
Gamete Collection, Artificial Fertilization and Captive-Rearing of Eggs in a Terrestrial-Breeding Anuran with Parental Care: Alytes obstetricans


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Captive Animals
2.2. Wild Animals
2.3. Experiment 1: Captive-Rearing of Embryos
2.4. Experiment 2: Hormonal Stimulation of Spermiation in Captive and Wild Males
2.5. Experiment 3: Hormonal Stimulation of Spermiation in Captive Males after Semi-Free-Ranging Period
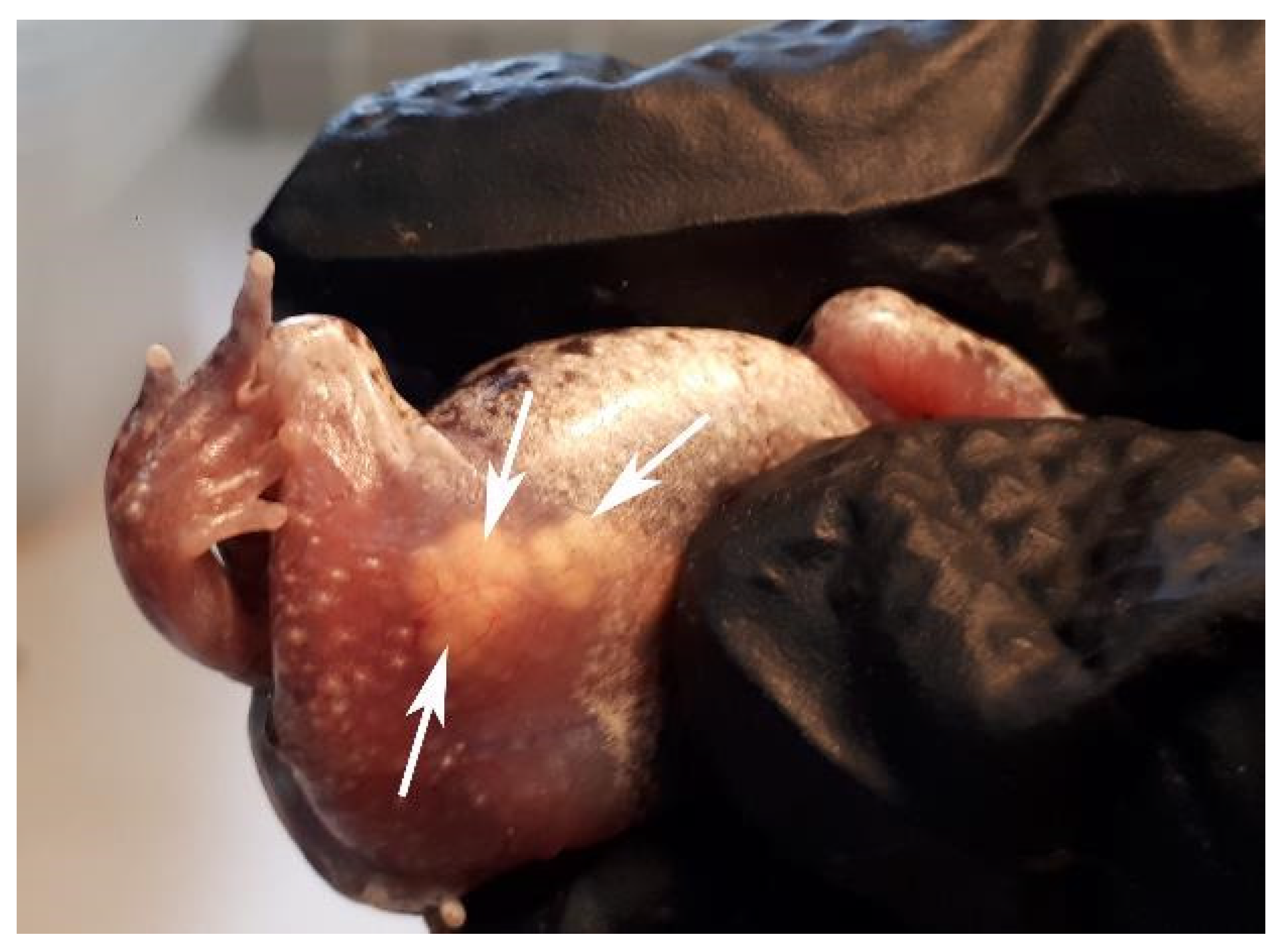
2.6. Experiment 4: Hormonal Stimulation of Ovulation and Artificial Fertilization
2.7. Statistical Analysis
3. Results
3.1. Experiment 1: Captive-Rearing of Embryos
3.2. Experiment 2: Hormonal Stimulation of Spermiation in Captive and Wild Males
3.3. Experiment 3: Hormonal Stimulation of Spermiation in Captive Males after Semi-Free-Ranging Period
3.4. Experiment 4: Hormonal Stimulation of Ovulation and Artificial Fertilization
4. Discussion
- (1)
- The spermiation response using hCG was more effective than using GnRHa. GnRHa was not able to stimulate spermiation at any of the concentrations and routes tested. However, we cannot dismiss the possibility that the concentration of GnRHa administrated exceeds the optimum for this species. hCG and GnRHa have different routes of action that are highly conserved among vertebrates. GnRHa is a synthetic analog of GnRH that is released by the hypothalamus and stimulates the production of luteinizing hormone (LH) and follicle-stimulating hormone (FSH) by the pituitary gland. LH and FSH act in the testes to stimulate testosterone release by Leydig cells and control spermatogenesis through the Sertoli cells, respectively. hCG binds to the same receptors as LH on the Leydig cells [27], and these hormones are involved in spermiation (release of sperm attached to the Sertoli cells) through their action on the Sertoli cells in anurans [28,29]. Species belonging to different families in anurans have shown differences in the sperm-release response triggered by these exogenous hormones [30,31,32,33], but further studies are required to clarify the mechanisms.
- (2)
- No differences in the proportion of responding males were found between intraperitoneal injection and intranasal application of hCG in captive males. Hormone administration in amphibians is commonly performed intraperitoneally or subcutaneously in the dorsal lymph sacs, but alternative non-invasive routes have been tested. Hormones have been administrated topically [34,35,36] and through intranasal application [37]. Although intraperitoneal treatment and intranasal application induced spermiation in 33% and 17% of males, respectively, it would be desirable to develop a reliable non-invasive method for hormonal treatment because it is less invasive for animals and it is easier to perform requiring less training for personnel.
- (3)
- Captivity seems to have a negative effect on sperm quality in Alytes, but this effect could be reverted. Some differences in the percentage of males responding to hormonal stimulation were observed among captive (33%), wild (50%) and captive males after a semi-free-ranging period (43%). Some captive males that did not respond to hormones released sperm after being allowed to range in the outdoor area. Although sperm samples from captive and wild males (that were in captivity for 16 days before treatment) had similar concentrations and percentages of forward motility, the quality of movement of sperm showing forward progression was different. Sperm in some samples from wild males were moving forward in a straight line and at a faster speed than any sample from captive males. It has been proposed that sperm in terrestrial-fertilizing anuran species are immotile, as found in primitive Leiopelmatidae frogs [33] or in foam-nesting Myobatrachidae [38], and motility may be activated through direct contact with the egg [39]. Further studies will be needed to clarify the factors implicated in sperm activation in Alytes. In addition, some samples from captive males that were allowed to range in a semi-free area for a month presented two to four times higher sperm concentration afterward. Similar results were found in Lithobates chiricahuensis (Chiricahua leopard frog), with semi-captive and wild males presenting higher sperm concentration than captive ones while no effect on motility was observed [40].
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silla, A.J.; Kouba, A.J.; Heatwole, H. Reproductive Technologies and Biobanking for the Conservation of Amphibians; Silla, A.J., Kouba, A.J., Heatwole, H., Eds.; CSIRO Publishing: Cayton, VIC, Australia, 2022; Volume 12, pp. 1–9. [Google Scholar]
- González-del-Pliego, P.; Freckleton, R.P.; Edwards, D.P.; Koo, M.S.; Scheffers, B.R.; Pyron, R.A.; Jetz, W. Phylogenetic and trait-based prediction of extinction risk for data-deficient amphibians. Curr. Biol. 2019, 29, 1557–1563.e1553. [Google Scholar] [CrossRef]
- Whittaker, K.; Koo, M.S.; Wake, D.B.; Vredenburg, V.T. Global Declines of Amphibians. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Waltham, MA, USA, 2013; Volume 3, pp. 691–699. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2. Available online: http://www.iucnredlist.org (accessed on 3 January 2022).
- Dufresnes, C.; Martínez-Solano, Í. Hybrid zone genomics supports candidate species in Iberian Alytes obstetricans. Amphib.-Reptil. 2020, 41, 105–112. [Google Scholar] [CrossRef]
- Michaels, C.J.; Bosch, J.; Jervis, P.A.; Márquez, R.; Rosa, G.M. Midwife Toads. The Genus Alytes. History, Ecology, Systematics, Captive Breeding; Edition Chimaira: Frankfurt am Main, Germany, 2023. [Google Scholar]
- Bosch, J.; Martínez-Solano, I.; García-París, M. Evidence of a chytrid fungus infection involved in the decline of the common midwife toad (Alytes obstetricans) in protected areas of central Spain. Biol. Conserv. 2001, 97, 331–337. [Google Scholar] [CrossRef]
- Thumsová, B.; Donaire-Barroso, D.; El Mouden, E.H.; Bosch, J. Fatal chytridiomycosis in the Moroccan midwife toad Alytes maurus and potential distribution of Batrachochytrium dendrobatidis across Morocco. Afr. J. Herpetol. 2022, 71, 72–82. [Google Scholar] [CrossRef]
- Bosch, J.; Mora-Cabello de Alba, A.; Marquínez, S.; Price, S.J.; Thumsová, B.; Bielby, J. Long-Term Monitoring of Amphibian Populations of a National Park in Northern Spain Reveals Negative Persisting Effects of Ranavirus, but Not Batrachochytrium dendrobatidis. Front. Vet. Sci. 2021, 8, 645491. [Google Scholar] [CrossRef]
- Price, S.J.; Garner, T.W.J.; Nichols, R.A.; Balloux, F.; Ayres, C.; Mora-Cabello de Alba, A.; Bosch, J. Collapse of Amphibian Communities Due to an Introduced Ranavirus. Curr. Biol. 2014, 24, 2586–2591. [Google Scholar] [CrossRef]
- Bloxam, Q.M.C.; Tonge, S.J. Amphibians: Suitable candidates for breeding-release programmes. Biodivers. Conserv. 1995, 4, 636–644. [Google Scholar] [CrossRef]
- Serra, J.M.; Griffiths, R.A.; Bosch, J.; Beebee, T.; Schmidt, B.; Tejedo, M.; Lizana, M.; Martínez-Solano, I.; Salvador, A.; García-Paris, M.; et al. Alytes muletensis. The IUCN Red List of Threatened Species. 2009. Available online: http://dx.doi.org/10.2305/IUCN.UK.2009.RLTS.T977A13099129.en (accessed on 28 July 2023).
- Bosch, J.; Sanchez-Tomé, E.; Fernández-Loras, A.; Oliver, J.A.; Fisher, M.C.; Garner, T.W.J. Successful elimination of a lethal wildlife infectious disease in nature. Biol. Lett. 2015, 11, 20150874. [Google Scholar] [CrossRef]
- Woodhams, D.C.; Bosch, J.; Briggs, C.J.; Cashins, S.; Davis, L.R.; Lauer, A.; Muths, E.; Puschendorf, R.; Schmidt, B.R.; Sheafor, B.; et al. Mitigating amphibian disease: Strategies to maintain wild populations and control chytridiomycosis. Front. Zool. 2011, 8, 8. [Google Scholar] [CrossRef]
- Wells, E.; Garcia-Alonso, D.; GonÇalo, M.R.; Garcia, G.; Tapley, B. Amphibian Taxon Advisory Group Best Practice Gudelines for Midwife toads (Alytes sp.). Version 1; EAZA: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Araujo, M.B.; Guilhaumon, F.; Rodrigues Neto, D.; Pozo Ortego, I.; Gomez Calmaestra, R. Impactos, Vulnerabilidad y Adaptación de la Biodiversidad Española Frente al Cambio Climatico; Fauna de Vertebrados: Madrid, Spain, 2011; Volume 2. [Google Scholar]
- Marquez, R.; Verrell, P. The courtship and mating of the Iberian midwife toad Alytes cisternasii (Amphibia: Anura: Discoglossidae). J. Zool. 1991, 225, 125–139. [Google Scholar] [CrossRef]
- Tonge, S.; Bloxam, Q. Breeding the Mallorcan midwife toad Alytes muletensis in captivity. Int. Zoo Yearb. 1989, 28, 45–53. [Google Scholar] [CrossRef]
- García-Paris, M.; Martinez Solano, I. Nuevo estatus taxonómico para las poblaciones ibero-mediterraneas de Alytes obstetricans (Anura: Discoglossidae). Rev. Esp. Herp. 2001, 15, 99–113. [Google Scholar]
- Kouba, A.J.; Vance, C.K.; Willis, E.L. Artificial fertilization for amphibian conservation: Current knowledge and future considerations. Theriogenology 2009, 71, 214–227. [Google Scholar] [CrossRef]
- Nunes-de-Almeida, C.H.L.; Haddad, C.F.B.; Toledo, L.F. A revised classification of the amphibian reproductive modes. Salamandra Geman J. Herpetol. 2021, 57, 413–427. [Google Scholar]
- Duellman, W.E.; Trueb, L. Biology of Amphibians; The Johns Hoppkins University Press: Baltimore, MD, USA; London, UK, 1986. [Google Scholar]
- Lopez Jurado, L.F.; Ruiz Caballero, M.; Dos-Santos Freitas, L. Biología de la reproduccion de Alytes cisternasii Boscá, 1879. Doñana Acta Vertebr. 1979, 6, 5–17. [Google Scholar]
- Márquez, R. Male Reproductive Success in Two Midwife Toads, Alytes obstetricans and A. cisternasii. Behav. Ecol. Sociobiol. 1993, 32, 283–291. [Google Scholar] [CrossRef]
- Pozzi, A.G.; Rosemblit, C.; Ceballos, N.R. Effect of human gonadotropins on spermiation and androgen biosynthesis in the testis of the toad Bufo arenarum (Amphibia, Anura). J. Exp. Zool. Part A Comp. Exp. Biol. 2006, 305 PtA, 96–102. [Google Scholar] [CrossRef]
- Trudeau, V.; Somoza, G.; Natale, G.; Pauli, B.; Wignall, J.; Jackman, P.; Doe, K.; Schueler, F. Hormonal induction of spawning in 4 species of frogs by coinjection with a gonadotropin-releasing hormone agonist and a dopamine antagonist. Reprod. Biol. Endocrinol. 2010, 8, 36. [Google Scholar] [CrossRef]
- Bellisario, R.; Bahl, O.P. Human chorionic gonadotropin. V. Tissue specificity of binding and partial characterization of soluble human chorionic gonadotropin-receptor complexes. J. Biol. Chem. 1975, 250, 3837–3844. [Google Scholar] [CrossRef]
- Burgos, M.H.; Vitale-Calpe, R. The mechanism of spermiation in the toad. Am. J. Anat. 1967, 120, 227–251. [Google Scholar] [CrossRef]
- Russo, J.; Burgos, M.H. Effect of HCG on the enzymic activity of toad testes. Gen. Comp. Endocrinol. 1969, 13, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Kouba, A.; del Barco-Trillo, J.; Vance, C.; Milam, C.; Carr, M. A comparison of human chorionic gonadotropin and luteinizing hormone releasing hormone on the induction of spermiation and amplexus in the American toad (Anaxyrus americanus). Reprod. Biol. Endocrinol. 2012, 10, 59. [Google Scholar] [CrossRef]
- Silla, A.J.; McFadden, M.S.; Byrne, P.G. Hormone-induced sperm-release in the critically endangered Booroolong frog (Litoria booroolongensis): Effects of gonadotropin-releasing hormone and human chorionic gonadotropin. Conserv. Physiol. 2019, 7, coy080. [Google Scholar] [CrossRef] [PubMed]
- Silla, A.J.; Roberts, J.D. Investigating patterns in the spermiation response of eight Australian frogs administered human chorionic gonadotropin (hCG) and luteinizing hormone-releasing hormone (LHRHa). Gen. Comp. Endocrinol. 2012, 179, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Germano, J.M.; Cree, A.; Molinia, F.; Arregui, L.; Bishop, P.J. Hormone treatment does not reliably induce spermiation or mating in Hamilton’s frog from the archaic leiopelmatid lineage. Reprod. Fertil. Dev. 2022, 34, 447–452. [Google Scholar] [CrossRef]
- Glass Campbell, L.; Anderson, K.A.; Marcec-Greaves, R. Topical application of hormone gonadotropin-releasing hormone (GnRH-A) simulates reproduction in the endangered Texas blind salamander (Eurycea rathbuni). Conserv. Sci. Pract. 2021, 4, e609. [Google Scholar] [CrossRef]
- Silla, A.J.; Roberts, J.D.; Byrne, P.G. The effect of injection and topical application of hCG and GnRH agonist to induce sperm-release in the roseate frog, Geocrinia rosea. Conserv. Physiol. 2020, 8, coaa104. [Google Scholar] [CrossRef]
- Rowson, A.D.; Obringer, A.R.; Roth, T.L. Non-invasive treatments luteinizing hormone-releasing hormone for inducing spermiation in American (Bufo americanus) and Gulf Coast (Bufo valliceps) toads. Zoo Biol. 2001, 20, 63–74. [Google Scholar] [CrossRef]
- Julien, A.R.; Kouba, A.J.; Kabelik, D.; Feugang, J.M.; Willard, S.T.; Kouba, C.K. Nasal administration of gonadotropin releasing hormone (GnRH) elicits sperm production in Fowler’s toads (Anaxyrus fowleri). BMC Zool. 2019, 4, 3. [Google Scholar] [CrossRef]
- Edwards, D.L.; Mahony, M.J.; Clulow, J. Effect of sperm concentration, medium osmolality and oocyte storage on artificial fertilisation success in a myobatrachid frog (Limnodynastes tasmaniensis). Reprod. Fertil. Dev. 2004, 16, 347–354. [Google Scholar] [CrossRef]
- Van der Horst, G.; Wilson, B.; Channing, A. Amphibian sperm: Phylogeny and fertilization environment. Mem. Mus. Natl. D’histoire Nat. 1995, 166, 333–342. [Google Scholar]
- Julien, A.R.; Counsell, K.R.; Burger, I.J.; Kouba, A.J.; Barber, D.; Cobos, C.; Jennings, R.D.; Christman, B.L.; Kouba, C.K. Effects of wild, semi-captive, and captive management on male Chiricahua leopard frog sperm quality with implications for conservation breeding programs. Conserv. Sci. Pract. 2023, 5, e12864. [Google Scholar] [CrossRef]
- Browne, R.K.; Silla, A.J.; Upton, R.; Della-Togna, G.; Marcec-Greaves, R.; Shishova, N.V.; Uteshev, V.K.; Proaño, B.; Pérez, O.D.; Mansour, N.; et al. Sperm collection and storage for the sustainable management of amphibian biodiversity. Theriogenology 2019, 133, 187–200. [Google Scholar] [CrossRef]
- Ogielska, M. Reproduction of Amphibians; Science Publishers: Enfield, NH, USA, 2009. [Google Scholar]
- Blüm, V. Comparative Anatomy of the Urogenital System. In Vertebrate Reproduction. A Textbook; Springer: Heidelberg/Berlin, Germany, 1986; pp. 43–47. [Google Scholar]
- Lombardi, J. Urogenital Structure and Integration. In Comparative Vertebrate Reproduction; Springer: New York, NY, USA, 1998; pp. 61–107. [Google Scholar]
- Martin, J.A.; Hillman, S.S. The Physical Movement of Urine from the Kidneys to the Urinary Bladder and Bladder Compliance in Two Anurans. Physiol. Biochem. Zool. Ecol. Evol. Approaches 2009, 82, 163–169. [Google Scholar] [CrossRef]
- Gosner, K. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 1960, 16, 183–190. [Google Scholar]
- Silla, A.J. Artificial fertilisation in a terrestrial toadlet (Pseudophryne guentheri): Effect of medium osmolality, sperm concentration and gamete storage. Reprod. Fertil. Dev. 2013, 25, 1134–1141. [Google Scholar] [CrossRef]
- Browne, R.K.; Clulow, J.; Mahony, M.; Clark, A. Successful recovery of motility and fertility of cryopreserved Cane toad (Bufo marinus) sperm. Cryobiology 1998, 37, 339–345. [Google Scholar] [CrossRef]
- Cabada, M.O. Sperm concentration and fertilization rate in Bufo arenarum (Amphibia: Anura). J. Exp. Biol. 1975, 62, 481–486. [Google Scholar] [CrossRef]
- Levitan, D.R. Sperm Limitation, Gamete Competition and Sexual Selection in External Fertilisers. In Sperm Competition and Sexual Selection; Birkhead, T.R., Möller, A.P., Eds.; Academic Press: London, UK, 1998; pp. 175–218. [Google Scholar]



{kind=link}
{kind=link}
| Hours After Final Treatment | Total Males * | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Priming | Final Treatment | Route | 0 h | 1 h | 2 h | 3 h | 4 h | 5 h | 6 h | |
| none | 0.4 µg GnRHa †/g | nasal | NA | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 |
| none | 0.4 µg GnRHa/g | IP | NA | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 |
| none | 0.8 µg GnRHa/g | IP | NA | 0/4 ‡ | 0/2 ‡ | 0/5 ‡ | 0/3 ‡ | 0/3 ‡ | 0/6 | |
| none | 15 IU hCG/g | IP | NA | 0/5 ‡ | 1/3 ‡ | 2/6 | 2/5 ‡ | 1/2 ‡ | 2/6 | |
| none | 0.8 µg GnRHa + 15 IU hCG/g | IP | NA | 0/4 ‡ | 0/3 ‡ | 2/5 ‡ | 1/4 ‡ | 1/3 ‡ | 2/6 | |
| 0.03 IU hCG/g | 15 IU hCG/g | nasal + IP | 0/2 | 1/2 | 0/2 | 0/2 | 1/2 | |||
| 0.03 IU hCG/g | 0.8 µg GnRHa + 15 IU hCG/g | nasal + IP | 1/2 | 1/2 | 0/2 | 0/2 | 1/2 | |||
| 0.025 µg GnRHa/g | 15 IU hCG/g | nasal + IP | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | |||
| 0.025 µg GnRHa/g | 0.8 µg GnRHa + 15 IU hCG/g | nasal + IP | 0/2 | 0/2 | 0/2 | 0/2 | 0/2 | |||
| Origin | Route | 2 h | 4 h | 6 h | 8 h | 10 h | 12 h | 24 h | Total * |
|---|---|---|---|---|---|---|---|---|---|
| captive | IP | 0/12 | 0/12 | 2/12 | 3/12 | 2/12 | 3/12 | 4/12 | 33% |
| captive | nasal | 0/12 | 2/12 | 1/12 | 1/12 | 1/12 | 0/12 | 0/12 | 17% |
| wild | IP | 0/4 | 1/4 | 1/4 | 1/4 | 2/4 | 2/4 | 1/4 | 50% |
| wild | nasal | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0/4 | 0 |
| Animal | Treatment | 3 h | 6 h | 9 h | 12 h | 24 h |
|---|---|---|---|---|---|---|
| Male 1 | 15 IU hCG/g | 2.2 | 15.2 | 6.7 | 88.0 | 0 |
| Male 2 | 15 IU hCG/g | 0 | 0.1 | 0.3 | 0.4 | 0 |
| Male 3 | 15 IU hCG/g | 3.0 | 9.8 | 49.9 | 52.5 | 30.4 |
| Male 4 | 20 IU hCG/g | 9.1 | 3.5 | 6.4 | 1.5 | 0 |
| Male 5 | 20 IU hCG/g | 0 | 0.2 | 0.4 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arregui, L.; Bosch, J. Gamete Collection, Artificial Fertilization and Captive-Rearing of Eggs in a Terrestrial-Breeding Anuran with Parental Care: Alytes obstetricans. Animals 2023, 13, 2802. https://doi.org/10.3390/ani13172802
Arregui L, Bosch J. Gamete Collection, Artificial Fertilization and Captive-Rearing of Eggs in a Terrestrial-Breeding Anuran with Parental Care: Alytes obstetricans. Animals. 2023; 13(17):2802. https://doi.org/10.3390/ani13172802
Chicago/Turabian StyleArregui, Lucía, and Jaime Bosch. 2023. "Gamete Collection, Artificial Fertilization and Captive-Rearing of Eggs in a Terrestrial-Breeding Anuran with Parental Care: Alytes obstetricans" Animals 13, no. 17: 2802. https://doi.org/10.3390/ani13172802