
Alginate Nanoencapsulated Synbiotic Composite of Pomegranate Peel Phytogenics and Multi-Probiotic Species as a Potential Feed Additive: Physicochemical, Antioxidant, and Antimicrobial Activities

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fabrication and Characterization of Synbiotics
2.1.1. Prebiotic Preparation and Identification of Phenolic Compounds
2.1.2. Probiotic Strains
2.1.3. Fabrication of Alginate-CaCl2-Based Nanosynbiotic and Free Synbiotics
2.2. Synbiotic Characteristics
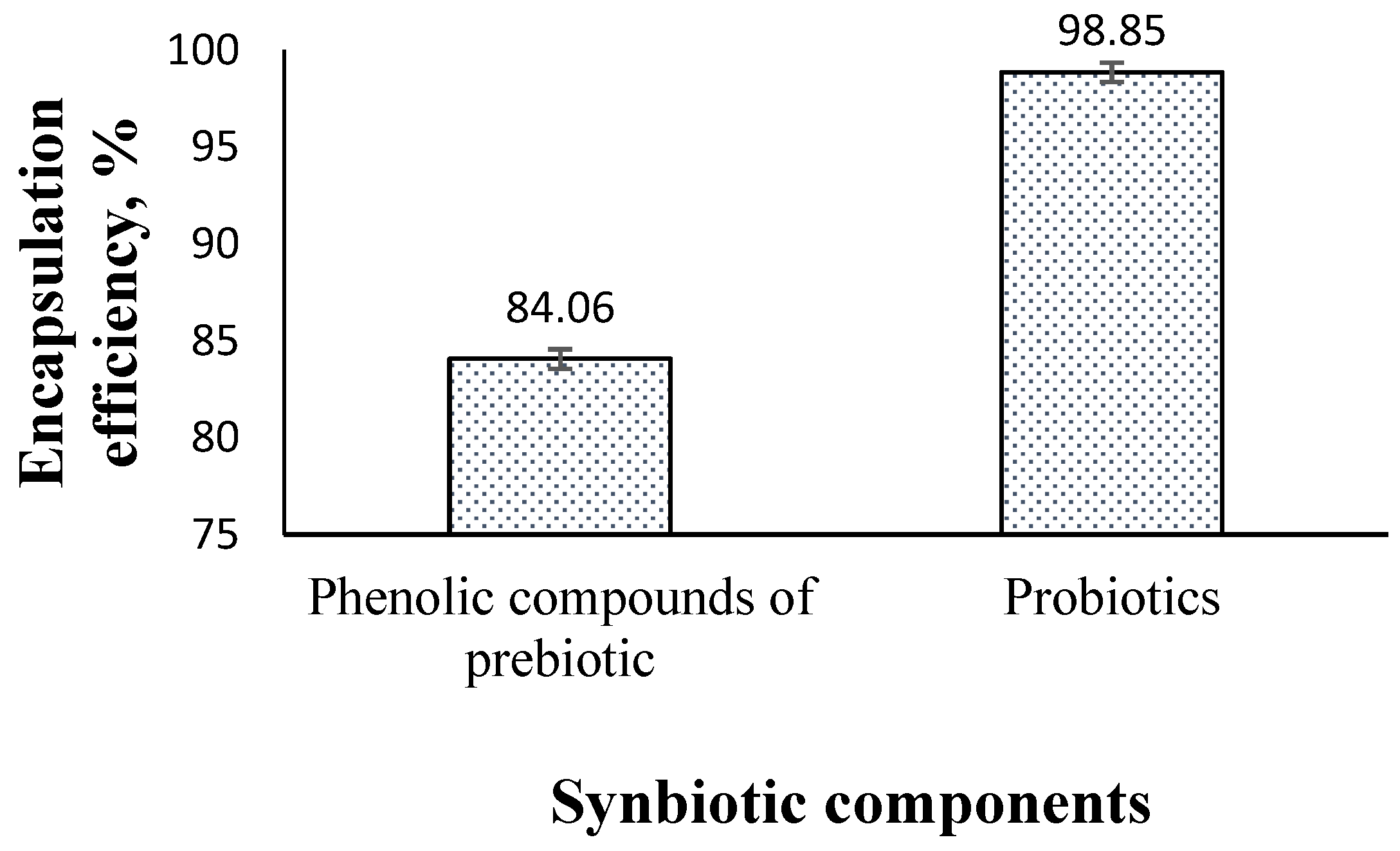
2.2.1. Encapsulation Efficiency
2.2.2. Morphology of Nanoencapsulated Synbiotics
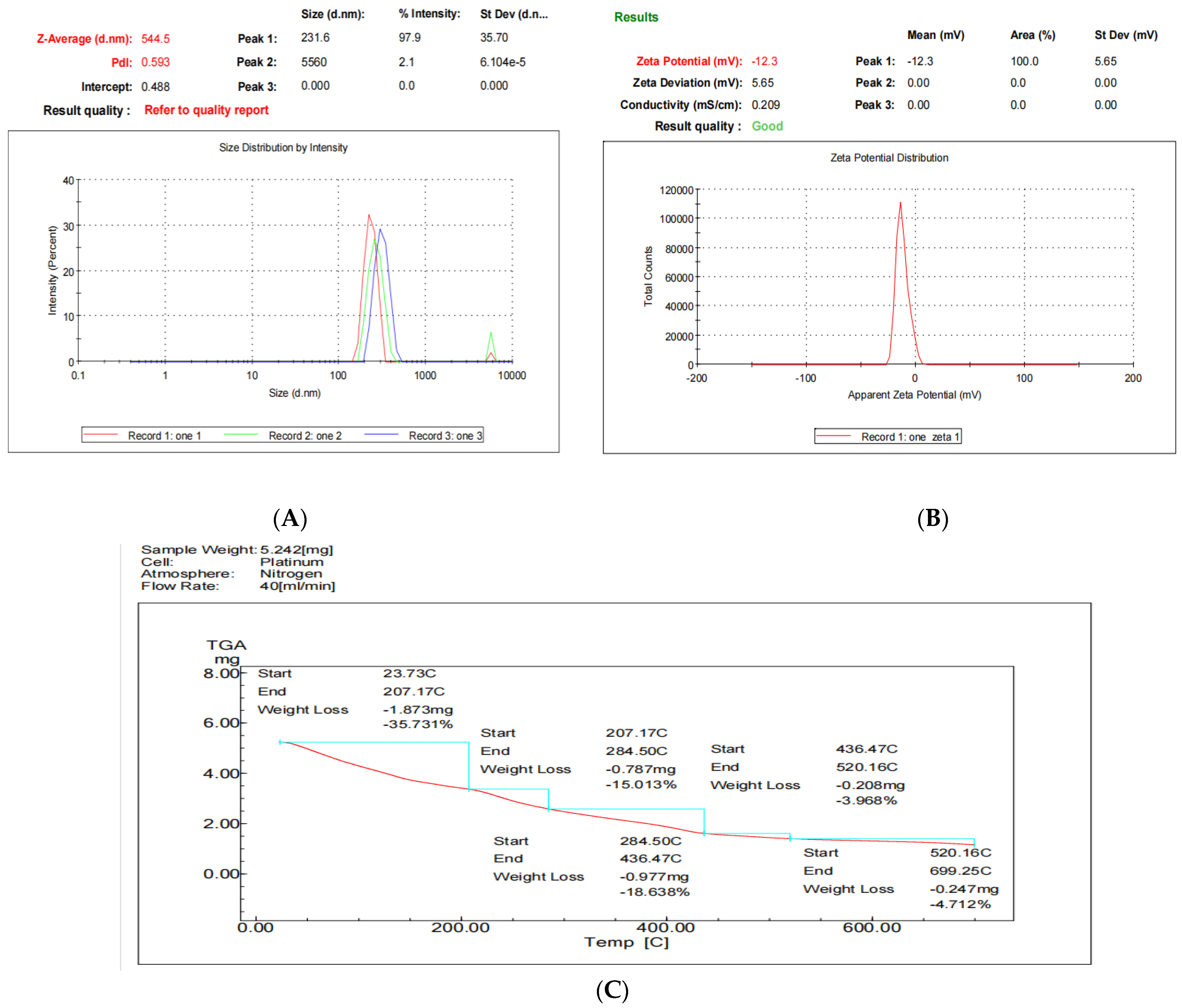
2.2.3. Size Distribution and Surface Charge
2.2.4. Thermogravimetric Analysis (TGA Analysis)
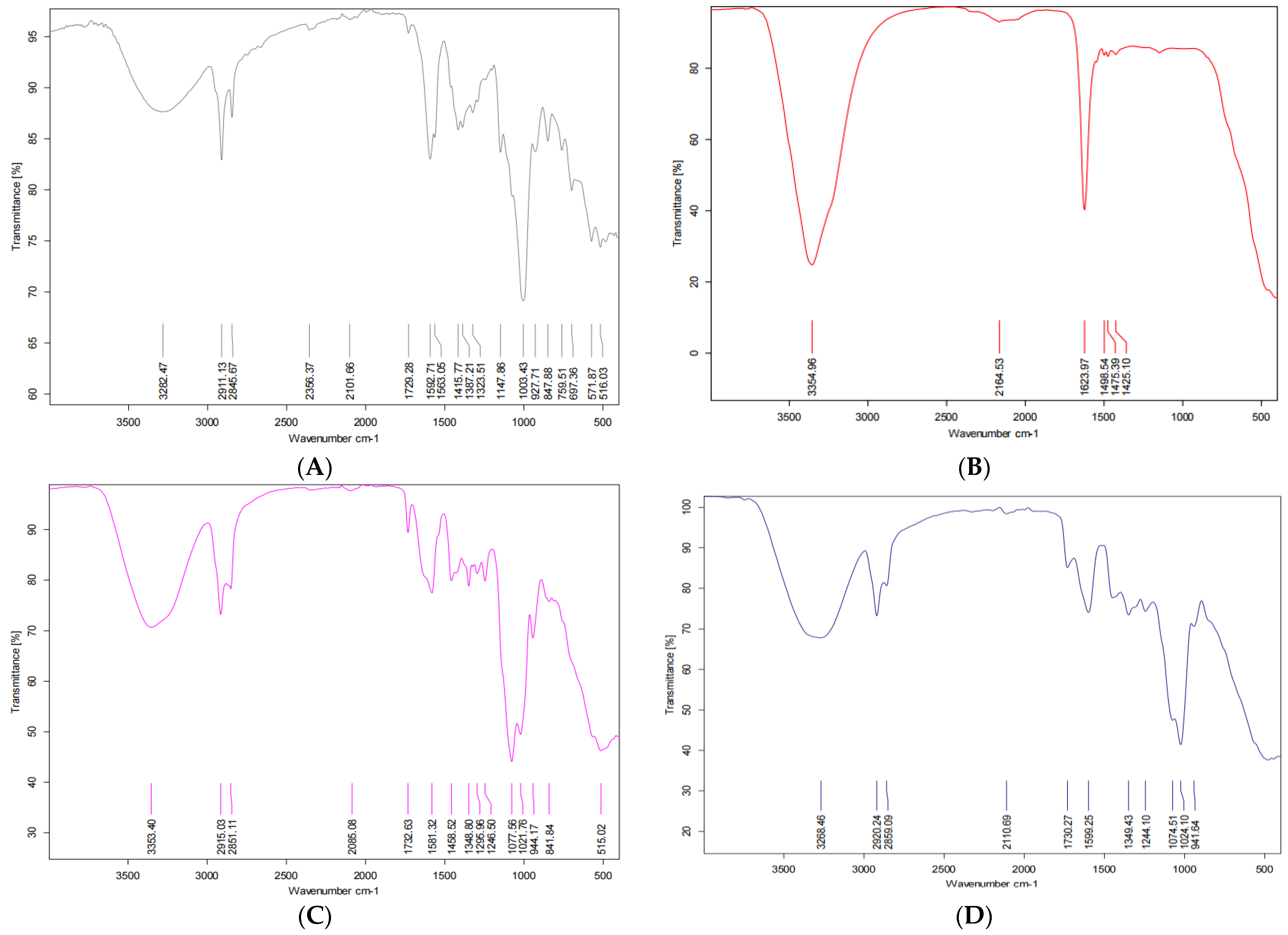
2.2.5. Fourier Transform Infrared Analysis
2.3. Gastric Tolerance and Storage Survivability of Probiotics
2.4. Antioxidant Activity of the Synbiotics (DPPH Scavenging Activity)
2.5. Antimicrobial Activity of the Synbiotics
2.6. Statistical Analyses
3. Results
3.1. Phenolic Compound of Prebiotic
3.2. The Physicochemical Characteristics of Nanoencapsulated Synbiotic
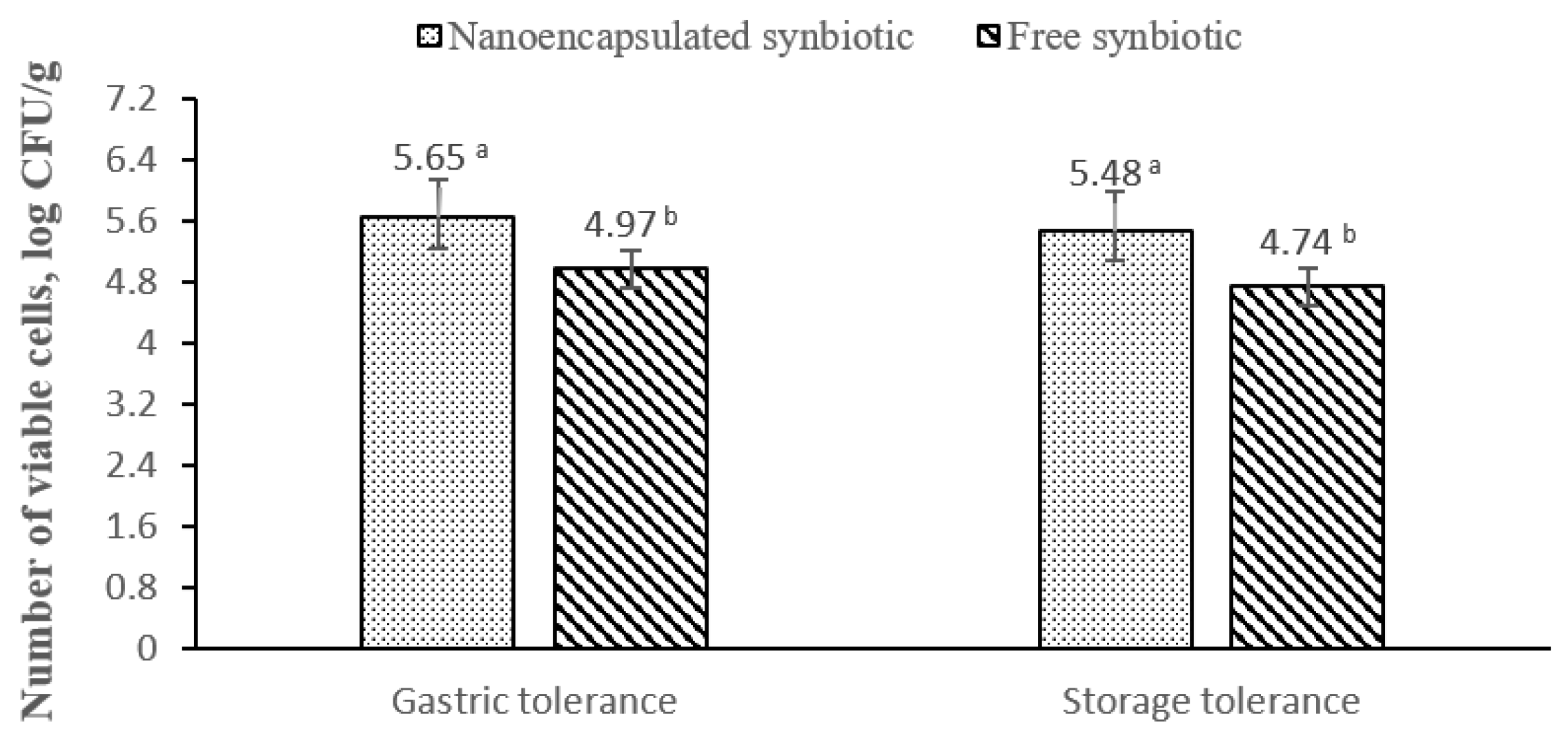
3.3. Gastric Tolerance and Storage Survivability of Probiotics
3.4. Antioxidant Activity of the Synbiotics
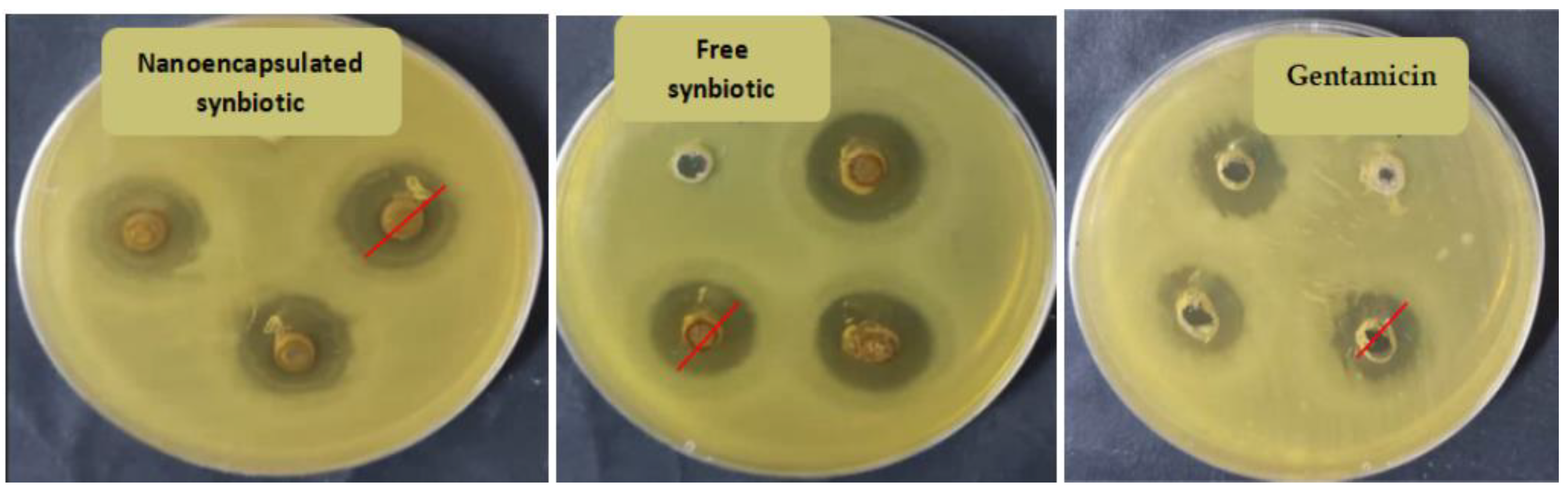
3.5. Antimicrobial Activity of the Synbiotics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hashem, N.; Soltan, Y.; El-Desoky, N.; Morsy, A.; Sallam, S. Effects of Moringa oleifera extracts and monensin on performance of growing rabbits. Livest. Sci. 2019, 228, 136–143. [Google Scholar] [CrossRef]
- Hashem, N.M.; Hosny, N.S.; El-Desoky, N.I.; Shehata, M.G. Effect of nanoencapsulated alginate-synbiotic on gut microflora balance, immunity, and growth performance of growing rabbits. Polymers 2021, 13, 4191. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, G. Synbiotic encapsulation of probiotic Latobacillus plantarum by alginate-arabinoxylan composite microspheres. Lwt 2018, 93, 135–141. [Google Scholar] [CrossRef]
- Das, T.K.; Pradhan, S.; Chakrabarti, S.; Mondal, K.C.; Ghosh, K. Current status of probiotic and related health benefits. Appl. Food Res. 2022, 2, 100185. [Google Scholar] [CrossRef]
- Hashem, N.M.; Gonzalez-Bulnes, A. The use of probiotics for management and improvement of reproductive eubiosis and function. Nutrients 2022, 14, 902. [Google Scholar] [CrossRef]
- Thanjavur, N.; Sangubotla, R.; Lakshmi, B.A.; Rayi, R.; Mekala, C.D.; Reddy, A.S.; Viswanath, B. Evaluating the antimicrobial and apoptogenic properties of bacteriocin (nisin) produced by Lactococcus lactis. Process Biochem. 2022, 122, 76–86. [Google Scholar] [CrossRef]
- Melo-Bolívar, J.F.; Ruiz Pardo, R.Y.; Hume, M.E.; Villamil Diaz, L.M. Multistrain probiotics use in main commercially cultured freshwater fish: A systematic review of evidence. Rev. Aquac. 2021, 13, 1758–1780. [Google Scholar] [CrossRef]
- Puvanasundram, P.; Chong, C.M.; Sabri, S.; Yusoff, M.S.; Karim, M. Multi-strain probiotics: Functions, effectiveness and formulations for aquaculture applications. Aquac. Rep. 2021, 21, 100905. [Google Scholar] [CrossRef]
- Chapman, C.; Gibson, G.R.; Rowland, I. Health benefits of probiotics: Are mixtures more effective than single strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, types, sources, mechanisms, and clinical applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- Tzounis, X.; Rodriguez-Mateos, A.; Vulevic, J.; Gibson, G.R.; Kwik-Uribe, C.; Spencer, J.P. Prebiotic evaluation of cocoa-derived flavanols in healthy humans by using a randomized, controlled, double-blind, crossover intervention study. Am. J. Clin. Nutr. 2011, 93, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Andishmand, H.; Azadmard-Damirchi, S.; Hamishekar, H.; Torbati, M.; Kharazmi, M.S.; Savage, G.P.; Tan, C.; Jafari, S.M. Nano-delivery systems for encapsulation of phenolic compounds from pomegranate peel. Adv. Colloid Interface Sci. 2023, 311, 102833. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimnezhad, P.; Khavarpour, M.; Khalili, S. Survival of Lactobacillus acidophilus as probiotic bacteria using chitosan nanoparticles. Int. J. Eng. 2017, 30, 456–463. [Google Scholar]
- El-Desoky, N.I.; Hashem, N.M.; Elkomy, A.G.; Abo-Elezz, Z.R. Improving rabbit doe metabolism and whole reproductive cycle outcomes via fatty acid-rich moringa oleifera leaf extract supplementation in free and nano-encapsulated forms. Animals 2022, 12, 764. [Google Scholar] [CrossRef] [PubMed]
- El-Desoky, N.I.; Hashem, N.M.; Gonzalez-Bulnes, A.; Elkomy, A.G.; Abo-Elezz, Z.R. Effects of a nanoencapsulated moringa leaf ethanolic extract on the physiology, metabolism and reproductive performance of rabbit does during summer. Antioxidants 2021, 10, 1326. [Google Scholar] [CrossRef]
- Lin, Y.-L.; Juan, I.-M.; Chen, Y.-L.; Liang, Y.-C.; Lin, J.-K. Composition of polyphenols in fresh tea leaves and associations of their oxygen-radical-absorbing capacity with antiproliferative actions in fibroblast cells. J. Agric. Food Chem. 1996, 44, 1387–1394. [Google Scholar] [CrossRef]
- Workie, M.; Kebede, B.; Tadesse, T.; Yimer, D.; Tibebu, T.; Abera, S.; Tilahun, A.; Alemu, M.; Daba, T.; Eshetu, A. In Vitro Evaluation of the Probiotic Potential of Lactic Acid Bacterial Strains Retrieved from Raw and Traditionally Fermented Cow Milk. East Afr. J. Sci. 2022, 16, 199–212. [Google Scholar]
- Hashem, N.M.; Shehata, M.G. Antioxidant and antimicrobial activity of Cleome droserifolia (Forssk.) Del. and its biological effects on redox status, immunity, and gut microflora. Animals 2021, 11, 1929. [Google Scholar] [CrossRef]
- Duman, D.; Karadag, A. Inulin added electrospun composite nanofibres by electrospinning for the encapsulation of probiotics: Characterisation and assessment of viability during storage and simulated gastrointestinal digestion. Int. J. Food Sci. Technol. 2021, 56, 927–935. [Google Scholar] [CrossRef]
- Atraki, R.; Azizkhani, M. Survival of probiotic bacteria nanoencapsulated within biopolymers in a simulated gastrointestinal model. Innov. Food Sci. Emerg. Technol. 2021, 72, 102750. [Google Scholar] [CrossRef]
- Dean, S.N.; Leary, D.H.; Sullivan, C.J.; Oh, E.; Walper, S.A. Isolation and characterization of Lactobacillus-derived membrane vesicles. Sci. Rep. 2019, 9, 877. [Google Scholar] [CrossRef] [Green Version]
- Hanafy, N.A.; Leporatti, S.; El-Kemary, M.A. Mucoadhesive hydrogel nanoparticles as smart biomedical drug delivery system. Appl. Sci. 2019, 9, 825. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Guillén, M.C.; Montero, M.P. Enhancement of oral bioavailability of natural compounds and probiotics by mucoadhesive tailored biopolymer-based nanoparticles: A review. Food Hydrocoll. 2021, 118, 106772. [Google Scholar] [CrossRef]
- Lamson, N.G.; Berger, A.; Fein, K.C.; Whitehead, K.A. Anionic nanoparticles enable the oral delivery of proteins by enhancing intestinal permeability. Nat. Biomed. Eng. 2020, 4, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Liang, Y.; Liu, L.; Yin, M.; Wang, A.; Sun, K.; Li, Y.; Shi, Y. Research on the fate of polymeric nanoparticles in the process of the intestinal absorption based on model nanoparticles with various characteristics: Size, surface charge and pro-hydrophobics. J. Nanobiotechnol. 2021, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Abasalizadeh, F.; Moghaddam, S.V.; Alizadeh, E.; Akbari, E.; Kashani, E.; Fazljou, S.M.B.; Torbati, M.; Akbarzadeh, A. Alginate-based hydrogels as drug delivery vehicles in cancer treatment and their applications in wound dressing and 3D bioprinting. J. Biol. Eng. 2020, 14, 8. [Google Scholar] [CrossRef]
- Chandrasekar, V.; Coupland, J.N.; Anantheswaran, R.C. Characterization of nisin containing chitosan-alginate microparticles. Food Hydrocoll. 2017, 69, 301–307. [Google Scholar] [CrossRef]
- Pacheco-Ordaz, R.; Wall-Medrano, A.; Goñi, M.G.; Ramos-Clamont-Montfort, G.; Ayala-Zavala, J.F.; González-Aguilar, G. Effect of phenolic compounds on the growth of selected probiotic and pathogenic bacteria. Lett. Appl. Microbiol. 2018, 66, 25–31. [Google Scholar] [CrossRef]
- Lantzouraki, D.Z.; Sinanoglou, V.J.; Zoumpoulakis, P.G.; Glamočlija, J.; Ćirić, A.; Soković, M.; Heropoulos, G.; Proestos, C. Antiradical–antimicrobial activity and phenolic profile of pomegranate (Punica granatum L.) juices from different cultivars: A comparative study. RSC Adv. 2015, 5, 2602–2614. [Google Scholar] [CrossRef]
- Gullon, B.; Pintado, M.E.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Assessment of polyphenolic profile and antibacterial activity of pomegranate peel (Punica granatum) flour obtained from co-product of juice extraction. Food Control 2016, 59, 94–98. [Google Scholar] [CrossRef]
- Kim, S.; Lee, J.Y.; Jeong, Y.; Kang, C.-H. Antioxidant activity and probiotic properties of lactic acid bacteria. Fermentation 2022, 8, 29. [Google Scholar] [CrossRef]
- Amaretti, A.; Di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef]
- Elbayomi, S.M.; Wang, H.; Tamer, T.M.; You, Y. Enhancement of antioxidant and hydrophobic properties of alginate via aromatic derivatization: Preparation, characterization, and evaluation. Polymers 2021, 13, 2575. [Google Scholar] [CrossRef] [PubMed]
- Soltan, Y.A.; Morsy, A.S.; Hashem, N.M.; Elazab, M.A.; Sultan, M.A.; El-Nile, A.; Marey, H.N.; El Lail, G.A.; El-Desoky, N.; Hosny, N.S. Potential of montmorillonite modified by an organosulfur surfactant for reducing aflatoxin B1 toxicity and ruminal methanogenesis in vitro. BMC Vet. Res. 2022, 18, 387. [Google Scholar] [CrossRef]
- Ruiz, F.O.; Gerbaldo, G.; Asurmendi, P.; Pascual, L.M.; Giordano, W.; Barberis, I.L. Antimicrobial activity, inhibition of urogenital pathogens, and synergistic interactions between lactobacillus strains. Curr. Microbiol. 2009, 59, 497–501. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compound | Concentration, µg/100 mL |
|---|---|
| Cinnamic acid | 13.26 |
| Salicylic acid | 5.36 |
| Chlorogenic acid | 5.33 |
| Syringenic | 4.68 |
| Catechol | 3.1 |
| Caffeic | 2.45 |
| Gallic | 2.19 |
| Concentration, µg/mL | DPPH Scavenging Activity, % | ||
|---|---|---|---|
| Nanoencapsulated Synbiotic | Free Synbiotic | Ascorbic Acid | |
| 1.95 | 41.2 a | 1.77 b | 41.7 a |
| 3.90 | 47.7 a | 3.5 b | 45.8 a |
| 7.81 | 56.5 a | 7.1 b | 56.3 a |
| 15.62 | 63.8 a | 14.2 b | 64.2 a |
| 31.25 | 70.6 a | 24.8 b | 71.2 a |
| 62.5 | 77.6 a | 47.8 b | 78.0 a |
| 125 | 85.3 a | 56.2 b | 86.4 a |
| 250 | 90.3 a | 62.2 b | 92.7 a |
| 500 | 94.3 a | 68.7 b | 94.5 a |
| 1000 | 96.2 a | 74.1 b | 97.0 a |
| SEM | 9.56 | 9.78 | 9.73 |
| Half-maximal inhibitory concentration | |||
| IC 50 | 3.96 ± 0.42 b | 65.75 ± 2.14 a | 4.08 ± 0.79 b |
| Treatment | Inhibition Zone, mm | |||||
|---|---|---|---|---|---|---|
| Bacterial Strains | Fungal Strains | |||||
| Escherichia coli (ATCC 8739) | Staphylococcus aureus (ATCC 6538) | Pseudomonas aeruginosa (ATCC 90274) | Salmonella typhi (ATCC 6539) | Aspergillus niger | Aspergillus flavus | |
| Nanoencapsulted synbiotic | 19.00 a | 20.67 a | 20.33 a | 19.33 a | 16.67 b | 17.00 b |
| Free synbiotic | 13.00 c | 16.00 b | 15.33 b | 19.00 a | 14.67 c | 13.33 c |
| Gentamicin | 17.00 b | 18.00 a,b | 19.33 a | 15.67 b | - | - |
| Fluconazol | - | - | - | - | 26.00 a | 21.00 a |
| SEM | 0.374 | 0.470 | 0.795 | 1.13 | 1.03 | 1.73 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hashem, N.M.; Hosny, N.S.; El-Desoky, N.; Soltan, Y.A.; Elolimy, A.A.; Sallam, S.M.A.; Abu-Tor, E.-S.M. Alginate Nanoencapsulated Synbiotic Composite of Pomegranate Peel Phytogenics and Multi-Probiotic Species as a Potential Feed Additive: Physicochemical, Antioxidant, and Antimicrobial Activities. Animals 2023, 13, 2432. https://doi.org/10.3390/ani13152432
Hashem NM, Hosny NS, El-Desoky N, Soltan YA, Elolimy AA, Sallam SMA, Abu-Tor E-SM. Alginate Nanoencapsulated Synbiotic Composite of Pomegranate Peel Phytogenics and Multi-Probiotic Species as a Potential Feed Additive: Physicochemical, Antioxidant, and Antimicrobial Activities. Animals. 2023; 13(15):2432. https://doi.org/10.3390/ani13152432
Chicago/Turabian StyleHashem, Nesrein M., Nourhan S. Hosny, Nagwa El-Desoky, Yosra A. Soltan, Ahmed A. Elolimy, Sobhy M. A. Sallam, and El-Sayed M. Abu-Tor. 2023. "Alginate Nanoencapsulated Synbiotic Composite of Pomegranate Peel Phytogenics and Multi-Probiotic Species as a Potential Feed Additive: Physicochemical, Antioxidant, and Antimicrobial Activities" Animals 13, no. 15: 2432. https://doi.org/10.3390/ani13152432