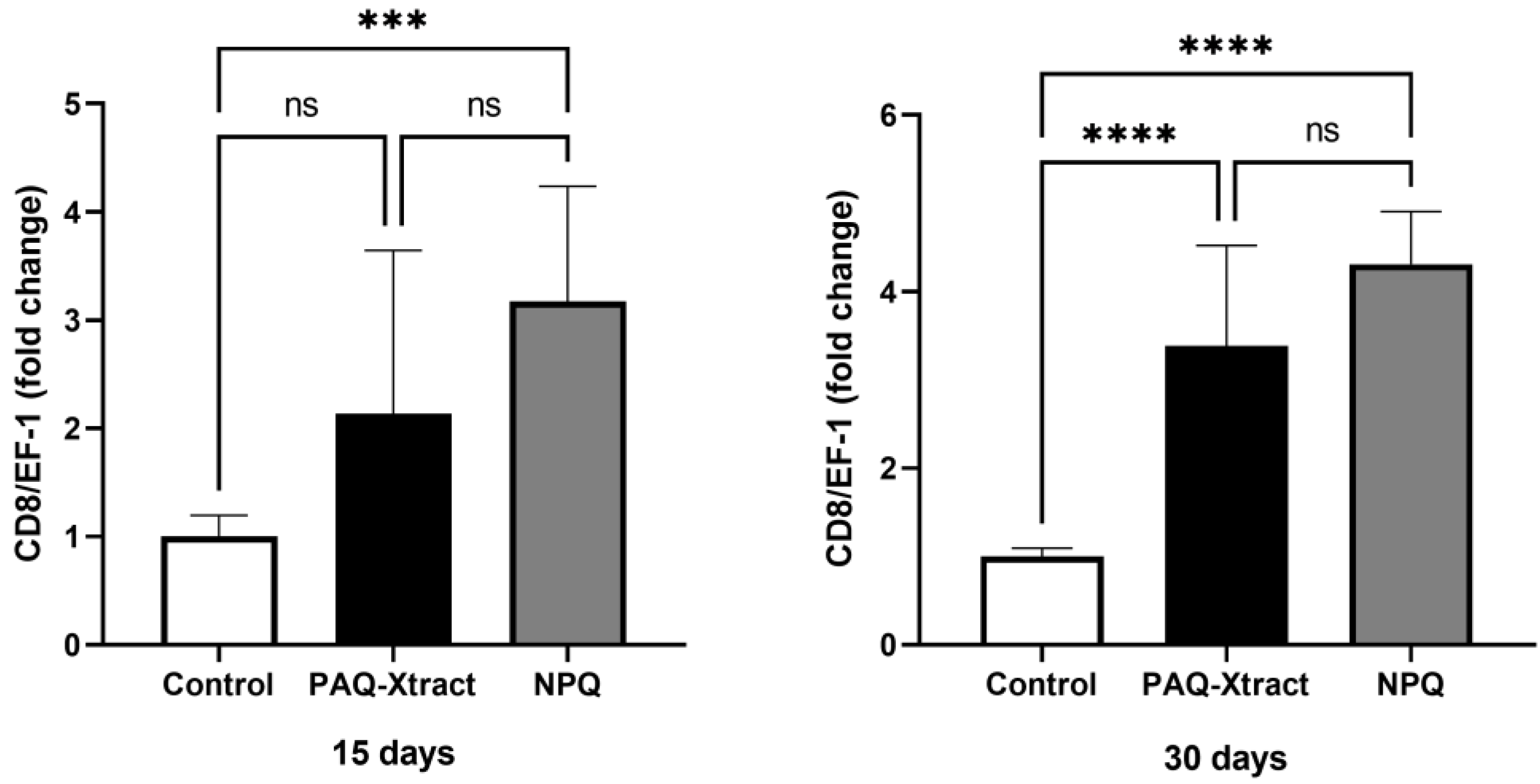
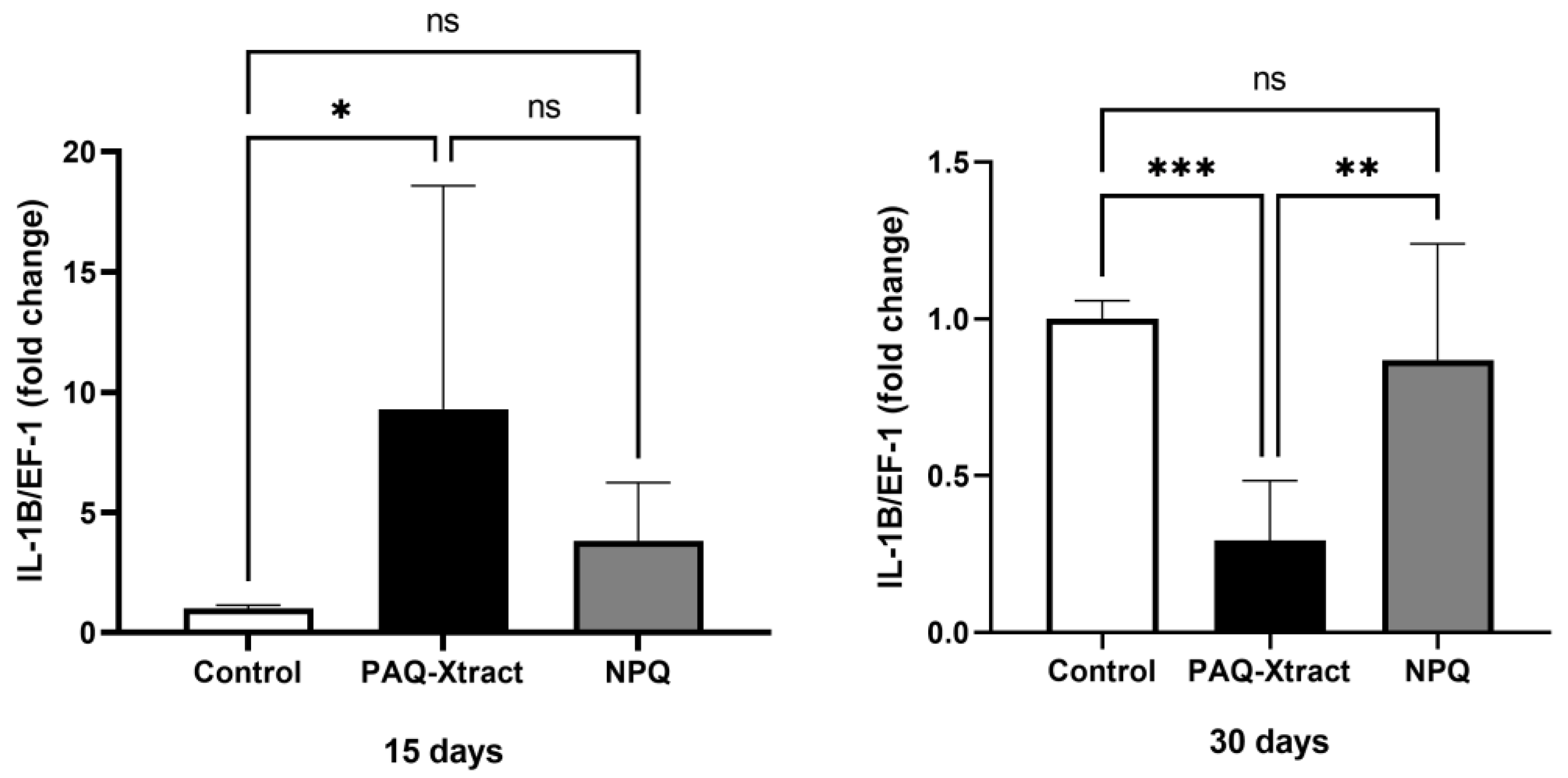
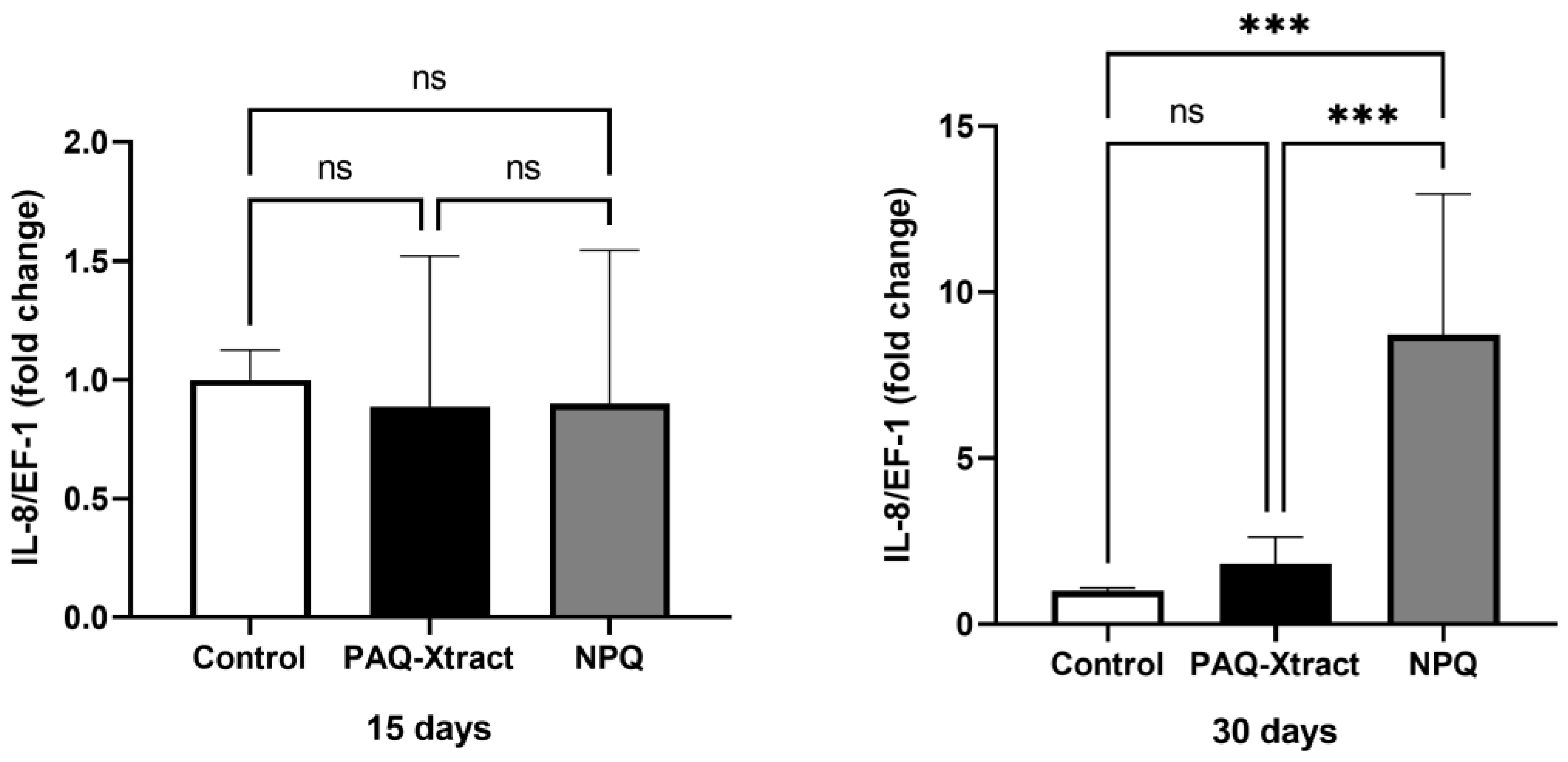
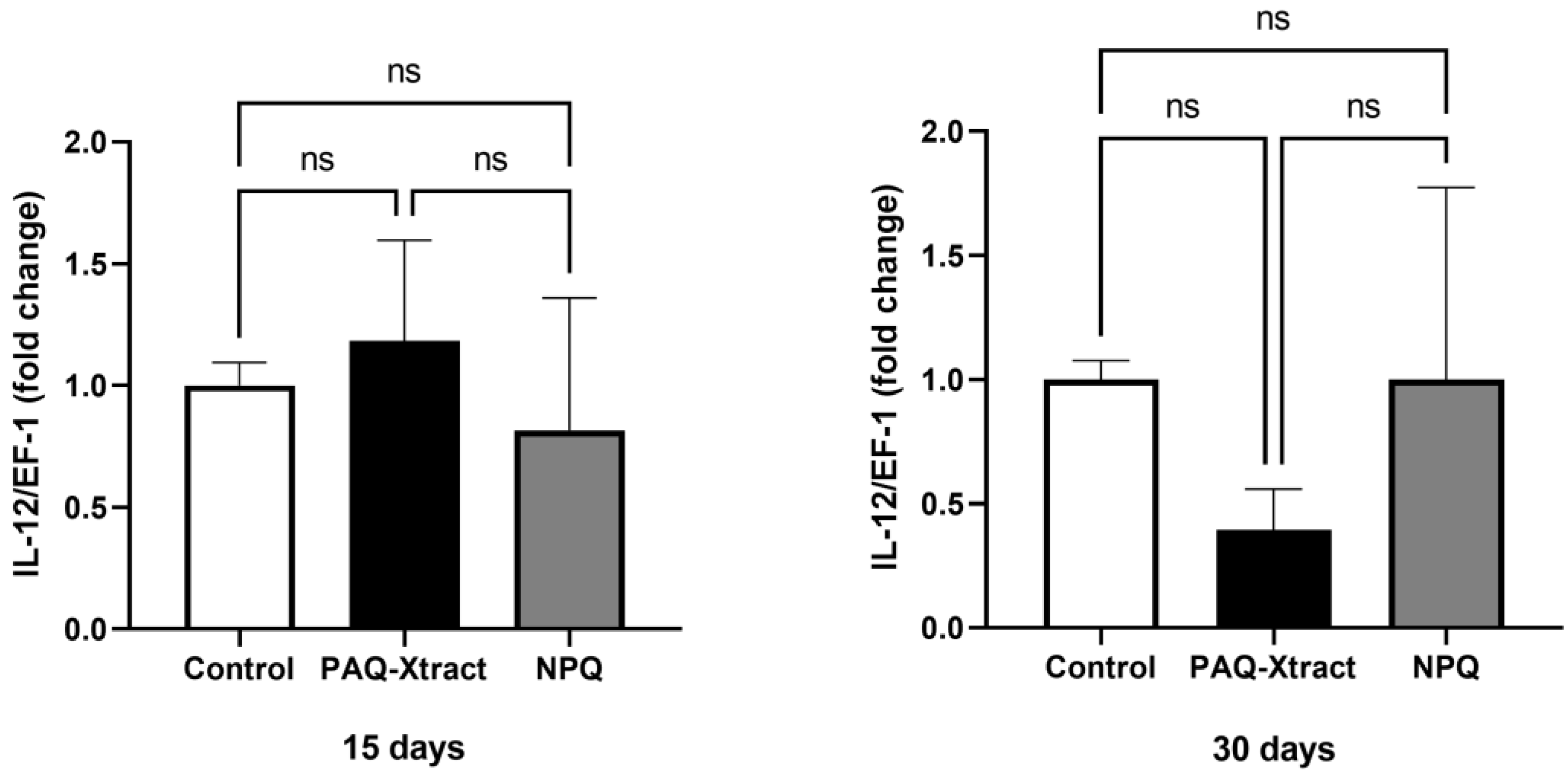
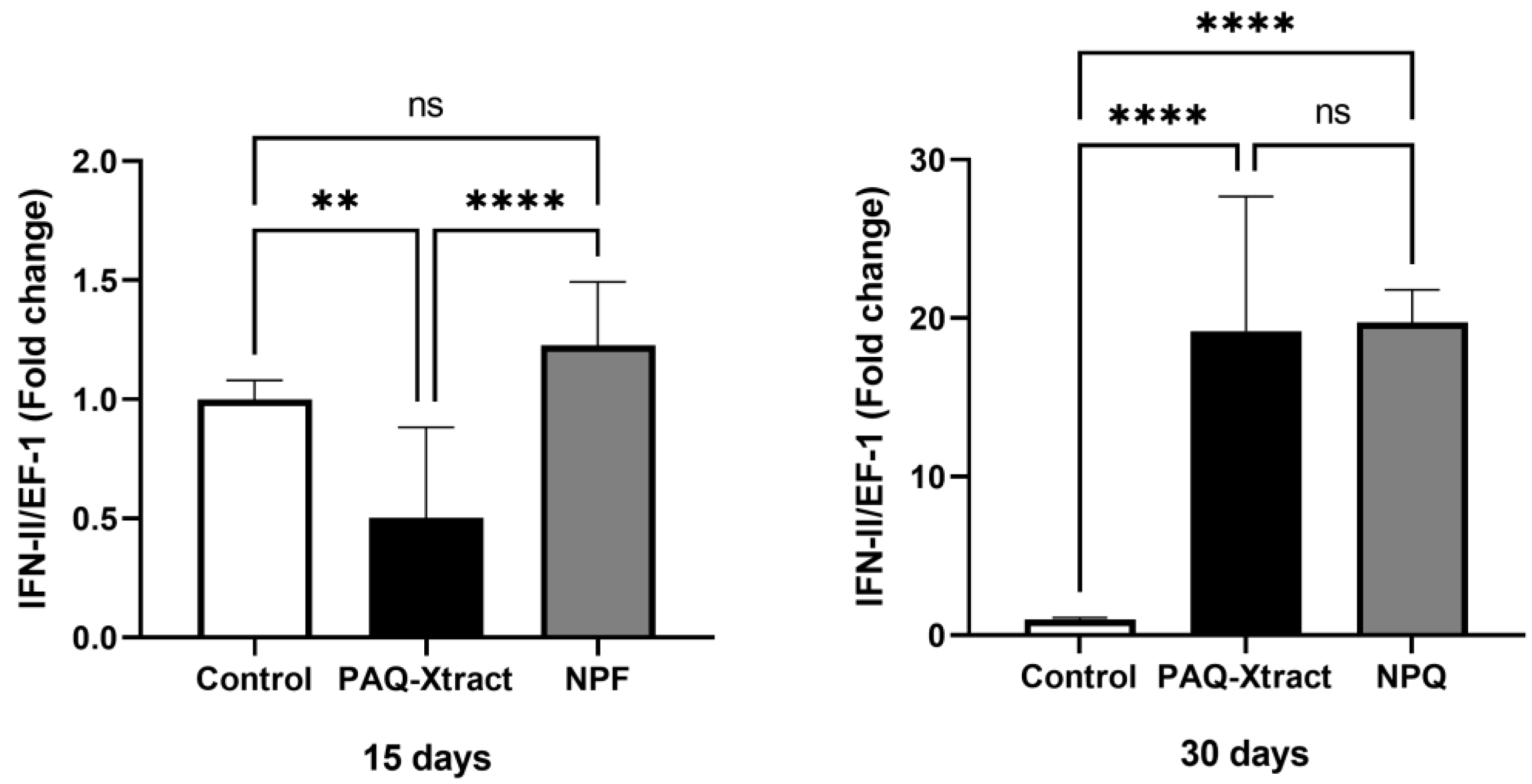
3.2. In Vivo Efficacy of Quillaja Extracts against P. salmonis in Atlantic Salmon Smolts, under a Cohabitation Challenge
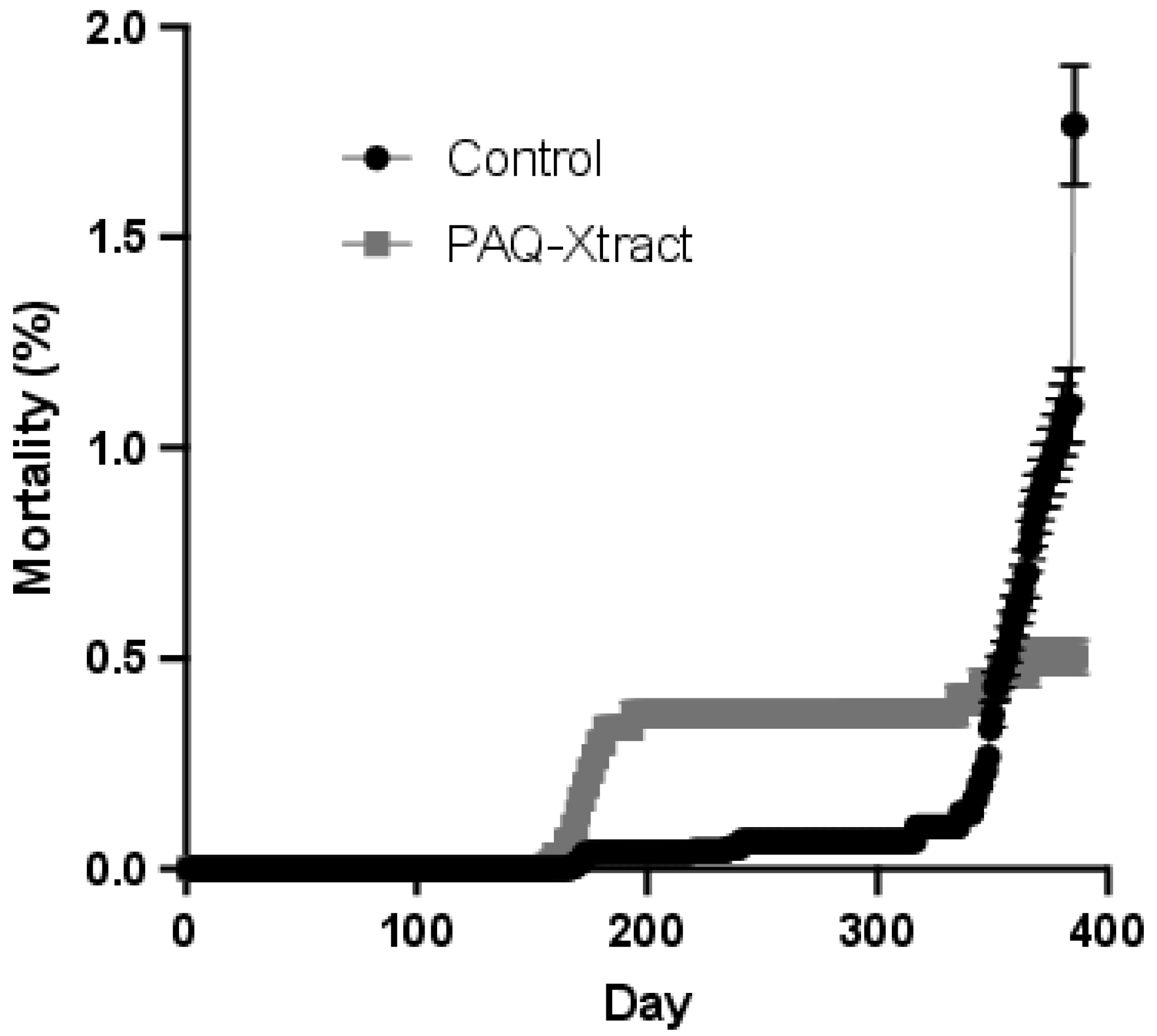
3.3. Efficacy of PAQ-Xtract in the Prevention of P. salmonis Infections in Atlantic Salmon in Seawater under Pilot Commercial Conditions
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rozas, M.; Enríquez, R. Piscirickettsiosis and Piscirickettsia Salmonis in Fish: A Review. J. Fish Dis. 2014, 37, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Fryer, J.L.; Hedrick, R.P. Piscirickettsia Salmonis: A Gram-Negative Intracellular Bacterial Pathogen of Fish. J. Fish Dis. 2003, 26, 251–262. [Google Scholar] [CrossRef]
- Departamento de Salud Animal. Informe Sanitario Con Información Sanitaria de Agua Dulce Y Mar 1° Semestre Año 2022 Departamento de Salud Animal Subdirección de Acuicultura Servicio Nacional de Pesca Y Acuicultura; Departamento de Salud Animal: Madrid, Spain, 2022. [Google Scholar]
- Watts, J.E.M.; Schreier, H.J.; Lanska, L.; Hale, M.S. The Rising Tide of Antimicrobial Resistance in Aquaculture: Sources, Sinks and Solutions. Mar. Drugs 2017, 15, 158. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.D.; Godoy, F.A.; Lee, M.R. Current Status of the Use of Antibiotics and the Antimicrobial Resistance in the Chilean Salmon Farms. Fron. Microbiol. 2018, 9, 1284. [Google Scholar] [CrossRef]
- Avendano-Herrera, R.; Mancilla, M.; Miranda, C.D. Use of Antimicrobials in Chilean Salmon Farming: Facts, Myths and Perspectives. Rev. Aquac. 2022, 15, 89–111. [Google Scholar] [CrossRef]
- Rozas-Serri, M. Why Does Piscirickettsia Salmonis Break the Immunological Paradigm in Farmed Salmon? Biological Context to Understand the Relative Control of Piscirickettsiosis. Front. Immunol. 2022, 13, 856896. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C. Heavy Use of Prophylactic Antibiotics in Aquaculture: A Growing Problem for Human and Animal Health and for the Environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Pulgar, R.; Hödar, C.; Travisany, D.; Zuñiga, A.; Domínguez, C.; Maass, A.; González, M.; Cambiazo, V. Transcriptional Response of Atlantic Salmon Families to Piscirickettsia Salmonis Infection Highlights the Relevance of the Iron-Deprivation Defence System. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.A.; Gomez, F.A.; Mercado, L.; Ramírez, R.; Marshall, S.H. Piscirickettsia Salmonis Imbalances the Innate Immune Response to Succeed in a Productive Infection in a Salmonid Cell Line Model. PLoS ONE 2016, 11, e0163943. [Google Scholar] [CrossRef]
- Uribe, C.; Folch, H.; Enriquez, R.; Moran, G. Innate and Adaptive Immunity in Teleost Fish: A Review. Vet. Med. 2011, 56, 486–503. [Google Scholar] [CrossRef]
- Zhao, M.; Liu, Y.; Gao, Y.; Wang, X.; Zhou, H.; Zhang, A. Insights into the Functional Role of Grass Carp IL-8 in Head Kidney Leukocytes: Pro-inflammatory Effects and Signalling Mechanisms. J. Fish Biol. 2022, 100, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Martínez, D.E.; Santana, P.A.; Aróstica, M.; Forero, J.C.; Guzmán, F.; Mercado, L. Immunodetection of Rainbow Trout IL-8 Cleaved-Peptide: Tissue Bioavailability and Potential Antibacterial Activity in a Bacterial Infection Context. Dev. Comp. Immunol. 2021, 124, 104182. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Bruce, T.J.; Jones, E.M.; Cain, K.D. A Review of Fish Vaccine Development Strategies: Conventional Methods and Modern Biotechnological Approaches. Microorganisms 2019, 7, 569. [Google Scholar] [CrossRef] [PubMed]
- Mezouar, S.; Mege, J.L. Changing the Paradigm of IFN-γ at the Interface between Innate and Adaptive Immunity: Macrophage-Derived IFN-γ. J. Leukoc. Biol. 2020, 108, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Mikkelsen, H.; Jungersen, G. Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J. Immunol. Res. 2019, 2019, 1356540. [Google Scholar] [CrossRef] [PubMed]
- Løvoll, M.; Johnsen, H.; Boshra, H.; Bøgwald, J.; Sunyer, J.O.; Dalmo, R.A. The Ontogeny and Extrahepatic Expression of Complement Factor C3 in Atlantic Salmon (Salmo salar). Fish Shellfish. Immunol. 2007, 23, 542–552. [Google Scholar] [CrossRef]
- San Martín, R.; Briones, R. Quality Control of Commercial Quillaja (Quillaja saponaria Molina) Extracts by Reverse Phase HPLC. J. Sci. Food Agric. 2000, 80, 2063–2068. [Google Scholar] [CrossRef]
- Maier, C.; Conrad, J.; Carle, R.; Weiss, J.; Schweiggert, R.M. Phenolic Constituents in Commercial Aqueous Quillaja (Quillaja saponaria Molina) Wood Extracts. J. Agric. Food Chem. 2015, 63, 1756–1762. [Google Scholar] [CrossRef]
- Cortés, H.D.; Gómez, F.A.; Marshall, S.H. The Phagosome–Lysosome Fusion Is the Target of a Purified Quillaja saponin Extract (Pqse) in Reducing Infection of Fish Macrophages by the Bacterial Pathogen Piscirickettsia salmonis. Antibiotics 2021, 10, 847. [Google Scholar] [CrossRef]
- Guerra, F.; Sepúlveda, S. Saponin Production from Quillaja Genus Species. An Insight into Its Applications and Biology. Sci. Agric. 2020, 78, 1–9. [Google Scholar] [CrossRef]
- Fleck, J.D.; Kauffmann, C.; Spilki, F.; Lencina, C.L.; Roehe, P.M.; Gosmann, G. Adjuvant Activity of Quillaja Brasiliensis Saponins on the Immune Responses to Bovine Herpesvirus Type 1 in Mice. Vaccine 2006, 24, 7129–7134. [Google Scholar] [CrossRef]
- Cañon-Jones, H.; Cortes, H.; Castillo-Ruiz, M.; Schlotterbeck, T.; Martín, R.S. Quillaja Saponaria (Molina) Extracts Inhibits in Vitro Piscirickettsia Salmonis Infections. Animals 2020, 10, 2286. [Google Scholar] [CrossRef]
- Peña, A.A.; Bols, N.C.; Marshall, S.H. An Evaluation of Potential Reference Genes for Stability of Expression in Two Salmonid Cell Lines after Infection with Either Piscirickettsia Salmonis or IPNV. BMC Res. Notes 2010, 3, 101. [Google Scholar] [CrossRef]
- Fernandes, R.D.N. [UNESP] Uso de Saponina de Quilaia (Quillaja saponaria Molina) Em Juvenis de Pacu. Aleph 2014, viii, 103. [Google Scholar] [CrossRef]
- Sannasimuthu, A.; Brindha, P.; Chandrasekar, S.; Mariappan, P.; Antony, A.; Velanganni, A.J.; Harikrishnan, R. Effects of Saponin Supplementation Diet on Immune Response in Cyprinus Carpio against Aeromonas Hydrophila. In Proceedings of the 7th National Symposium on Advance Research in Biosciences, Thanjavur, India, 3–4 March 2014. [Google Scholar]
- Welsby, I.; Detienne, S.; N’Kuli, F.; Thomas, S.; Wouters, S.; Bechtold, V.; De Wit, D.; Gineste, R.; Reinheckel, T.; Elouahabi, A.; et al. Lysosome-Dependent Activation of Human Dendritic Cells by the Vaccine Adjuvant QS-21. Front. Immunol. 2017, 7, 663. [Google Scholar] [CrossRef]
- Katayama, S.; Oda, K.; Ohgitani, T.; Hirahara, T.; Shimizu, Y. Influence of Antigenic Forms and Adjuvants on the IgG Subclass Antibody Response to Aujeszky’s Disease Virus in Mice. Vaccine 1999, 17, 2733–2739. [Google Scholar] [CrossRef]
- Rojas, V.; Galanti, N.; Bols, N.C.; Marshall, S.H. Productive Infection of Piscirickettsia Salmonis in Macrophages and Monocyte-like Cells from Rainbow Trout, a Possible Survival Strategy. J. Cell. Biochem. 2009, 108, 631–637. [Google Scholar] [CrossRef]
- Martínez, D.P.; Oliver, C.; Santibañez, N.; Coronado, J.L.; Oyarzún-Salazar, R.; Enriquez, R.; Vargas-Chacoff, L.; Romero, A. PAMPs of Piscirickettsia Salmonis Trigger the Transcription of Genes Involved in Nutritional Immunity in a Salmon Macrophage-Like Cell Line. Front. Immunol. 2022, 13, 1667. [Google Scholar] [CrossRef]
- Lee, H.-J.; Woo, Y.; Hahn, T.-W.; Jung, Y.M.; Jung, Y.-J. Formation and Maturation of the Phagosome: A Key Mechanism in Innate Immunity against Intracellular Bacterial Infection. Microorganisms 2020, 8, 1298. [Google Scholar] [CrossRef]
- Hirayama, D.; Iida, T.; Nakase, H. The Phagocytic Function of Macrophage-Enforcing Innate Immunity and Tissue Homeostasis. Int. J. Mol. Sci. 2018, 19, 92. [Google Scholar] [CrossRef]
- Ninomiya, M.; Hatta, H.; Fujiki, M.; Kim, M.; Yamamoto, T.; Kusuda, R. Enhancement of Chemotactic Activity of Yellowtail (Seriola quinqueradiata) Leucocytes by Oral Administration of Quillaja Saponin. Fish Shellfish. Immunol. 1995, 5, 325–328. [Google Scholar] [CrossRef]
- Sparg, S.G.; Light, M.E.; Van Staden, J. Biological Activities and Distribution of Plant Saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef] [PubMed]
- Fleck, J.D.; Betti, A.H.; Pereira da Silva, F.; Troian, E.A.; Olivaro, C.; Ferreira, F.; Verza, S.G. Saponins from Quillaja Saponaria and Quillaja Brasiliensis: Particular Chemical Characteristics and Biological Activities. Molecules 2019, 24, 171. [Google Scholar] [CrossRef] [PubMed]
- Rajput, Z.I.; Hu, S.H.; Xiao, C.W.; Arijo, A.G. Review: Adjuvant Effects of Saponins on Animal Immune Responses. J. Zhejiang Univ. Sci. B 2007, 8, 153. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The Biological Action of Saponins in Animal Systems: A Review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Ng’ambi, J.W.; Li, R.; Mu, C.; Song, W.; Liu, L.; Wang, C. Dietary Administration of Saponin Stimulates Growth of the Swimming Crab Portunus Trituberculatus and Enhances Its Resistance against Vibrio Alginolyticus Infection. Fish Shellfish. Immunol. 2016, 59, 305–311. [Google Scholar] [CrossRef]
- Cicchese, J.M.; Evans, S.; Hult, C.; Joslyn, L.R.; Wessler, T.; Millar, J.A.; Marino, S.; Cilfone, N.A.; Mattila, J.T.; Linderman, J.J.; et al. Dynamic Balance of Pro- and Anti-Inflammatory Signals Controls Disease and Limits Pathology. Immunol. Rev. 2018, 285, 147–167. [Google Scholar] [CrossRef]
- Morales-Lange, B.; Nombela, I.; Ortega-Villaizán, M.D.M.; Imarai, M.; Schmitt, P.; Mercado, L. Induction of Foxp3 during the Crosstalk between Antigen Presenting Like-Cells MHCII+CD83+ and Splenocytes CD4+IgM− in Rainbow Trout. Biology 2021, 10, 324. [Google Scholar] [CrossRef]
- Zhou, W.; Krogdahl, Å.; Sæle, Ø.; Chikwati, E.; Løkka, G.; Kortner, T.M. Digestive and Immune Functions in the Intestine of Wild Ballan Wrasse (Labrus bergylta). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2021, 260, 111011. [Google Scholar] [CrossRef]
- Bureau, D.P.; Harris, A.M.; Young Cho, C. The Effects of Purified Alcohol Extracts from Soy Products on Feed Intake and Growth of Chinook Salmon (Oncorhynchus tshawytscha) and Rainbow Trout (Oncorhynchus mykiss). Aquaculture 1998, 161, 27–43. [Google Scholar] [CrossRef]
- Mohammad, R.D. Effect of Different Levels of Saponin on Growth Performance and Food Efficiency in Convict Cichlid (Amatitlania nigrofasciata). J. Fish Aquat. Sci. 2019, 15, 22–26. [Google Scholar] [CrossRef]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. Reflections on ‘The Biological Action of Saponins in Animal Systems: A Review’. Br. J. Nutr. 2022, 127, 1034–1036. [Google Scholar] [CrossRef] [PubMed]
- Henríquez, P.; Kaiser, M.; Bohle, H.; Bustos, P.; Mancilla, M. Comprehensive Antibiotic Susceptibility Profiling of Chilean Piscirickettsia salmonis Field Isolates. J. Fish Dis. 2016, 39, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Bondad-Reantaso, M.G.; MacKinnon, B.; Karunasagar, I.; Fridman, S.; Alday-Sanz, V.; Brun, E.; Le Groumellec, M.; Li, A.; Surachetpong, W.; Karunasagar, I.; et al. Review of Alternatives to Antibiotic Use in Aquaculture. Rev. Aquac. 2023, 1–31. [Google Scholar] [CrossRef]
- Rahimi, N.N.M.N.; Ikhsan, N.F.M.; Loh, J.Y.; Ranzil, F.K.E.; Gina, M.; Lim, S.H.E.; Lai, K.S.; Chong, C.M. Phytocompounds as an Alternative Antimicrobial Approach in Aquaculture. Antibiotics 2022, 11, 469. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences (5′-3′) | Reference/ Accession Number GenBank | |
|---|---|---|---|
| IFN-I | F R | CCTGCCATGAAACCTGAGAAGA TTTCCTGATGAGCTCCCATGC | NM_001123710.1 |
| IFN-II | F R | TTCAGGAGACCCAGAAACACTAC TAATGAACTCGGACAGAGCCTTC | AY795563.1 |
| IL-8 | F R | GCAACAGCGGTCAGGAGATT TGGAATGATTCCCCTTCTTCA | HM162835.1 |
| IL-12 | F R | CTGAATGAGGTGGACTGGTATG ATCGTCCTGTTCCTCCG | XM_014205516.1 |
| cd8 | F R | CACTGAGAGAGACGGAAGACG TTCAAAAACCTGCCATAAAGC | AY693393.1 |
| IL-1β | F R | CAAGCTGCCTCAGGGTCT CGGCACCCTTTAACCTCTCC | NM_001123582.1 |
| C3 | F R | TCCCTGGTGGTCACCAGTACAC ATGATGGTGGACTGTGTGGATC | [17] |
| EF-1 | F R | CCCCTCCAGGACGTTTACAAA CACACGGCCCACAGGTACA | NM_001123629.1 |
| Group | Control | PAQ-Xtract |
|---|---|---|
| Initial biomass (kg) | 1619 ± 258 | 1556 ± 410 |
| Biomass at harvest (kg) | 51,634 ± 6691 | 57,200 ± 4527 |
| Economic feed conversion rate | 1.18 ± 0.06 | 1.13 ± 0.04 |
| Biological feed conversion rate | 1.15 ± 0.02 a | 1.09 ± 0.03 b |
| Specific Growth Rate | 0.93 ± 0.01 | 0.94 ± 0.05 |
| Antibiotic treatment | 100.0% | 33.3% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortés, H.; Castillo-Ruiz, M.; Cañon-Jones, H.; Schlotterbeck, T.; San Martín, R.; Padilla, L. In Vivo Efficacy of Purified Quillaja Saponin Extracts in Protecting against Piscirickettsia salmonis Infections in Atlantic Salmon (Salmo salar). Animals 2023, 13, 2845. https://doi.org/10.3390/ani13182845
Cortés H, Castillo-Ruiz M, Cañon-Jones H, Schlotterbeck T, San Martín R, Padilla L. In Vivo Efficacy of Purified Quillaja Saponin Extracts in Protecting against Piscirickettsia salmonis Infections in Atlantic Salmon (Salmo salar). Animals. 2023; 13(18):2845. https://doi.org/10.3390/ani13182845
Chicago/Turabian StyleCortés, Hernán, Mario Castillo-Ruiz, Hernán Cañon-Jones, Trinidad Schlotterbeck, Ricardo San Martín, and Leandro Padilla. 2023. "In Vivo Efficacy of Purified Quillaja Saponin Extracts in Protecting against Piscirickettsia salmonis Infections in Atlantic Salmon (Salmo salar)" Animals 13, no. 18: 2845. https://doi.org/10.3390/ani13182845