
Insights on the Evolutionary History and Genetic Patterns of Octopus vulgaris Cuvier, 1797 in the Northeastern Atlantic Using Mitochondrial DNA
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
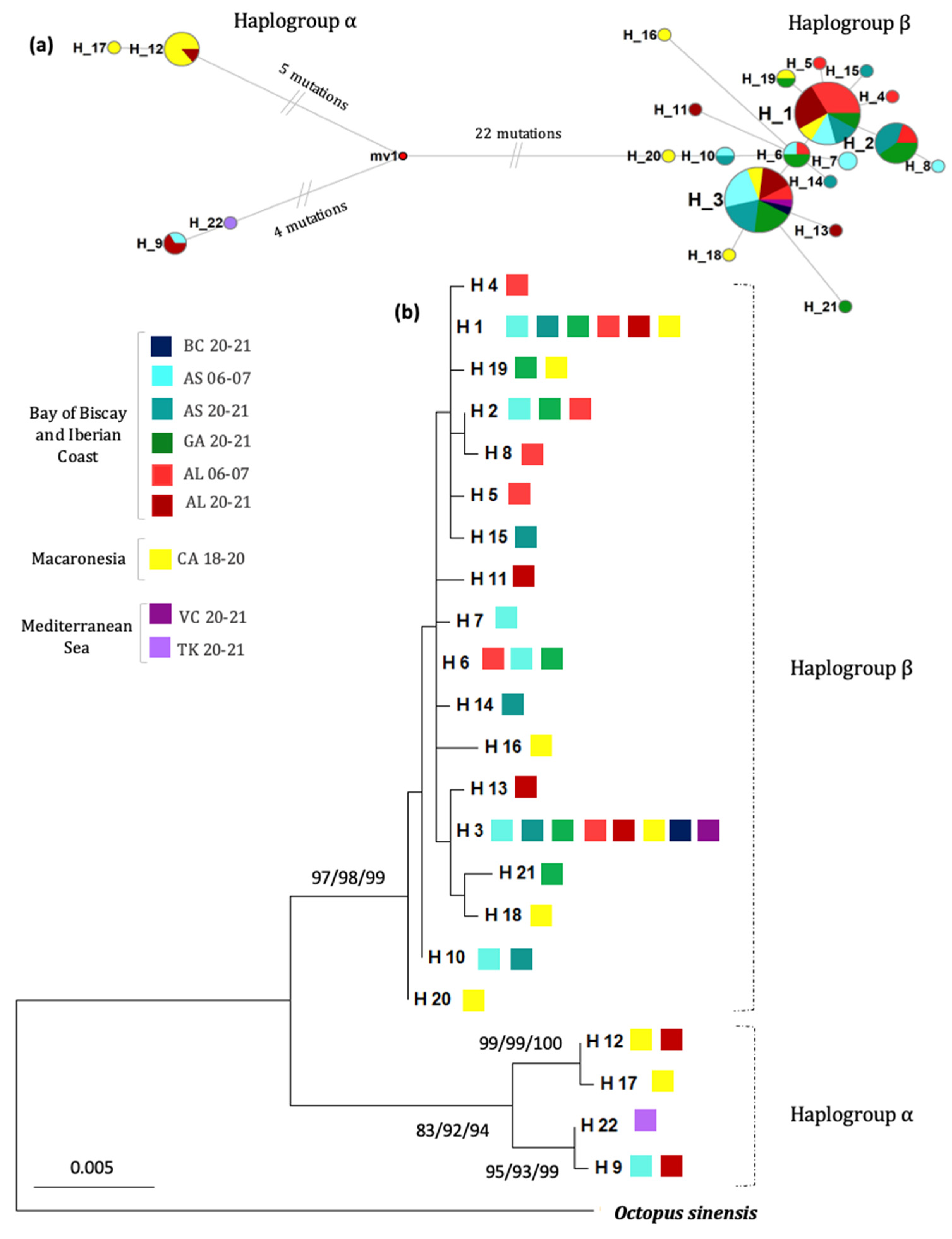
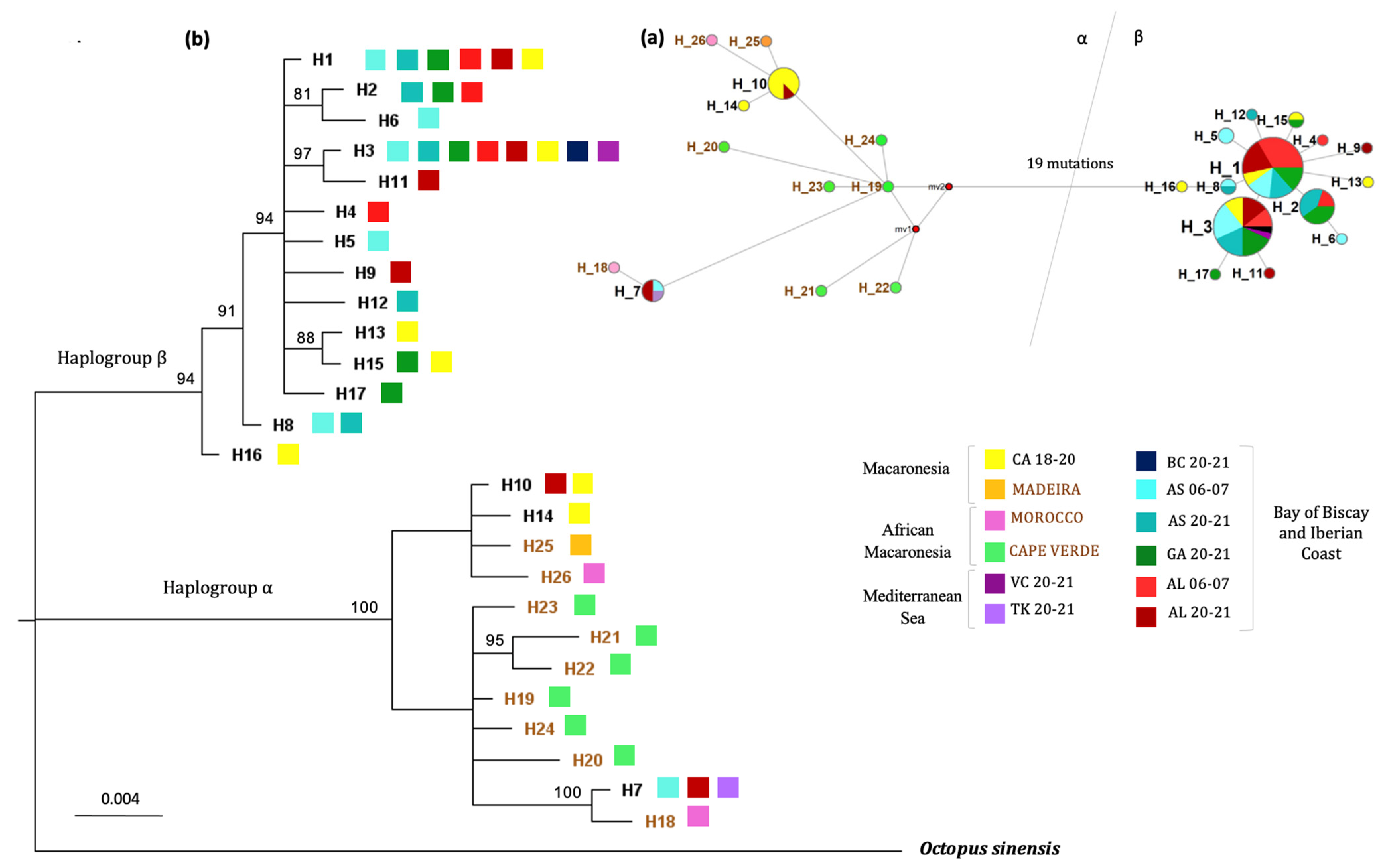
3. Results
4. Discussion
4.1. Demography, Phylogenetic and Phylogeography of O. vulgaris Sensu Stricto
4.2. Current Genetic Structuring Patterns in the O. vulgaris Northeastern Atlantic Populations
4.3. Assessing Temporal Genetic Variation in Exploited Asturian and Algarve O. vulgaris Stocks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Bank The Hidden Harvest. The Global Contribution of Capture Fisheries; World Bank: Washington, DC, USA, 2012. [Google Scholar]
- Villasante, S.; Sumaila, R.; García-Negro, M.; Rodriguez, G. Beijer Discussion Paper Series No. 223; The Beijer Institute of Ecological Economics: Stockholm, Sweden, 2010. [Google Scholar]
- García-de-la-Fuente, L. The Artisanal Fishing Fleet in Asturias: Characterization and Quantitative Analysis of Its Economic and Social Importance; University of Oviedo: Oviedo, Spain, 2020. [Google Scholar]
- FAO. Social Protection for Small-Scale Fisheries in the Mediterranean Region: A Review; FAO: Rome, Italy, 2019; ISBN 978-92-5-131479-1. [Google Scholar]
- Harley, C.D.G.; Randall Hughes, A.; Hultgren, K.M.; Miner, B.G.; Sorte, C.J.B.; Thornber, C.S.; Rodriguez, L.F.; Tomanek, L.; Williams, S.L. The Impacts of Climate Change in Coastal Marine Systems. Ecol. Lett. 2006, 9, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Costello, C.; Ovando, D.; Hilborn, R.; Gaines, S.D.; Deschenes, O.; Lester, S.E. Status and Solutions for the World’s Unassessed Fisheries. Science 2012, 338, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Worm, B.; Hilborn, R.; Baum, J.K.; Branch, T.A.; Collie, J.S.; Costello, C.; Fogarty, M.J.; Fulton, E.A.; Hutchings, J.A.; Jennings, S.; et al. Rebuilding Global Fisheries. Science 2009, 325, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Roa-Ureta, R.H.; del Fernández-Rueda, M.P.; Acuña, J.L.; Rivera, A.; González-Gil, R.; García-Flórez, L. Estimation of the Spawning Stock and Recruitment Relationship of Octopus vulgaris in Asturias (Bay of Biscay) with Generalized Depletion Models: Implications for the Applicability of MSY. ICES J. Mar. Sci. 2021, 78, 2256–2270. [Google Scholar] [CrossRef]
- Arkhipkin, A.I.; Hendrickson, L.C.; Payá, I.; Pierce, G.J.; Roa-Ureta, R.H.; Robin, J.-P.; Winter, A. Stock Assessment and Management of Cephalopods: Advances and Challenges for Short-Lived Fishery Resources. ICES J. Mar. Sci. 2021, 78, 714–730. [Google Scholar] [CrossRef]
- Sánchez, P.; Villanueva, R.; Jereb, P.; Guerra, A.; González, A.F.; Sobrino, I.; Balguerias, E.; Pereira, J.; Moreno, A.; Allcock, A.L.; et al. Octopus Vulgaris Cuvier, 1797. In Cephalopod Biology and Fisheries in Europe: II: Species Accounts; ICES Cooperative Researh, Report; Jereb, P., Allcock, A.L., Lefkaditou, E., Piatkowski, U., Hastie, L.C., Pierce, G.J., Eds.; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2015; pp. 14–28. ISBN 978-87-7482-155-7. [Google Scholar]
- Pita, C.; Roumbedakis, K.; Fonseca, T.; Matos, F.L.; Pereira, J.; Villasante, S.; Pita, P.; Bellido, J.M.; Gonzalez, A.F.; García-Tasende, M.; et al. Fisheries for Common Octopus in Europe: Socioeconomic Importance and Management. Fish. Res. 2021, 235, 105820. [Google Scholar] [CrossRef]
- Fernández Sánchez, J.L.; Fernández Polanco, J.M.; Llorente García, I. Evidence of Price Premium for MSC-Certified Products at Fishers’ Level: The Case of the Artisanal Fleet of Common Octopus from Asturias (Spain). Mar. Policy 2020, 119, 104098. [Google Scholar] [CrossRef]
- González, A.F.; Macho, G.; De Novoa, J.; García, M. Western Asturias Octopus Traps Fishery of Artisanal Cofradias; CAB: BUREAU VERITAS IBERIA: Madrid, Spain, 2016. [Google Scholar]
- Pirhadi, N.; Parrondo, M.; Romero-Bascones, A.; Thoppil, R.; Martínez, J.L.; Fernández-Rueda, M.P.; Márquez, I.; García-Flórez, L.; Dopico, E.; Pérez, T.; et al. Genetic Monitoring on the World’s First MSC Eco-Labeled Common Octopus (O. Vulgaris) Fishery in Western Asturias, Spain. Sci. Rep. 2023, 13, 2730. [Google Scholar] [CrossRef]
- Rios, J.; Macho, G. Western Asturias Octopus Traps Fishery of Artisanal Cofradías; CAB: BUREAU VERITAS IBERIA: Madrid, Spain, 2023; p. 37. [Google Scholar]
- Bhaskar, R.; Sharon, E.A. Molecular Genetic Approaches in Wildlife Conservation. In Genetic Diversity—Recent Advances and Applications; IntechOpen: London, UK, 2022; ISBN 978-1-80356-945-1. [Google Scholar]
- Wright, J.M.; Bentzen, P. Microsatellites: Genetic Markers for the Future. Rev. Fish Biol. Fish. 1994, 4, 384–388. [Google Scholar] [CrossRef]
- Patarnello, T.; Volckaert, F.M.J.; Castilho, R. Pillars of Hercules: Is the Atlantic–Mediterranean Transition a Phylogeographical Break? Mol. Ecol. 2007, 16, 4426–4444. [Google Scholar] [CrossRef]
- Cuéllar-Pinzón, J.; Presa, P.; Hawkins, S.J.; Pita, A. Genetic Markers in Marine Fisheries: Types, Tasks and Trends. Fish. Res. 2016, 173, 194–205. [Google Scholar] [CrossRef]
- Amor, M.D.; Norman, M.D.; Cameron, H.E.; Strugnell, J.M. Allopatric Speciation within a Cryptic Species Complex of Australasian Octopuses. PLoS ONE 2014, 9, e98982. [Google Scholar] [CrossRef]
- Leite, T.S.; Haimovici, M.; Molina, W.; Warnke, K. Morphological and Genetic Description of Octopus Insularis, a New Cryptic Species in the Octopus Vulgaris Complex (Cephalopoda: Octopodidae) from the Tropical Southwestern Atlantic. J. Molluscan Stud. 2008, 74, 63–74. [Google Scholar] [CrossRef]
- Söller, R.; Warnke, K.; Saint-Paul, U.; Blohm, D. Sequence Divergence of Mitochondrial DNA Indicates Cryptic Biodiversity in Octopus vulgaris and Supports the Taxonomic Distinctiveness of Octopus mimus (Cephalopoda: Octopodidae). Mar. Biol. 2000, 136, 29–35. [Google Scholar] [CrossRef]
- Warnke, K.; Söller, R.; Blohm, D.; Saint-Paul, U. A New Look at Geographic and Phylogenetic Relationships within the Species Group Surrounding Octopus vulgaris (Mollusca, Cephalopoda): Indications of Very Wide Distribution from Mitochondrial DNA Sequences. J. Zool. Syst. Evol. Res. 2004, 42, 306–312. [Google Scholar] [CrossRef]
- Sales, J.B.D.L.; Rego, P.S.D.; Hilsdorf, A.W.S.; Moreira, A.A.; Haimovici, M.; Tomás, A.R.; Batista, B.B.; Marinho, R.A.; Markaida, U.; Schneider, H.; et al. Phylogeographical Features of Octopus Vulgaris and Octopus Insularis in the Southeastern Atlantic Based on the Analysis of Mitochondrial Markers. J. Shellfish. Res. 2013, 32, 325–339. [Google Scholar] [CrossRef]
- Avendaño, O.; Roura, Á.; Cedillo-Robles, C.E.; González, Á.F.; Rodríguez-Canul, R.; Velázquez-Abunader, I.; Guerra, Á. Octopus Americanus: A Cryptic Species of the O. Vulgaris Species Complex Redescribed from the Caribbean. Aquat. Ecol. 2020, 54, 909–925. [Google Scholar] [CrossRef]
- Norman, M.D.; Finn, J.K. World octopod fisheries. In Cephalopods of the World. An Annotated and Illustrated Catalogue of Cephalopod Species Known to Date. Octopods and Vampire Squids; FAO Species Catalogue for Fishery Purposes; Jereb, P., Roper, C.F.E., Norman, M.D., Finn, J.K., Eds.; FAO: Rome, Italy, 2016; Volume 3, pp. 9–14. ISSN 1020-8682. [Google Scholar]
- Fadhlaoui-Zid, K.; Knittweis, L.; Aurelle, D.; Nafkha, C.; Ezzeddine, S.; Fiorentino, F.; Ghmati, H.; Ceriola, L.; Jarboui, O.; Maltagliati, F. Genetic Structure of Octopus vulgaris (Cephalopoda, Octopodidae) in the Central Mediterranean Sea Inferred from the Mitochondrial COIII Gene. Comptes Rendus Biol. 2012, 335, 625–636. [Google Scholar] [CrossRef] [PubMed]
- De Luca, D.; Catanese, G.; Procaccini, G.; Fiorito, G. Octopus vulgaris (Cuvier, 1797) in the Mediterranean Sea: Genetic Diversity and Population Structure. PLoS ONE 2016, 11, e0149496. [Google Scholar] [CrossRef]
- Van Nieuwenhove, A.H.M.; Ratsimbazafy, H.A.; Kochzius, M. Cryptic Diversity and Limited Connectivity in Octopuses: Recommendations for Fisheries Management. PLoS ONE 2019, 14, e0214748. [Google Scholar] [CrossRef]
- Quinteiro, J.; Rodríguez-Castro, J.; Rey-Méndez, M.; González-Henríquez, N. Phylogeography of the Insular Populations of Common Octopus, Octopus vulgaris Cuvier, 1797, in the Atlantic Macaronesia. PLoS ONE 2020, 15, e0230294. [Google Scholar] [CrossRef] [PubMed]
- Teske, P.R.; Oosthuizen, A.; Papadopoulos, I.; Barker, N.P. Phylogeographic Structure of Octopus vulgaris in South Africa Revisited: Identification of a Second Lineage near Durban Harbour. Mar. Biol. 2007, 151, 2119–2122. [Google Scholar] [CrossRef]
- Gleadall, I.G. Octopus sinensis d’Orbigny, 1841 (Cephalopoda: Octopodidae): Valid Species Name for the Commercially Valuable East Asian Common Octopus. Species Divers. 2016, 21, 31–42. [Google Scholar] [CrossRef]
- Murphy, J.; Balguerias, E.; Key, L.N.; Boyle, P.R. Microsatellite DNA Markers Discriminate between Two Octopus Vulgaris (Cephalopoda: Octopoda) Fisheries along the Northwest African Coast. Bull. Mar. Sci. 2002, 71, 545–553. [Google Scholar]
- Oosthuizen, A.; Jiwaji, M.; Shaw, P. Genetic Analysis of the Octopus vulgaris Population on the Coast of South Africa. S. Afr. J. Sci. 2004, 100, 603–607. [Google Scholar]
- Cabranes, C.; Fernandez-Rueda, P.; Martínez, J.L. Genetic Structure of Octopus Vulgaris around the Iberian Peninsula and Canary Islands as Indicated by Microsatellite DNA Variation. ICES J. Mar. Sci. 2008, 65, 12–16. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. Bold: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-Based Registry for All Animal Species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Bohonak, A.J. IBD (Isolation by Distance): A Program for Analyses of Isolation by Distance. J. Hered. 2002, 93, 153–154. [Google Scholar] [CrossRef]
- Hasegawa, M.; Yano, T.; Kishino, H. A New Molecular Clock of Mitochondrial DNA and the Evolution of Hominoids. Proc. Jpn. Acad. Ser. B 1984, 60, 95–98. [Google Scholar] [CrossRef]
- Felsenstein, J. Phylogenies and the Comparative Method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Roura, Á.; Amor, M.; González, Á.F.; Guerra, Á.; Barton, E.D.; Strugnell, J.M. Oceanographic Processes Shape Genetic Signatures of Planktonic Cephalopod Paralarvae in Two Upwelling Regions. Prog. Oceanogr. 2019, 170, 11–27. [Google Scholar] [CrossRef]
- Melis, R.; Vacca, L.; Cuccu, D.; Mereu, M.; Cau, A.; Follesa, M.C.; Cannas, R. Genetic Population Structure and Phylogeny of the Common Octopus Octopus vulgaris Cuvier, 1797 in the Western Mediterranean Sea through Nuclear and Mitochondrial Markers. Hydrobiologia 2018, 807, 277–296. [Google Scholar] [CrossRef]
- Jenkins, T.L.; Castilho, R.; Stevens, J.R. Meta-Analysis of Northeast Atlantic Marine Taxa Shows Contrasting Phylogeographic Patterns Following Post-LGM Expansions. PeerJ 2018, 6, e5684. [Google Scholar] [CrossRef]
- Maggs, C.A.; Castilho, R.; Foltz, D.; Henzler, C.; Jolly, M.T.; Kelly, J.; Olsen, J.; Perez, K.E.; Stam, W.; Väinölä, R.; et al. Evaluating Signatures of Glacial Refugia for North Atlantic Benthic Marine Taxa. Ecology 2008, 89, S108–S122. [Google Scholar] [CrossRef]
- Pérez-Losada, M.; Guerra, Á.; Sanjuan, A. Allozyme Differentiation in the Cuttlefish Sepia Officinalis (Mollusca: Cephalopoda) from the NE Atlantic and Mediterranean. Heredity 1999, 83, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, R.; Vidal, E.A.G.; Fernández-Álvarez, F.Á.; Nabhitabhata, J. Early Mode of Life and Hatchling Size in Cephalopod Molluscs: Influence on the Species Distributional Ranges. PLoS ONE 2016, 11, e0165334. [Google Scholar] [CrossRef] [PubMed]
- Barton, E.D.; Arístegui, J.; Tett, P.; Navarro-Pérez, E. Variability in the Canary Islands Area of Filament-Eddy Exchanges. Prog. Oceanogr. 2004, 62, 71–94. [Google Scholar] [CrossRef]
- Quinteiro, J.; Rodríguez-Castro, J.; Rey-Méndez, M. Population Genetic Structure of the Stalked Barnacle Pollicipes Pollicipes (Gmelin, 1789) in the Northeastern Atlantic: Influence of Coastal Currents and Mesoscale Hydrographic Structures. Mar. Biol. 2007, 153, 47–60. [Google Scholar] [CrossRef]
- Puerta, P.; Quetglas, A.; Hidalgo, M. Seasonal Variability of Cephalopod Populations: A Spatio-Temporal Approach in the Western Mediterranean Sea. Fish. Oceanogr. 2016, 25, 373–389. [Google Scholar] [CrossRef]
- Sobrino, I.; Rueda, L.; Tugores, M.P.; Burgos, C.; Cojan, M.; Pierce, G.J. Abundance Prediction and Influence of Environmental Parameters in the Abundance of Octopus (Octopus vulgaris Cuvier, 1797) in the Gulf of Cadiz. Fish. Res. 2020, 221, 105382. [Google Scholar] [CrossRef]
- Otero, J.; Álvarez-Salgado, X.A.; González, Á.F.; Miranda, A.; Groom, S.B.; Cabanas, J.M.; Casas, G.; Wheatley, B.; Guerra, Á. Bottom-up Control of Common Octopus Octopus vulgaris in the Galician Upwelling System, Northeast Atlantic Ocean. Mar. Ecol. Prog. Ser. 2008, 362, 181–192. [Google Scholar] [CrossRef]
- Moreno, A.; Lourenço, S.; Pereira, J.; Gaspar, M.B.; Cabral, H.N.; Pierce, G.J.; Santos, A.M.P. Essential habitats for pre-recruit Octopus vulgaris along the Portuguese coast. Fisheries Res. 2014, 152, 74–85. [Google Scholar] [CrossRef]
- Calderón, I.; Pita, L.; Brusciotti, S.; Palacín, C.; Turon, X. Time and Space: Genetic Structure of the Cohorts of the Common Sea Urchin Paracentrotuslividus in Western Mediterranean. Mar. Biol. 2012, 159, 187–197. [Google Scholar] [CrossRef]
- Baptista, V.; Silva, P.L.; Relvas, P.; Teodósio, M.A.; Leitão, F. Sea Surface Temperature Variability along the Portuguese Coast since 1950. Int. J. Climatol. 2018, 38, 1145–1160. [Google Scholar] [CrossRef]
- Leitão, F.; Maharaj, R.R.; Vieira, V.M.N.C.S.; Teodósio, A.; Cheung, W.W.L. The Effect of Regional Sea Surface Temperature Rise on Fisheries along the Portuguese Iberian Atlantic Coast. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 1351–1359. [Google Scholar] [CrossRef]
- Perales-Raya, C.; Nande, M.; Roura, A.; Bartolomé, A.; Gestal, C.; Otero, J.J.; García-Fernández, P.; Almansa, E. Comparative Study of Age Estimation in Wild and Cultured Octopus vulgaris Paralarvae: Effect of Temperature and Diet. Mar. Ecol. Prog. Ser. 2018, 598, 247–259. [Google Scholar] [CrossRef]
- Parrondo, M.; Morán, P.; Ballenghien, M.; Acuña, J.L.; Aguión, A.; Arrontes, J.; Chiss, J.; Cruz, T.; Fernandes, J.N.; García-Flórez, L.; et al. Chaotic Genetic Patchiness in the Highly Valued Atlantic Stalked Barnacle Pollicipes Pollicipes From the Iberian Peninsula: Implications for Fisheries Management. Front. Mar. Sci. 2022, 9, 801780. [Google Scholar] [CrossRef]
- Dietrich, D.E.; Tseng, Y.-H.; Medina, R.; Piacsek, S.A.; Liste, M.; Olabarrieta, M.; Bowman, M.J.; Mehra, A. Mediterranean Overflow Water (MOW) Simulation Using a Coupled Multiple-Grid Mediterranean Sea/North Atlantic Ocean Model. J. Geophys. Res. Ocean. 2008, 113. [Google Scholar] [CrossRef]
- Silva, L.; Sobrino, I.; Ramos, F. Reproductive Biology of the Common Octopus, Octopus vulgaris Cuvier, 1797 (Cephalopoda: Octopodidae) in the Gulf of Cádiz (SW Spain). Bull. Mar. Sci. 2002, 71, 837–850. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
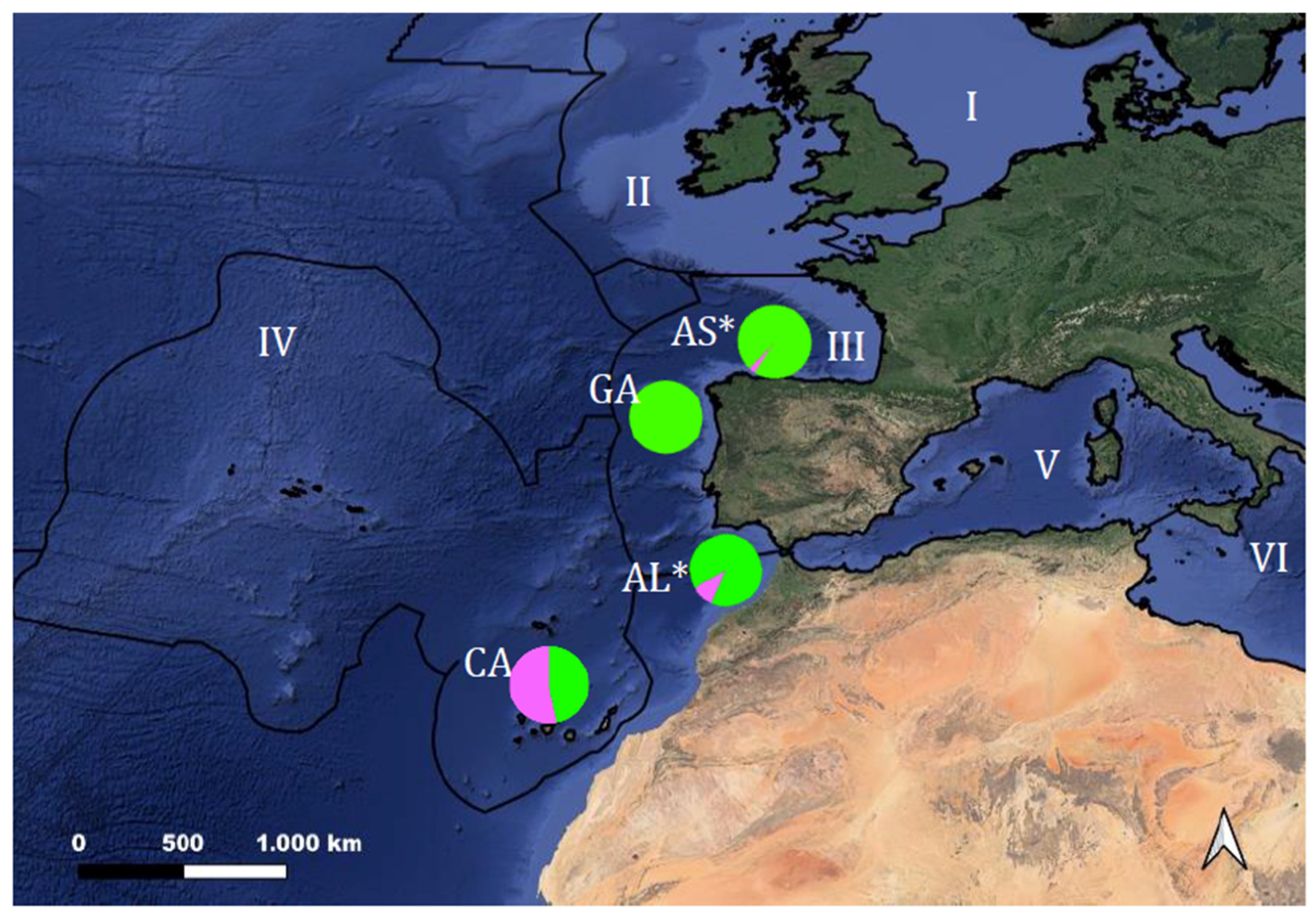
| Sampling Region | Locality | Code | Coordinates | FAO Division | Fishing Season | n |
|---|---|---|---|---|---|---|
| Asturias 1 Cantabrian Sea | Puerto de Vega | AS | 43.566/−6.633 | 27.8. c | 2006–2007 2020–2021 | 15 15 |
| Galicia 1 Northeastern Atlantic | Bueu | GA | 42.363/−8.851 | 27.9. a | 2020–2021 | 15 |
| Algarve 1 Northeastern Atlantic | Olhão | AL | 36.947/−7.954 | 27.9. a | 2006–2007 2020–2021 | 15 15 |
| Canary Islands 2 Northeastern Atlantic | San Andrés | CA | 28.616/−16.335 | 34.1.2 | 2018–2020 | 15 |
| Cytochrome Oxidase 1 (cox1) | Control Region (CR) | Concatenated (cox1 + CR) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Population | N | Nh | hs | h | π | k | Nh | hs | h | π | k | Nh | hs | h | π | k |
| Asturias 2006–2007 | 15 | 3 | 0 | 0.514 | 0.00168 | 1.086 | 6 | 2 | 0.790 | 0.00633 | 4.095 | 7 | 2 | 0.819 | 0.00401 | 5.181 |
| Asturias 2020–2021 | 15 | 3 | 1 | 0.600 | 0.00103 | 0.667 | 5 | 1 | 0.790 | 0.00179 | 1.162 | 6 | 2 | 0.819 | 0.00141 | 1.829 |
| Galicia 2020–2021 | 15 | 3 | 0 | 0.600 | 0.00103 | 0.667 | 6 | 1 | 0.819 | 0.00205 | 1.333 | 6 | 1 | 0.838 | 0.00154 | 2.000 |
| Algarve 2006–2007 | 15 | 3 | 1 | 0.448 | 0.00074 | 0.476 | 4 | 1 | 0.552 | 0.00097 | 0.629 | 6 | 2 | 0.714 | 0.00085 | 1.105 |
| Algarve 2020–2021 | 15 | 3 | 0 | 0.705 | 0.00404 | 2.610 | 6 | 2 | 0.790 | 0.01330 | 8.629 | 6 | 2 | 0.790 | 0.00868 | 11.238 |
| Canary Islands 2018–2020 | 15 | 5 | 1 | 0.743 | 0.00590 | 3.810 | 7 | 3 | 0.819 | 0.01966 | 12.743 | 8 | 4 | 0.838 | 0.01279 | 16.552 |
| Asturias 2006–2007 | Asturias 2020–2021 | Galicia 2020–2021 | Algarve 2006–2007 | Algarve 2020–2021 | Canary Islands 2018–2021 | |
|---|---|---|---|---|---|---|
| Asturias 2006–2007 | 0.25304 | 0.34254 | 0.00861 | 0.53163 | 0.00386 | |
| Asturias 2020–2021 | 0.01038 | 0.99990 | 0.21156 | 0.11256 | 0.00129 | |
| Galicia 2020–2021 | 0.00265 | −0.04978 | 0.10068 | 0.10692 | 0.00109 | |
| Algarve 2006–2007 | 0.09957 * | 0.03509 | 0.07656 | 0.01346 | 0.00000 | |
| Algarve 2020–2021 | −0.00602 | 0.09650 | 0.09608 | 0.13592 * | 0.05148 | |
| Canary Islands 2018–2021 | 0.27796 * | 0.38857 * | 0.38857 * | 0.41558 * | 0.11231 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez, T.; Romero-Bascones, A.; Pirhadi, N.; Coya, R.; Fernández-Rueda, M.d.P.; Márquez, I.; García-Flórez, L.; Borrell, Y.J. Insights on the Evolutionary History and Genetic Patterns of Octopus vulgaris Cuvier, 1797 in the Northeastern Atlantic Using Mitochondrial DNA. Animals 2023, 13, 2708. https://doi.org/10.3390/ani13172708
Pérez T, Romero-Bascones A, Pirhadi N, Coya R, Fernández-Rueda MdP, Márquez I, García-Flórez L, Borrell YJ. Insights on the Evolutionary History and Genetic Patterns of Octopus vulgaris Cuvier, 1797 in the Northeastern Atlantic Using Mitochondrial DNA. Animals. 2023; 13(17):2708. https://doi.org/10.3390/ani13172708
Chicago/Turabian StylePérez, Trinidad, Andrea Romero-Bascones, Negin Pirhadi, Ruth Coya, María del Pino Fernández-Rueda, Isabel Márquez, Lucía García-Flórez, and Yaisel J. Borrell. 2023. "Insights on the Evolutionary History and Genetic Patterns of Octopus vulgaris Cuvier, 1797 in the Northeastern Atlantic Using Mitochondrial DNA" Animals 13, no. 17: 2708. https://doi.org/10.3390/ani13172708