
Cobalt Protoporphyrin Blocks EqHV-8 Infection via IFN-α/β Production
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, Antibodies, and Chemicals
2.2. CoPP Cytotoxicity
2.3. Inhibition Assay of CoPP In Vitro
2.4. Effects of CoPP on EqHV-8 Infection
2.5. The Change of IFN Response Treated with CoPP
2.6. Animal Experiments
2.6.1. Histopathology Evaluation
2.6.2. Virus Replication in Tissues
2.7. Gene Transcription and Immunoblot Analysis
2.7.1. Real-Time Quantitative PCR (qPCR)
2.7.2. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. Chemical Structure and CoPP Cytotoxicity
3.2. Inhibition Assay of CoPP In Vitro
3.3. Effects of CoPP on EqHV-8 Infection
3.4. The Change of IFN Response Treated with CoPP
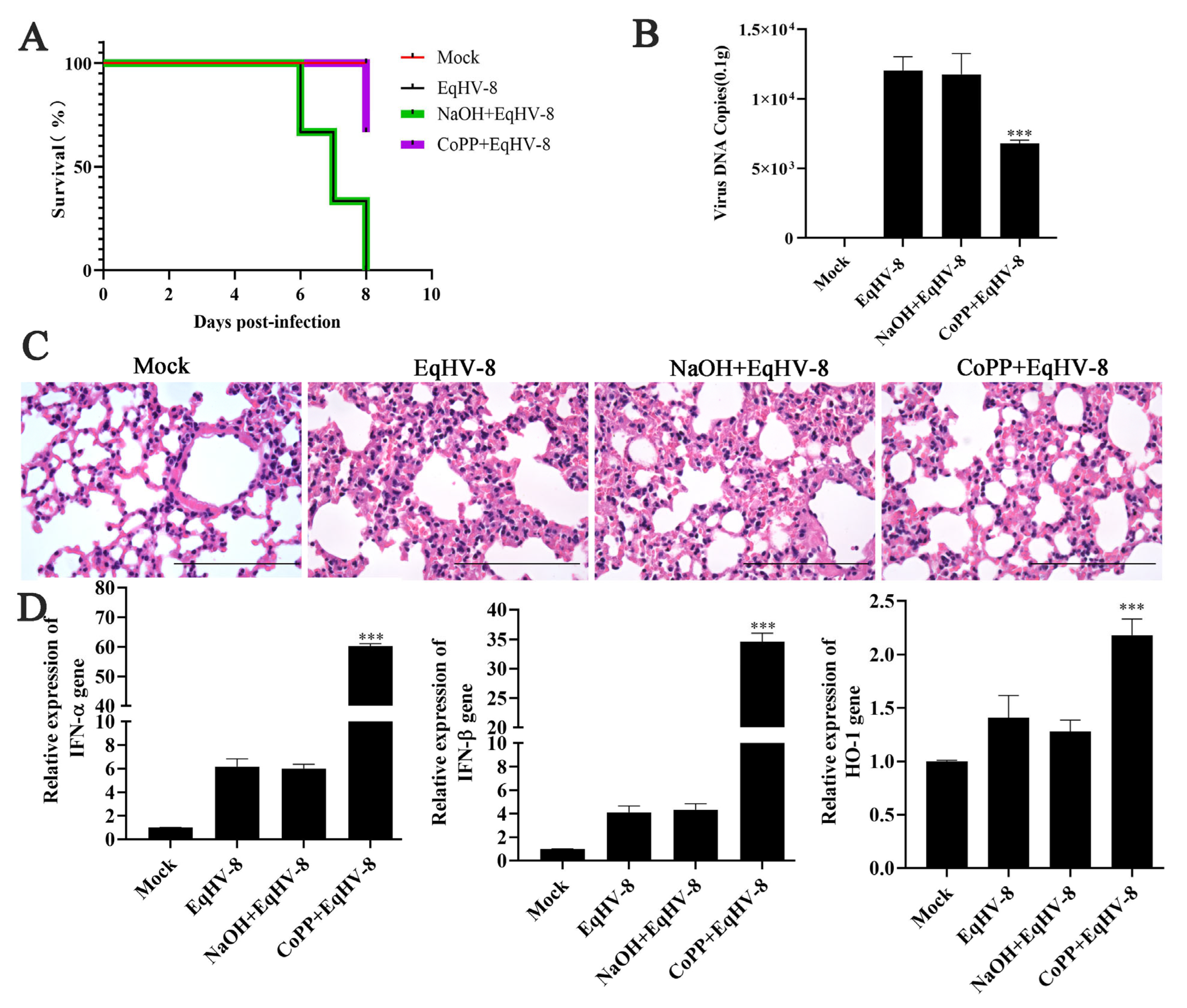
3.5. CoPP Decreased EqHV-8 Infection in Mice Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, C.; Guo, W.; Lu, G.; Xiang, W.; Wang, X. Complete genomic sequence of an equine herpesvirus type 8 Wh strain isolated from China. J. Virol. 2012, 86, 5407. [Google Scholar] [CrossRef]
- Garvey, M.; Suarez, N.M.; Kerr, K.; Hector, R.; Moloney-Quinn, L.; Arkins, S.; Davison, A.J.; Cullinane, A. Equid herpesvirus 8: Complete genome sequence and association with abortion in mares. PLoS ONE 2018, 13, e0192301. [Google Scholar] [CrossRef]
- Schvartz, G.; Edery, N.; Moss, L.; Hadad, R.; Steinman, A.; Karniely, S. Equid Herpesvirus 8 Isolated From an Adult Donkey in Israel. J. Equine Vet. Sci. 2020, 94, 103247. [Google Scholar] [CrossRef]
- Wang, T.; Hu, L.; Wang, Y.; Liu, W.; Liu, G.; Zhu, M.; Zhang, W.; Wang, C.; Ren, H.; Li, L. Identification of equine herpesvirus 8 in donkey abortion: A case report. Virol. J. 2022, 19, 10. [Google Scholar] [CrossRef]
- Takeda, T.A.; Sasai, M.; Adachi, Y.; Ohnishi, K.; Fujisawa, J.I.; Izawa, S.; Taketani, S. Potential role of heme metabolism in the inducible expression of heme oxygenase-1. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1813–1824. [Google Scholar] [CrossRef]
- Negi, G.; Nakkina, V.; Kamble, P.; Sharma, S.S. Heme oxygenase-1, a novel target for the treatment of diabetic complications: Focus on diabetic peripheral neuropathy. Pharmacol. Res. 2015, 102, 158–167. [Google Scholar] [CrossRef]
- Tao, Y.; Cai, L.; Zhou, D.; Wang, C.; Ma, Z.; Dong, X.; Peng, G. CoPP-Induced-Induced HO-1 Overexpression Alleviates Photoreceptor Degeneration With Rapid Dynamics: A Therapeutic Molecular Against Retinopathy. Investig. Ophthalmol. Vis. Sci. 2019, 60, 5080–5094. [Google Scholar] [CrossRef]
- Yang, W.; Wang, Y.; Zhang, C.; Huang, Y.; Yu, J.; Shi, L.; Zhang, P.; Yin, Y.; Li, R.; Tao, K. Maresin1 Protect Against Ferroptosis-Induced Liver Injury Through ROS Inhibition and Nrf2/HO-1/GPX4 Activation. Front. Pharmacol. 2022, 13, 865689. [Google Scholar] [CrossRef]
- Feng, Y.; Guo, X.; Tian, H.; He, Y.; Li, Y.; Jiang, X.; Zheng, H.; Xiao, S. Induction of HOXA3 by Porcine Reproductive and Respiratory Syndrome Virus Inhibits Type I Interferon Response through Negative Regulation of HO-1 Transcription. J. Virol. 2022, 96, e0186321. [Google Scholar] [CrossRef]
- Wu, C.C.; Lu, K.C.; Chen, J.S.; Hsieh, H.Y.; Lin, S.H.; Chu, P.; Wang, J.Y.; Sytwu, H.K.; Lin, Y.F. HO-1 induction ameliorates experimental murine membranous nephropathy: Anti-oxidative, anti-apoptotic and immunomodulatory effects. Nephrol. Dial. Transplant. 2008, 23, 3082–3090. [Google Scholar] [CrossRef]
- Liu, X.M.; Durante, Z.E.; Peyton, K.J.; Durante, W. Heme oxygenase-1-derived bilirubin counteracts HIV protease inhibitor-mediated endothelial cell dysfunction. Free Radic. Biol. Med. 2016, 94, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.A.; Leon, M.A.; Cespedes, P.F.; Gomez, R.S.; Canedo-Marroquin, G.; Riquelme, S.A.; Salazar-Echegarai, F.J.; Blancou, P.; Simon, T.; Anegon, I.; et al. Heme Oxygenase-1 Modulates Human Respiratory Syncytial Virus Replication and Lung Pathogenesis during Infection. J. Immunol. 2017, 199, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, W.N.; Mathahs, M.M.; Zhu, Z. Heme and HO-1 Inhibition of HCV, HBV, and HIV. Front. Pharmacol. 2012, 3, 129. [Google Scholar] [CrossRef] [PubMed]
- Hill-Batorski, L.; Halfmann, P.; Neumann, G.; Kawaoka, Y. The cytoprotective enzyme heme oxygenase-1 suppresses Ebola virus replication. J. Virol. 2013, 87, 13795–13802. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Pu, F.; Zhang, A.; Xu, L.; Li, N.; Yan, Y.; Gao, J.; Liu, H.; Zhang, G.; Goodfellow, I.G.; et al. Heme Oxygenase-1 Suppresses Bovine Viral Diarrhoea Virus Replication in vitro. Sci. Rep. 2015, 5, 15575. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, A.; Zhang, C.; Ni, H.; Gao, J.; Wang, C.; Zhao, Q.; Wang, X.; Wang, X.; Ma, C.; et al. Heme oxygenase-1 acts as an antiviral factor for porcine reproductive and respiratory syndrome virus infection and over-expression inhibits virus replication in vitro. Antivir. Res. 2014, 110, 60–69. [Google Scholar] [CrossRef]
- Kim, D.H.; Ahn, H.S.; Go, H.J.; Kim, D.Y.; Kim, J.H.; Lee, J.B.; Park, S.Y.; Song, C.S.; Lee, S.W.; Choi, I.S. Heme Oxygenase-1 Exerts Antiviral Activity against Hepatitis A Virus In Vitro. Pharmaceutics 2021, 13, 1229. [Google Scholar] [CrossRef]
- Matsumoto, M.; Funami, K.; Tanabe, M.; Oshiumi, H.; Shingai, M.; Seto, Y.; Yamamoto, A.; Seya, T. Subcellular localization of Toll-like receptor 3 in human dendritic cells. J. Immunol. 2003, 171, 3154–3162. [Google Scholar] [CrossRef]
- Samuel, C.E. Antiviral actions of interferons. Clin. Microbiol. Rev. 2001, 14, 778–809. [Google Scholar] [CrossRef]
- Diamond, M.S.; Harris, E. Interferon inhibits dengue virus infection by preventing translation of viral RNA through a PKR-independent mechanism. Virology 2001, 289, 297–311. [Google Scholar] [CrossRef]
- Jones, M.; Davidson, A.; Hibbert, L.; Gruenwald, P.; Schlaak, J.; Ball, S.; Foster, G.R.; Jacobs, M. Dengue virus inhibits alpha interferon signaling by reducing STAT2 expression. J. Virol. 2005, 79, 5414–5420. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.M.; Kim, H.; Lee, S.A.; Lee, S.Y.; Kim, B.J. A Telomerase-Derived Peptide Exerts an Anti-Hepatitis B Virus Effect via Mitochondrial DNA Stress-Dependent Type I Interferon Production. Front. Immunol. 2020, 11, 652. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.; Vasconcelos, L.R.; Travassos, L.H. The different facets of heme-oxygenase 1 in innate and adaptive immunity. Cell Biochem. Biophys. 2022, 80, 609–631. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Wang, T.; Ren, H.; Liu, W.; Li, Y.; Wang, C.; Li, L. Characterizing the Pathogenesis and Immune Response of Equine Herpesvirus 8 Infection in Lung of Mice. Animals 2022, 12, 2495. [Google Scholar] [CrossRef]
- Li, L.; Sun, W.; Hu, Q.; Wang, T.; Zhu, G.; Zhao, Q.; Zhou, E.M. Identification of MYH9 Key Domain Involved in the Entry of PRRSV Into Permissive Cells. Front. Microbiol. 2022, 13, 865343. [Google Scholar] [CrossRef]
- Tzima, S.; Victoratos, P.; Kranidioti, K.; Alexiou, M.; Kollias, G. Myeloid heme oxygenase-1 regulates innate immunity and autoimmunity by modulating IFN-beta production. J. Exp. Med. 2009, 206, 1167–1179. [Google Scholar] [CrossRef]
- Ma, L.L.; Wang, H.Q.; Wu, P.; Hu, J.; Yin, J.Q.; Wu, S.; Ge, M.; Sun, W.F.; Zhao, J.Y.; Aisa, H.A.; et al. Rupestonic acid derivative YZH-106 suppresses influenza virus replication by activation of heme oxygenase-1-mediated interferon response. Free Radic. Biol. Med. 2016, 96, 347–361. [Google Scholar] [CrossRef]
- Ma, L.L.; Zhang, P.; Wang, H.Q.; Li, Y.F.; Hu, J.; Jiang, J.D.; Li, Y.H. heme oxygenase-1 agonist CoPP suppresses influenza virus replication through IRF3-mediated generation of IFN-alpha/beta. Virology 2019, 528, 80–88. [Google Scholar] [CrossRef]
- Wang, T.; Hu, L.; Liu, M.; Wang, T.; Hu, X.; Li, Y.; Liu, W.; Li, Y.; Wang, Y.; Ren, H.; et al. The Emergence of Viral Encephalitis in Donkeys by Equid Herpesvirus 8 in China. Front. Microbiol. 2022, 13, 840754. [Google Scholar] [CrossRef]
- Browning, G.F.; Ficorilli, N.; Studdert, M.J. Asinine herpesvirus genomes: Comparison with those of the equine herpesviruses. Arch. Virol. 1988, 101, 183–190. [Google Scholar] [CrossRef]
- Rubio-Atonal, L.F.; Serrano-Garcia, N.; Limon-Pacheco, J.H.; Pedraza-Chaverri, J.; Orozco-Ibarra, M. Cobalt protoporphyrin decreases food intake, body weight, and the number of neurons in the Nucleus Accumbens in female rats. Brain Res. 2021, 1758, 147337. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, L.; Jin, L.; Yuan, J. Heme Oxygenase-1 inhibits spring viremia of carp virus replication through carbon monoxide mediated cyclic GMP/Protein kinase G signaling pathway. Fish. Shellfish Immunol. 2018, 79, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yuyin, D.; Fengyang, C.; Xukang, Z.; Jianliang, L. Heme Oxygenase-1 suppresses duck Tembusu virus replication in vitro. Vet. Microbiol. 2020, 251, 108885. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Wan, B.; Jiang, D.; Wu, Y.; Ji, P.; Du, Y.; Zhang, G. The Cytoprotective Enzyme Heme Oxygenase-1 Suppresses Pseudorabies Virus Replication in vitro. Front. Microbiol. 2020, 11, 412. [Google Scholar] [CrossRef] [PubMed]
- El Kalamouni, C.; Frumence, E.; Bos, S.; Turpin, J.; Nativel, B.; Harrabi, W.; Wilkinson, D.A.; Meilhac, O.; Gadea, G.; Desprès, P.; et al. Subversion of the Heme Oxygenase-1 Antiviral Activity by Zika Virus. Viruses 2018, 11, 2. [Google Scholar] [CrossRef]
- Toro, A.; Ruiz, M.S.; Lage-Vickers, S.; Sanchis, P.; Sabater, A.; Pascual, G.; Seniuk, R.; Cascardo, F.; Ledesma-Bazan, S.; Vilicich, F.; et al. A Journey into the Clinical Relevance of Heme Oxygenase 1 for Human Inflammatory Disease and Viral Clearance: Why Does It Matter on the COVID-19 Scene? Antioxidants 2022, 11, 276. [Google Scholar] [CrossRef]
- Tseng, C.K.; Lin, C.K.; Wu, Y.H.; Chen, Y.H.; Chen, W.C.; Young, K.C.; Lee, J.C. Human heme oxygenase 1 is a potential host cell factor against dengue virus replication. Sci. Rep. 2016, 6, 32176. [Google Scholar] [CrossRef]
- Protzer, U.; Seyfried, S.; Quasdorff, M.; Sass, G.; Svorcova, M.; Webb, D.; Bohne, F.; Hösel, M.; Schirmacher, P.; Tiegs, G. Antiviral activity and hepatoprotection by heme oxygenase-1 in hepatitis B virus infection. Gastroenterology 2007, 133, 1156–1165. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequences (5′-3′) |
|---|---|
| HO-1-F | AGTTCATGAAGAACTTTCA |
| HO-1-R | TACCAGAAGGCCATGTCC |
| EqHV-8 186-F | CCCACGTGTGCAACGCCTAT |
| EqHV-8 186-R | ATACAGTCCCGAGGCAGAGT |
| EqHV-8-gD-F | GATGCCAAACCGAATCAGCC |
| EqHV-8-gD-R | TAGGCGAGTCAAGCCGTTTT |
| IFN-α-F | TACTCAGCAGACCTTGAACCT |
| IFN-α-R | CAGTATTGGCAGCAAGTTGAC |
| IFN-β-F | AGCTCCAAGAAAGGACGAACAT |
| IFN-β-R | GCCCTGTAGGTGAGGTTGATCT |
| OAS1-F | GGAGGCGGTTGGCTGAAGAGG |
| OAS1-R | GAACCACCGTCGGCACATCC |
| PKR-F | CGTTTCTTGCCTCCTGCTTTG |
| PKR-R | GGGACCTCCACATGACAGAAG |
| GAPDH-F | CCTTCCGTGTCCCTACTGCCAAC |
| GAPDH-R | GACGCCTGCTTCACCACCTTCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Hu, X.; Li, S.; Li, Y.; Zhao, S.; Shen, F.; Wang, C.; Li, Y.; Wang, T. Cobalt Protoporphyrin Blocks EqHV-8 Infection via IFN-α/β Production. Animals 2023, 13, 2690. https://doi.org/10.3390/ani13172690
Li L, Hu X, Li S, Li Y, Zhao S, Shen F, Wang C, Li Y, Wang T. Cobalt Protoporphyrin Blocks EqHV-8 Infection via IFN-α/β Production. Animals. 2023; 13(17):2690. https://doi.org/10.3390/ani13172690
Chicago/Turabian StyleLi, Liangliang, Xinyao Hu, Shuwen Li, Ying Li, Shengmiao Zhao, Fengzhen Shen, Changfa Wang, Yubao Li, and Tongtong Wang. 2023. "Cobalt Protoporphyrin Blocks EqHV-8 Infection via IFN-α/β Production" Animals 13, no. 17: 2690. https://doi.org/10.3390/ani13172690