

A Study on Differential Biomarkers in the Milk of Holstein Cows with Different Somatic Cells Count Levels
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Samples Collection
2.2. Milk Microorganism DNA Extraction, PCR Amplification, and Sequencing
2.3. Metabolomics Analysis
2.4. Data Processing and Analyses
3. Results
3.1. Routine Analysis of Milk Composition
3.2. Quality Control Results of Microbiological Group
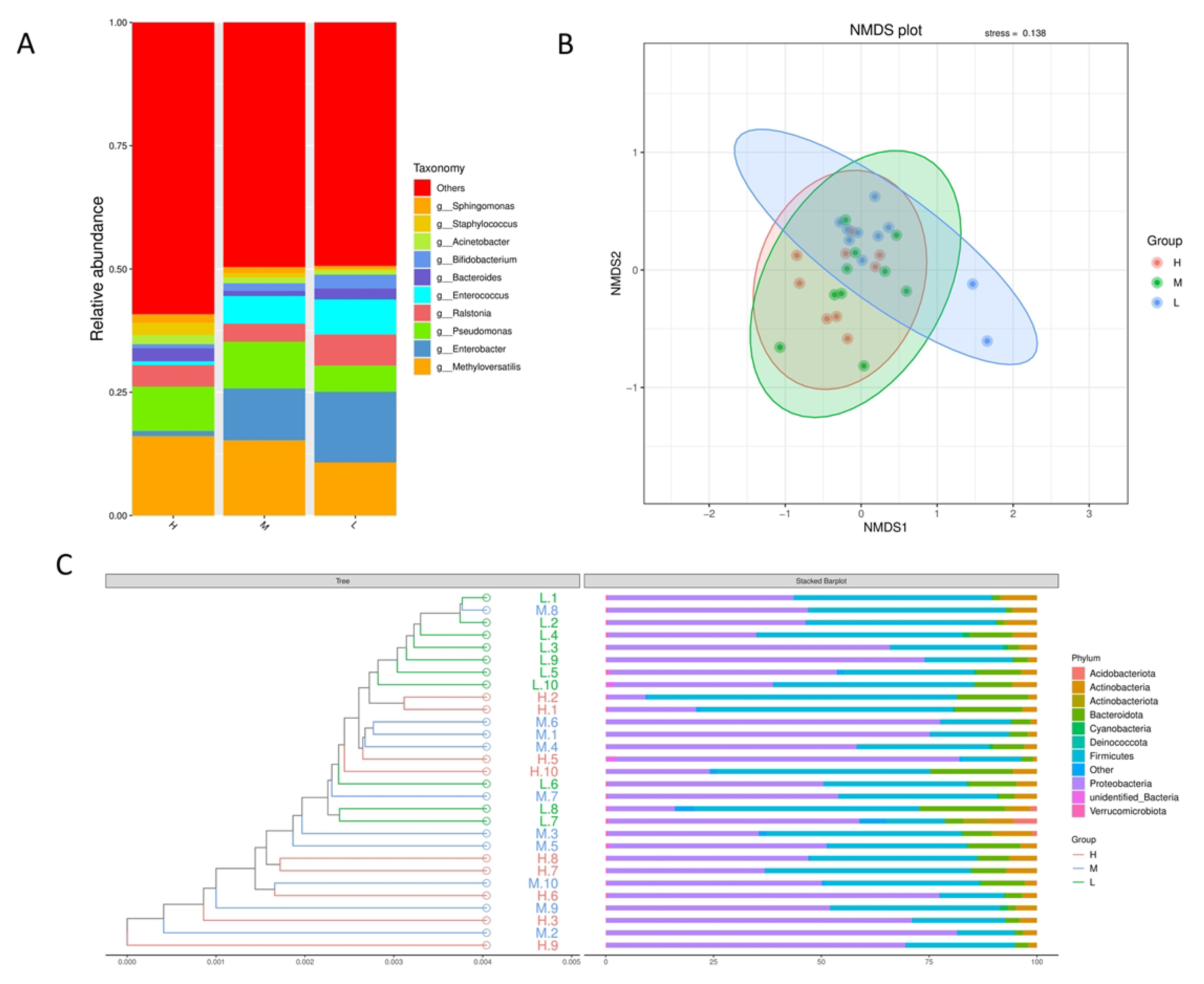
3.3. Effects of Different Somatic Cell Levels on the Diversity of Microbiota in Milk Samples
3.4. Effects of Different Somatic Cell Levels on the Composition of Microflora in Milk Samples
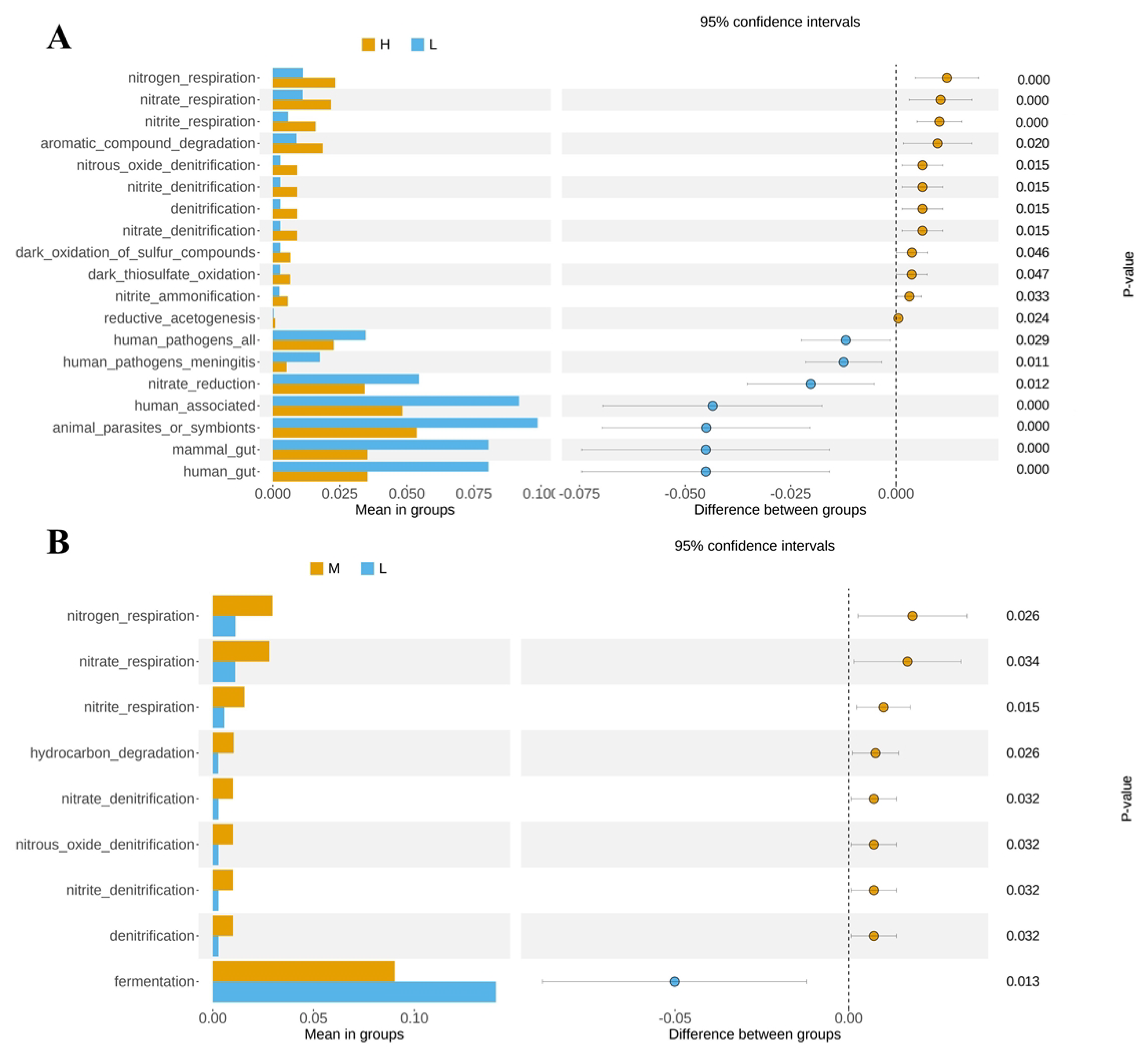
3.5. Functional Prediction of the Predominant Taxa
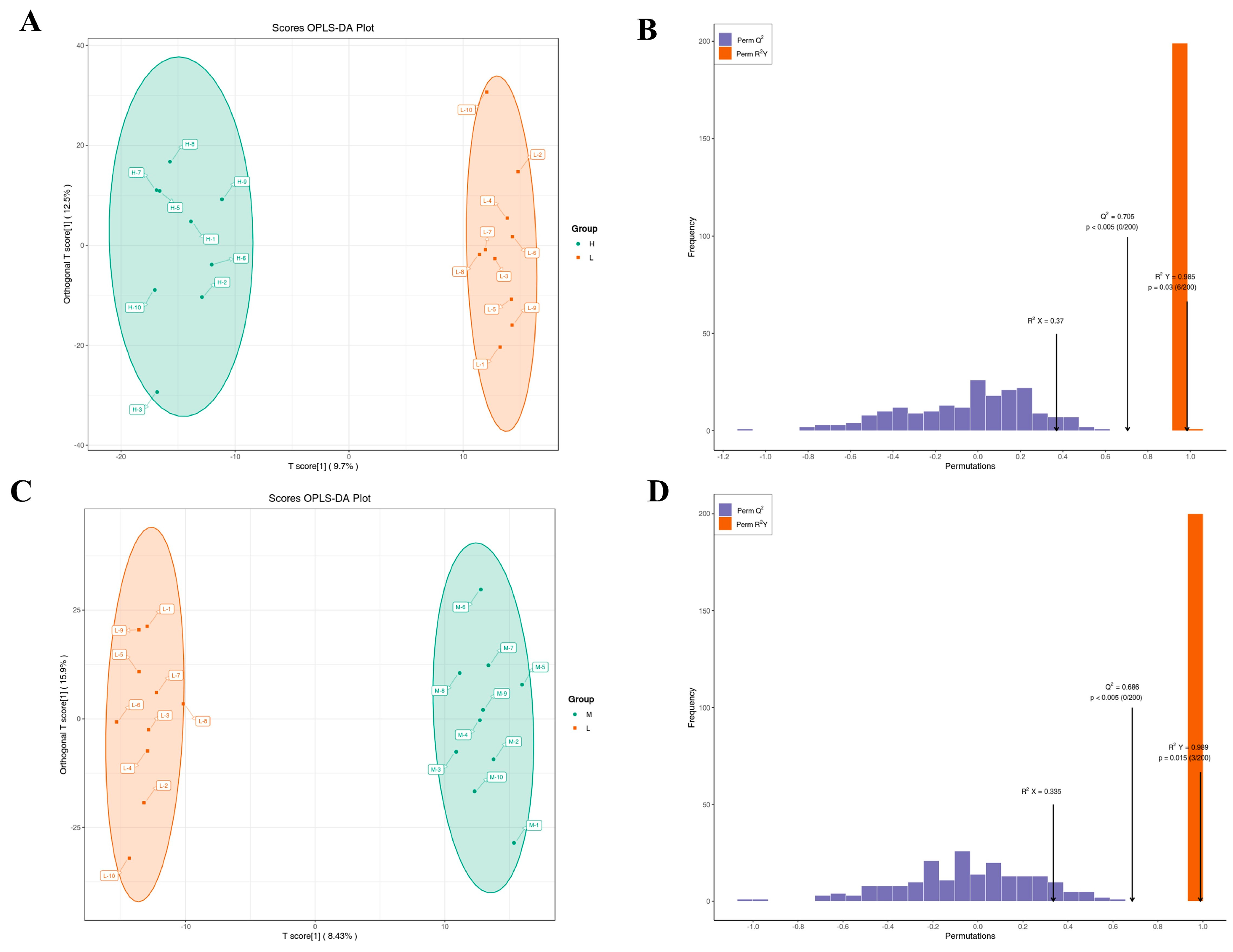
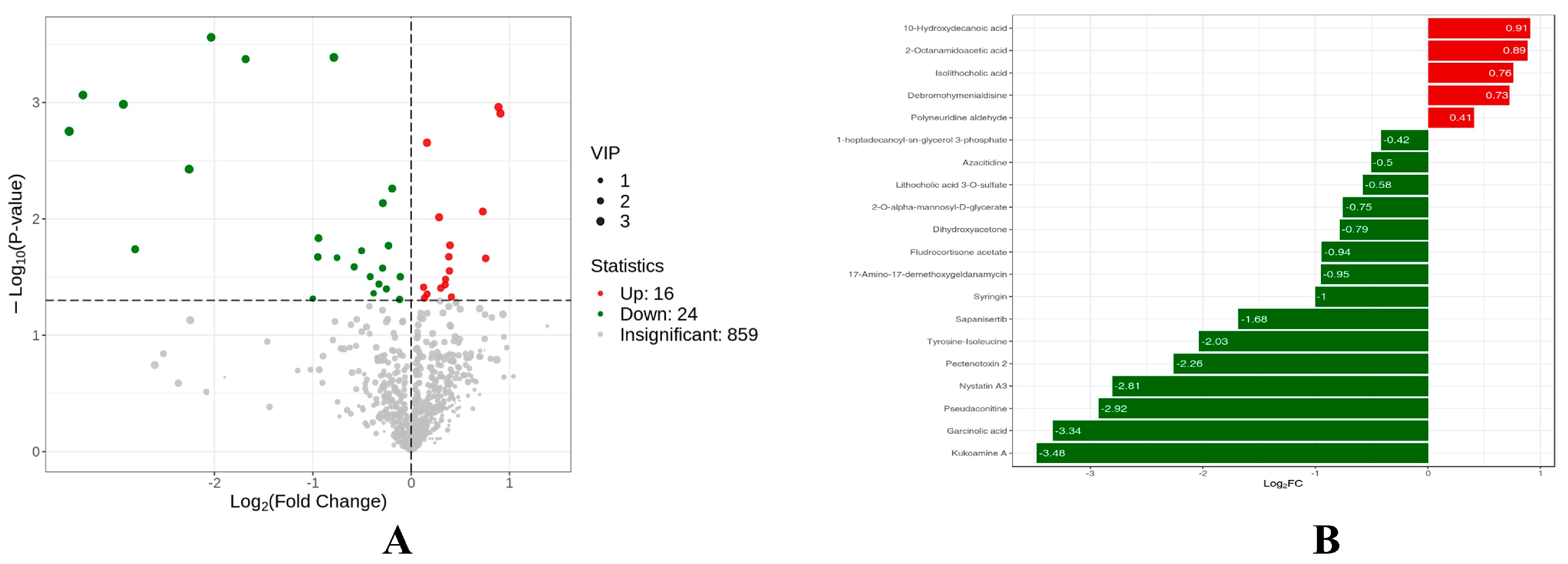
3.6. Metabolome Results
3.7. Correlation between Milk Microbiota and Metabolites at Different Somatic Cell Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leischner, C.; Egert, S.; Burkard, M.; Venturelli, S. Potential Protective Protein Components of Cow’s Milk against Certain Tumor Entities. Nutrients 2021, 13, 1974. [Google Scholar] [CrossRef] [PubMed]
- Stelwagen, K.; Carpenter, E.; Haigh, B.; Hodgkinson, A.; Wheeler, T.T. Immune components of bovine colostrum and milk. J. Anim. Sci. 2009, 87, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Bar, D.; Tauer, L.; Bennett, G.; González, R.N.; Hertl, J.; Schukken, Y.; Schulte, H.; Welcome, F.; Gröhn, Y. The Cost of Generic Clinical Mastitis in Dairy Cows as Estimated by Using Dynamic Programming. J. Dairy Sci. 2008, 91, 2205–2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Zhou, C.; Basang, W.; Zhu, Y.; Wang, X.; Li, C.; Chen, L.; Zhou, X. Mechanisms by which mastitis affects reproduction in dairy cow: A review. Reprod. Domest. Anim. 2021, 56, 1165–1175. [Google Scholar] [CrossRef]
- Ruegg, P.L. Management of mastitis on organic and conventional dairy farms. J. Anim. Sci. 2009, 87, 43–55. [Google Scholar] [CrossRef]
- Olechnowicz, J.; Jaškowski, J.M. Somatic Cells Count in Cow’s Bulk Tank Milk. J. Vet. Med. Sci. 2012, 74, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Jayarao, B.; Pillai, S.; Sawant, A.; Wolfgang, D.; Hegde, N. Guidelines for Monitoring Bulk Tank Milk Somatic Cell and Bacterial Counts. J. Dairy Sci. 2004, 87, 3561–3573. [Google Scholar] [CrossRef] [Green Version]
- Souto, L.I.; Minagawa, C.Y.; Telles, E.O.; Garbuglio, M.A.; Amaku, M.; Melville, P.A.; Dias, R.A.; Sakata, S.T.; Benites, N.R. Correlation between mastitis occurrence and the count of microorganisms in bulk raw milk of bovine dairy herds in four selective culture media. J. Dairy Res. 2009, 77, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Yatoo, M.I.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R.; et al. Advances in therapeutic and managemental approaches of bovine mastitis: A comprehensive review. Vet. Q. 2021, 41, 107–136. [Google Scholar] [CrossRef] [PubMed]
- Schukken, Y.H.; Wilson, D.J.; Welcome, F.; Garrison-Tikofsky, L.; Gonzalez, R.N. Monitoring udder health and milk quality using somatic cell counts. Vet. Res. 2003, 34, 579–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepers, A.; Lam, T.; Schukken, Y.; Wilmink, J.; Hanekamp, W. Estimation of Variance Components for Somatic Cell Counts to Determine Thresholds for Uninfected Quarters. J. Dairy Sci. 1997, 80, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- Sargeant, J.M.; Schukken, Y.H.; Leslie, K.E. Ontario Bulk Milk Somatic Cell Count Reduction Program: Progress and Outlook. J. Dairy Sci. 1998, 81, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Dohoo, I.R.; Smith, J.; Andersen, S.; Kelton, D.F.; Godden, S.; Mastitis Research Workers’ Conference. Diagnosing intramammary infections: Evaluation of definitions based on a single milk sample. J. Dairy Sci. 2011, 94, 250–261. [Google Scholar] [CrossRef]
- Timms, L.; Schultz, L. Dynamics and Significance of Coagulase-Negative Staphylococcal Intramammary Infections. J. Dairy Sci. 1987, 70, 2648–2657. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.; Barcelos, M.M.; Cue, R.I.; Anderson, K.L.; Dos Santos, M.V.; Gonçalves, J.L. Chronic subclinical mastitis reduces milk and components yield at the cow level. J. Dairy Res. 2020, 87, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Hand, K.; Godkin, A.; Kelton, D. Milk production and somatic cell counts: A cow-level analysis. J. Dairy Sci. 2012, 95, 1358–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagnestam-Nielsen, C.; Emanuelson, U.; Berglund, B.; Strandberg, E. Relationship between somatic cell count and milk yield in different stages of lactation. J. Dairy Sci. 2009, 92, 3124–3133. [Google Scholar] [CrossRef] [Green Version]
- McDougall, S.; Williamson, J.; Lacy-Hulbert, J. Bacteriological outcomes following random allocation to quarter-level selection based on California Mastitis Test score or cow-level allocation based on somatic cell count for dry cow therapy. J. Dairy Sci. 2022, 105, 2453–2472. [Google Scholar] [CrossRef]
- Xiao, H.-B.; Wang, C.-R.; Liu, Z.-K.; Wang, J.-Y. LPS induces pro-inflammatory response in mastitis mice and mammary epithelial cells: Possible involvement of NF-κB signaling and OPN. Pathol. Biol. 2015, 63, 11–16. [Google Scholar] [CrossRef]
- Hu, H.; Fang, Z.; Mu, T.; Wang, Z.; Ma, Y.; Ma, Y. Application of Metabolomics in Diagnosis of Cow Mastitis: A Review. Front. Vet. Sci. 2021, 8, 747519. [Google Scholar] [CrossRef]
- Wang, Y.; Nan, X.; Zhao, Y.; Wang, H.; Wang, M.; Jiang, L.; Zhang, F.; Xue, F.; Hua, D.; Li, K.; et al. Coupling 16S rDNA Sequencing and Untargeted Mass Spectrometry for Milk Microbial Composition and Metabolites from Dairy Cows with Clinical and Subclinical Mastitis. J. Agric. Food Chem. 2020, 68, 8496–8508. [Google Scholar] [CrossRef] [PubMed]
- Paape, M.; Tucker, H.; Hafs, H. Comparison of Methods for Estimating Milk Somatic Cells. J. Dairy Sci. 1965, 48, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.; Schultz, L.; Barr, G. Investigations on Possible Use of Mastitis-Screening Tests in Dairy Herd Improvement Association Central Laboratories. J. Dairy Sci. 1967, 50, 47–52. [Google Scholar] [CrossRef]
- Wittek, T.; Mader, C.; Ribitsch, V.; Burgstaller, J. Measurement of oxygen concentration for detection of subclinical mastitis. Schweiz. Arch. Tierheilkd. 2019, 161, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Nyman, A.-K.; Waller, K.P.; Bennedsgaard, T.; Larsen, T.; Emanuelson, U. Associations of udder-health indicators with cow factors and with intramammary infection in dairy cows. J. Dairy Sci. 2014, 97, 5459–5473. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.; García, M.B.M. A 100-Year Review: Advances in goat milk research. J. Dairy Sci. 2017, 100, 10026–10044. [Google Scholar] [CrossRef] [Green Version]
- Stocco, G.; Summer, A.; Cipolat-Gotet, C.; Zanini, L.; Vairani, D.; Dadousis, C.; Zecconi, A. Differential Somatic Cell Count as a Novel Indicator of Milk Quality in Dairy Cows. Animals 2020, 10, 753. [Google Scholar] [CrossRef]
- Ruegg, P.L. A 100-Year Review: Mastitis detection, management, and prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [Green Version]
- Plastridge, W. Bovine Mastitis: A Review. J. Dairy Sci. 1958, 41, 1141–1181. [Google Scholar] [CrossRef]
- Oliveira, L.; Hulland, C.; Ruegg, P. Characterization of clinical mastitis occurring in cows on 50 large dairy herds in Wisconsin. J. Dairy Sci. 2013, 96, 7538–7549. [Google Scholar] [CrossRef] [PubMed]
- Bradley, A.J.; Green, M.J. Aetiology of clinical mastitis in six Somerset dairy herds. Vet. Rec. 2001, 148, 683–686. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Ibeagha-Awemu, E.M.; Liang, G.; Beaudoin, F.; Zhao, X.; Guan, L.L. Transcriptome microRNA profiling of bovine mammary epithelial cells challenged with Escherichia coli or Staphylococcus aureus bacteria reveals pathogen directed microRNA expression profiles. BMC Genom. 2014, 15, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, P.-A.; Chien, S.-Y.; Chan, Y.-L.; Lu, M.-K.; Wu, C.-H.; Kong, Z.-L.; Wu, C.-J. Inhibition of Lipopolysaccharide (LPS)-Induced Inflammatory Responses by Sargassum hemiphyllum Sulfated Polysaccharide Extract in RAW 264.7 Macrophage Cells. J. Agric. Food Chem. 2011, 59, 2062–2068. [Google Scholar] [CrossRef] [PubMed]
- Califf, R.M. Biomarker definitions and their applications. Exp. Biol. Med. 2018, 243, 213–221. [Google Scholar] [CrossRef]
- Aronson, J.K.; Ferner, R.E. Biomarkers—A General Review. Curr. Protoc. Pharmacol. 2017, 76, 9–23. [Google Scholar] [CrossRef]
- Wisnieski, L.; Norby, B.; Pierce, S.J.; Becker, T.; Sordillo, L.M. Prospects for predictive modeling of transition cow diseases. Anim. Health Res. Rev. 2019, 20, 19–30. [Google Scholar] [CrossRef]
- Leblanc, S. Monitoring Metabolic Health of Dairy Cattle in the Transition Period. J. Reprod. Dev. 2010, 56, S29–S35. [Google Scholar] [CrossRef] [Green Version]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell. Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef]
- Miles, A.M.; Posbergh, C.J.; Huson, H.J. Direct Phenotyping and Principal Component Analysis of Type Traits Implicate Novel QTL in Bovine Mastitis through Genome-Wide Association. Animals 2021, 11, 1147. [Google Scholar] [CrossRef]
- Lai, Y.C.; Lai, Y.T.; Rahman, M.M.; Chen, H.W.; Husna, A.A.; Fujikawa, T.; Ando, T.; Kitahara, G.; Koiwa, M.; Kubota, C.; et al. Bovine milk transcriptome analysis reveals microRNAs and RNU2 involved in mastitis. FEBS J. 2019, 287, 1899–1918. [Google Scholar] [CrossRef]
- Bathla, S.; Sindhu, A.; Kumar, S.; Dubey, S.K.; Pattnaik, S.; Rawat, P.; Chopra, A.; Dang, A.; Kaushik, J.K.; Mohanty, A.K. Tandem Mass Tag (TMT)-based quantitative proteomics reveals potential targets associated with onset of Sub-clinical Mastitis in cows. Sci. Rep. 2020, 10, 9321. [Google Scholar] [CrossRef] [PubMed]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies—Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [Green Version]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taponen, S.; Salmikivi, L.; Simojoki, H.; Koskinen, M.; Pyörälä, S. Real-time polymerase chain reaction-based identification of bacteria in milk samples from bovine clinical mastitis with no growth in conventional culturing. J. Dairy Sci. 2009, 92, 2610–2617. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.H.; Vaidya, Y.H.; Patel, R.J.; Pandit, R.J.; Joshi, C.G.; Kunjadiya, A.P. Culture independent assessment of human milk microbial community in lactational mastitis. Sci. Rep. 2017, 7, 7804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idriss, S.E.A.B.; Tančin, V.; Foltys, V.; Kirchnerová, K.; Tančinová, D.; Vršková, M. Relationship between mastitis causative pathogens and somatic cell counts in dairy cows. Potravin. Slovak J. Food Sci. 2013, 7, 207–212. [Google Scholar] [CrossRef]
- Sundekilde, U.; Poulsen, N.; Larsen, L.; Bertram, H. Nuclear magnetic resonance metabonomics reveals strong association between milk metabolites and somatic cell count in bovine milk. J. Dairy Sci. 2013, 96, 290–299. [Google Scholar] [CrossRef] [Green Version]
- Huszenicza, G.; Jánosi, S.; Gáspárdy, A.; Kulcsár, M. Endocrine aspects in pathogenesis of mastitis in postpartum dairy cows. Anim. Reprod. Sci. 2004, 82–83, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Abril, A.G.; Gonzalez-Villa, T.; Barros-Velázquez, J.; Cañas, B.; Sánchez-Pérez, A.; Calo-Mata, P.; Carrera, M. Staphylococcus aureus Exotoxins and Their Detection in the Dairy Industry and Mastitis. Toxins 2020, 12, 537. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Xue, M.; Liu, J. Composition of Rumen Bacterial Community in Dairy Cows with Different Levels of Somatic Cell Counts. Front. Microbiol. 2018, 9, 3217. [Google Scholar] [CrossRef] [Green Version]
- Davin-Regli, A.; Lavigne, J.-P.; Pagès, J.-M. Enterobacter spp.: Update on Taxonomy, Clinical Aspects, and Emerging Antimicrobial Resistance. Clin. Microbiol. Rev. 2019, 32, e00002-19. [Google Scholar] [CrossRef]
- Qiao, J.; Kwok, L.; Zhang, J.; Gao, P.; Zheng, Y.; Guo, Z.; Hou, Q.; Huo, D.; Huang, W.; Zhang, H. Reduction of Lactobacillus in the milks of cows with subclinical mastitis. Benef. Microbes 2015, 6, 485–490. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Mining Lactobacillus and Bifidobacterium for organisms with long-term gut colonization potential. Clin. Nutr. 2020, 39, 1315–1323. [Google Scholar] [CrossRef] [PubMed]
- Petschow, B.W.; Talbott, R.D. Response of Bifidobacterium Species to Growth Promoters in Human and Cow Milk. Pediatr. Res. 1991, 29, 208–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, N.; Guidot, A.; Vailleau, F.; Valls, M. Ralstonia solanacearum, a widespread bacterial plant pathogen in the post-genomic era. Mol. Plant Pathol. 2013, 14, 651–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Liu, M.; Feng, L.; Wang, Z. Comparative genomic analysis reveals the evolution and environmental adaptation of Acinetobacter johnsonii. Gene 2021, 808, 145985. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Guan, Y.; Li, X.; Fan, X.; Zhu, Z.; Xing, H.; Wang, Z. Phenotype profiles and adaptive preference of Acinetobacter johnsonii isolated from Ba River with different environmental backgrounds. Environ. Res. 2021, 196, 110913. [Google Scholar] [CrossRef]
- Ma, T.; Shen, L.; Wen, Q.; Lv, R.; Hou, Q.; Kwok, L.Y.; Sun, Z.; Zhang, H. PacBio sequencing revealed variation in the microbiota diversity, species richness and composition between milk collected from healthy and mastitis cows. Microbiology 2021, 167, e000968. [Google Scholar] [CrossRef]
- Berardo, N.; Giraudo, J.; Magnano, G.; Nader-Macias, M.E.F.; Bogni, C.; Pellegrino, M. Lactococcus lactis subsp. lactis CRL1655 and Schleiferilactobacillus perolens CRL1724 inhibit the adherence of common bovine mastitis pathogens to mammary gland cells, without causing histological changes in the mammary gland. J. Appl. Microbiol. 2022, 133, 733–742. [Google Scholar] [CrossRef]
- Mielko, K.A.; Jabłoński, S.J.; Milczewska, J.; Sands, D.; Łukaszewicz, M.; Młynarz, P. Metabolomic studies of Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2019, 35, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves Pessoa, R.B.; de Oliveira, W.F.; Marques, D.S.C.; dos Santos Correia, M.T.; de Carvalho, E.V.M.M.; Coelho, L.C.B.B. The genus Aeromonas: A general approach. Microb. Pathog. 2019, 130, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jin, S.; Feng, C.; Song, H.; Raza, S.H.A.; Yu, H.; Zhang, L.; Chi, T.; Qi, Y.; Zhang, D.; et al. Aeromonas veronii virulence and adhesion attenuation mediated by the gene aodp. J. Fish Dis. 2021, 45, 231–247. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [Green Version]
- Duskova, D.; Marounek, M. Fermentation of pectin and glucose, and activity of pectin-degrading enzymes in the rumen bacterium Lachnospira multiparus. Lett. Appl. Microbiol. 2001, 33, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia muciniphila: Paradigm for next-generation beneficial microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila is a promising probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gryaznova, M.V.; Syromyatnikov, M.Y.; Dvoretskaya, Y.D.; Solodskikh, S.A.; Klimov, N.T.; Mikhalev, V.I.; Zimnikov, V.I.; Mikhaylov, E.V.; Popov, V.N. Microbiota of Cow’s Milk with Udder Pathologies. Microorganisms 2021, 9, 1974. [Google Scholar] [CrossRef]
- Liu, G.; Liu, Y.; Ali, T.; Ferreri, M.; Gao, J.; Chen, W.; Yin, J.; Su, J.; Fanning, S.; Han, B. Molecular and Phenotypic Characterization of Aerococcus viridans Associated with Subclinical Bovine Mastitis. PLoS ONE 2015, 10, e0125001. [Google Scholar] [CrossRef] [Green Version]
- Grispoldi, L.; Karama, M.; Ianni, F.; La Mantia, A.; Pucciarini, L.; Camaioni, E.; Sardella, R.; Sechi, P.; Natalini, B.; Cenci-Goga, B.T. The Relationship between S. aureus and Branched-Chain Amino Acids Content in Composite Cow Milk. Animals 2019, 9, 981. [Google Scholar] [CrossRef] [Green Version]
- Mehta, R.; Sonavane, M.; Migaud, M.E.; Gassman, N.R. Exogenous exposure to dihydroxyacetone mimics high fructose induced oxidative stress and mitochondrial dysfunction. Environ. Mol. Mutagen. 2021, 62, 185–202. [Google Scholar] [CrossRef]
- Mori, M.-A.; Matsuhashi, T.; Miyahara, T.; Shibata, S.; Īzima, C.; Kozuka, H. Reduction of 2,4-dinitrotoluene by Wistar rat liver microsomal and cytosol fractions. Toxicol. Appl. Pharmacol. 1984, 76, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Mi, J.; Denman, S.E.; Basangwangdui; Pingcuozhandui; Zhang, Q.; Long, R.; McSweeney, C.S. Changes in rumen microbial community composition in yak in response to seasonal variations. J. Appl. Microbiol. 2021, 132, 1652–1665. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, H.; Lobley, G.E.; Doepel, L.; Raggio, G.; Rulquin, H.; Lemosquet, S. Triennial Lactation Symposium: Mammary metabolism of amino acids in dairy cows. J. Anim. Sci. 2012, 90, 1708–1721. [Google Scholar] [CrossRef]
- Hussain, A.; Hassan, Q.P.; Shouche, Y.S. New approaches for antituberculosis leads from Actinobacteria. Drug Discov. Today 2020, 25, 2335–2342. [Google Scholar] [CrossRef] [PubMed]
- Kuhla, B.; Kaever, V.; Tuchscherer, A.; Kuhla, A. Involvement of Plasma Endocannabinoids and the Hypothalamic Endocannabinoid System in Increasing Feed Intake after Parturition of Dairy Cows. Neuroendocrinology 2019, 110, 246–257. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Fat (%) | Pro (%) | Lact (%) | Urea (%) | SCC (Cells/mL) |
|---|---|---|---|---|---|
| H | 3.51 | 3.37 | 5.23 | 9.41 | 587,000 |
| H | 5.81 | 4.30 | 4.91 | 6.2 | 750,000 |
| H | 4.47 | 3.42 | 4.56 | 16.76 | 551,000 |
| H | 4.69 | 3.38 | 4.88 | 13.72 | 2,612,000 |
| H | 5.35 | 3.42 | 5.13 | 12.3 | 2,794,000 |
| H | 5.36 | 3.20 | 5.1 | 12.47 | 568,000 |
| H | 4.29 | 3.07 | 5.35 | 11.02 | 765,000 |
| H | 5.35 | 3.40 | 4.97 | 9.74 | 889,000 |
| H | 4.50 | 3.51 | 5.16 | 12.2 | 659,000 |
| H | 5.47 | 3.53 | 5.34 | 13.3 | 3,546,000 |
| M | 6.37 | 3.29 | 5.4 | 9.56 | 262,000 |
| M | 5.8 | 3.24 | 5.32 | 11.88 | 339,000 |
| M | 5.09 | 3.67 | 4.95 | 13.91 | 329,000 |
| M | 4.78 | 3.50 | 5.02 | 14.87 | 337,000 |
| M | 4.09 | 3.24 | 5.14 | 11.14 | 217,000 |
| M | 4.87 | 3.25 | 4.96 | 12.02 | 316,000 |
| M | 4.91 | 3.55 | 5.06 | 11.75 | 226,000 |
| M | 4.88 | 3.23 | 5.11 | 12.3 | 299,000 |
| M | 5.42 | 3.59 | 4.87 | 14.36 | 221,000 |
| M | 4.82 | 3.35 | 5.18 | 12.76 | 312,000 |
| L | 5.00 | 3.48 | 5.33 | 13.46 | 55,000 |
| L | 5.64 | 3.70 | 5.05 | 14.44 | 31,000 |
| L | 3.40 | 3.10 | 5.33 | 13.46 | 51,000 |
| L | 5.12 | 3.17 | 5.38 | 13.52 | 40,000 |
| L | 5.31 | 3.39 | 5.15 | 6.69 | 103,000 |
| L | 5.12 | 3.31 | 5.34 | 16.19 | 65,000 |
| L | 5.19 | 3.18 | 5 | 14.33 | 114,000 |
| L | 4.26 | 3.40 | 5.33 | 18.89 | 66,000 |
| L | 4.66 | 3.18 | 5.23 | 10.33 | 103,000 |
| L | 4.90 | 3.25 | 5.4 | 9.85 | 52,000 |
| Group | Raw Tags | Clean Tags | Effective Tags | Mean Length | GC (%) |
|---|---|---|---|---|---|
| H | 763,556 | 634,829 | 568,851 | 409 | 53.21 |
| M | 786,328 | 700,067 | 616,384 | 417 | 53.61 |
| L | 731,107 | 712,575 | 615,691 | 420 | 54.10 |
| Total | 2,280,991 | 2,047,471 | 1,800,926 | 415 | 53.66 |
| Group | Shannon | Simpson | Chao1 | ACE |
|---|---|---|---|---|
| H | 6.53 ± 1.75 | 0.91 ± 0.09 | 1475.80 ± 383.74 | 1486.88 ± 380.89 |
| M | 5.97 ± 1.44 | 0.88 ± 0.08 | 1566.86 ± 492.09 | 1561.38 ± 499.96 |
| L | 6.10 ± 1.45 | 0.88 ± 0.09 | 1721.27 ± 432.20 | 1722.79 ± 441.51 |
| Index | Taxonomy | Correlation | p-Value | Compounds |
|---|---|---|---|---|
| MW0016991 | p__Actinobacteriota | −0.7509 | 0.0002 | Cer(d18:0/23:0) |
| MW0016991 | p__Verrucomicrobiota | −0.7509 | 0.0002 | Cer(d18:0/23:0) |
| MW0016070 | p__Actinobacteriota | −0.7193 | 0.0005 | benzoyl-coenzyme A |
| MW0016070 | p__Verrucomicrobiota | −0.7070 | 0.0007 | benzoyl-coenzyme A |
| MW0155767 | p__Actinobacteriota | −0.7632 | 0.0001 | Pro-Ala-Leu |
| MW0150025 | p__Actinobacteriota | −0.7035 | 0.0008 | Glu-His-His |
| MEDN0061 | p__Verrucomicrobiota | 0.8088 | 0.0000 | Phenylacetylglycine |
| MW0007503 | p__Verrucomicrobiota | 0.8000 | 0.0000 | Lidocaine |
| MW0011802 | p__Verrucomicrobiota | 0.7088 | 0.0007 | prostaglandin E2 1-glyceryl ester |
| MW0157826 | p__Actinobacteriota | 0.7789 | 0.0001 | Thr-Trp-Met |
| MW0157826 | p__Verrucomicrobiota | 0.7474 | 0.0002 | Thr-Trp-Met |
| MW0105257 | p__Verrucomicrobiota | 0.7579 | 0.0002 | Ketoleucine |
| MW0110625 | p__Verrucomicrobiota | 0.7123 | 0.0006 | 1-O-Hexadecyl-2-deoxy-2-thio-S-acetyl-sn-glyceryl-3-phosphorylcholine |
| MW0054970 | p__Actinobacteriota | 0.7684 | 0.0001 | MG(0:0/18:2(9Z,12Z)/0:0) |
| MW0012825 | p__Actinobacteriota | −0.7000 | 0.0008 | 1-O-Hexadecyl-sn-glycero-3-phosphocholine |
| MW0012825 | p__Verrucomicrobiota | −0.7246 | 0.0005 | 1-O-Hexadecyl-sn-glycero-3-phosphocholine |
| MEDN0311 | p__Actinobacteriota | 0.7228 | 0.0005 | Dodecanedioic Aicd |
| MW0055024 | p__Actinobacteriota | 0.7000 | 0.0008 | MG(18:2/0:0/0:0) |
| MW0062272 | p__Verrucomicrobiota | 0.7509 | 0.0002 | Prostaglandin h2 |
| MW0012772 | p__Actinobacteriota | 0.7491 | 0.0002 | Glyceryl arachidonate |
| MEDN0658 | p__Actinobacteriota | 0.7193 | 0.0005 | Hexadecanedioic acid |
| MW0054060 | p__Verrucomicrobiota | 0.7018 | 0.0008 | Isobutyryl CoA |
| Index | Taxonomy | Correlation | p-Value | Compounds |
|---|---|---|---|---|
| MW0155873 | p__Deferribacteres | −0.7262 | 0.0003 | Pro-Gly-Val |
| MW0159163 | Others | 0.7910 | 0.0000 | Val-Ser-Ser-Ser-Leu |
| MW0108785 | p__Deferribacteres | −0.7490 | 0.0001 | NG-Amino-l-arginine |
| MW0121539 | p__Deferribacteres | −0.7323 | 0.0002 | 5-Hydroxyisourate |
| MW0112789 | Others | 0.7383 | 0.0002 | 3-Dehydrosphinganine |
| MW0056887 | Others | −0.7880 | 0.0000 | 1-Palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine |
| MW0137085 | p__Deferribacteres | −0.7132 | 0.0004 | 7-Hydroxywarfarin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
She, Y.; Liu, J.; Su, M.; Li, Y.; Guo, Y.; Liu, G.; Deng, M.; Qin, H.; Sun, B.; Guo, J.; et al. A Study on Differential Biomarkers in the Milk of Holstein Cows with Different Somatic Cells Count Levels. Animals 2023, 13, 2446. https://doi.org/10.3390/ani13152446
She Y, Liu J, Su M, Li Y, Guo Y, Liu G, Deng M, Qin H, Sun B, Guo J, et al. A Study on Differential Biomarkers in the Milk of Holstein Cows with Different Somatic Cells Count Levels. Animals. 2023; 13(15):2446. https://doi.org/10.3390/ani13152446
Chicago/Turabian StyleShe, Yuanhang, Jianying Liu, Minqiang Su, Yaokun Li, Yongqing Guo, Guangbin Liu, Ming Deng, Hongxian Qin, Baoli Sun, Jianchao Guo, and et al. 2023. "A Study on Differential Biomarkers in the Milk of Holstein Cows with Different Somatic Cells Count Levels" Animals 13, no. 15: 2446. https://doi.org/10.3390/ani13152446